中國荷斯坦奶牛Nramp1基因多態性與泌乳性狀及產奶量的關聯性分析
2017-04-08 03:00:05劉麗霞李強子劉吳鑫
浙江農業學報 2017年3期
張 麗,劉麗霞,陳 紅,李強子,劉吳鑫
(西北民族大學 生命科學與工程學院,甘肅 蘭州730030)
中國荷斯坦奶牛Nramp1基因多態性與泌乳性狀及產奶量的關聯性分析
張 麗,劉麗霞,陳 紅,李強子,劉吳鑫
(西北民族大學 生命科學與工程學院,甘肅 蘭州730030)
為尋找與乳房炎相關的分子標記,為荷斯坦牛的抗病育種提供理論基礎,試驗采用PCR-SSCP技術和測序的方法對寧夏農墾303頭中國荷斯坦奶牛的Nramp1基因進行了遺傳多態性分析,利用一般線性模型分析Nramp1基因c.200C>G突變對泌乳性狀及產奶量影響。結果顯示,Nramp1基因c.200C>G突變與乳脂率、日產奶量和體細胞評分值存在顯著(P<0.05)或極顯著(P<0.01)關聯。因此,可以把該位點作為中國荷斯坦奶牛體細胞評分、測定日產奶量和日乳脂率的遺傳標記進行進一步研究。
荷斯坦牛;Nramp1基因;遺傳多態性;泌乳性狀;產奶量
奶牛乳腺炎在泌乳期牛群體中發病率極高,其發病率與抗病候選基因的多態性相關。因此,利用分子標記輔助選擇技術選育乳腺炎抗性的奶牛,有望成為控制和降低乳腺炎發生的有效途徑。天然抗性相關巨噬蛋白基因(natural resistance associated macrophage protein,Nramp1)首次在小鼠中被發現并克隆[1],其主要在巨噬細胞和多核型白細胞中特異表達,通過轉運細菌自身合成防御酶系所必需的Mn2+或Fe2+等金屬離子使細菌無法合成防御酶系,從而使動物抵抗病原菌的侵染[2-3]。Nramp1在家畜機體中具有重要的抵抗胞內寄生菌活性,已經成為家畜抗病育種的研究熱點之一。研究表明,Nramp1基因與牛布氏桿菌病、結核病等傳染病的抗性相關。Nramp1基因3’UTR區遺傳變異對荷斯坦奶牛布氏桿菌、分枝桿菌的抗性差異顯著[4-8]。吳宏梅等[9]研究表明,豬Nramp1基因內含子6多態性與中性粒細胞及單核巨噬細胞吞噬功能顯著相關。Hebert等[10]報道,Nramp1基因可抵抗細胞內乳腺炎病原金黃色葡萄球菌的感染。Joo等[11]報道,乳腺炎抗性奶牛Nramp1基因mRNA表達量高于易感牛群,可根據Nramp1基因表達量的差異選育乳腺炎抗性奶牛群,從而揭示Nramp1基因多態性與乳腺炎抗性相關。胡海川[12]研究表明,荷斯坦奶牛Nramp1基因多態性與體細胞評分(somatic cell score,SCS)、305 d產奶量存在顯著或極顯著相關性,且SCS數值越高,產奶量下降越明顯,牛場效應和胎次效應對乳腺炎的影響也很大。因此,可將Nramp1基因作為奶牛乳腺炎候選基因應用于分子標記輔助選擇育種。但是,牛Nramp1基因的研究多局限于3’UTR區與布氏桿菌病、結核桿菌病的相關性研究,對其他位點的單核苷酸多態(single nucleotide polymorphism,SNP)及其與泌乳性狀及產奶量的研究未見報道。本研究采用PCR-SSCP和測序技術對Nramp1基因的SNP進行了篩選,并結合乳脂率、乳蛋白率、體細胞評分及產奶量數據,分析其多態性與中國荷斯坦奶牛泌乳性狀和乳腺炎間的相關性,找到對泌乳性狀和乳腺炎間有顯著影響的SNP位點,為奶牛抗病育種提供參考。
1 材料與方法
1.1 試驗牛血樣采集、產奶量和乳成分測定
本試驗樣本采集于2013年3月—2014年5月在寧夏農墾賀蘭山奶業有限公司完成。選擇體況相近、無臨床乳腺炎、胎次在 3~4 胎具有完整系譜資料的中國荷斯坦奶牛303頭,經尾靜脈采血10 mL·頭-1,醫用采血管低溫保存帶回實驗室冷凍備用。試驗牛每日采集一次奶樣,記錄日產奶量,全天按照早∶中∶晚=4∶3∶3的比例混合奶樣100 mL送寧夏 DHI 測定中心進行個體牛乳蛋白率、乳脂率及乳中體細胞數測定。對泌乳天數多于305 d的個體只取其305 d的產奶量,對泌乳天數少于305 d的個體根據校正系數校正成305 d產奶量。
1.2 血樣DNA提取和引物設計
采用常規苯酚-氯仿抽提法從備用凍存血樣中提取荷斯坦奶牛基因組DNA,超純水溶解,保存母液,取部分DNA母液樣品稀釋至100 ng·μL-1,-20 ℃保存備用。
根據GenBank公布的荷斯坦奶牛Nramp1基因DNA序列(登錄號:DQ493965),應用Primer6.0軟件設計2對特異性引物用于擴增荷斯坦奶牛Nramp1基因第10外顯子和第11外顯子序列,其引物序列見表1,該引物由大連寶生物公司合成,雙蒸水溶解,-20 ℃保存備用。
1.3 PCR-SSCP檢測與等位基因序列測定
PCR擴增體系總體積為20 μL:基因組DNA模板2.0 μL,0.25 μmol·L-1上下游引物各1.0 μL,2×TaqPCR MasterMix 10.0 μL,雙蒸水6.0 μL。PCR擴增程序為:94 ℃預變性4 min;94 ℃變性30 s,60 ℃退火30 s,72 ℃延伸30 s,35個循環;72 ℃延伸5 min,降溫至4 ℃保存,1%瓊脂糖凝膠對其擴增結果進行檢測。
取3 μL無非特異性條帶的PCR產物,加入7 μL變性上樣緩沖液(0.025%溴酚藍、98%去離子甲酰胺、10 mmol·L-1EDTA、0.025%二甲苯氰),98 ℃變性10 min后再立即冰浴10 min。將變性后的PCR產物依次上樣到10%的非變性聚丙烯酰胺凝膠孔中(Acr∶Scr=39∶1),室溫160 V電壓下分別電泳4 h(exon10)和6 h(exon11)。電泳結束后用硝酸銀染色法染色確定PCR產物的基因型。
表1 引物信息
Table 1 The information of primers

引物名稱Primername引物序列Primersequence(5’-3’)退火溫度Annealingtemperature/℃擴增區域Amplifiedregion目的片段長度Productsize/bpP1F:5’-TGTCGGGCTGGATTCAGACT-3’R:5’-CCTGCCGTTGGCTTGCTTAC-3’60exon10231P2F:5’-GACCGTGGCAGTGGACATT-3’R:5’-CTGCCTTGTGCTCAGACACC-3’60exon11327
PCR產物經SSCP電泳判型后,如果新發現的等位基因是純合型個體,其PCR產物直接送北京華大公司測序。如果新發現的等位基因是雜合型個體,可從聚丙烯酰胺凝膠中切下等位基因中的一條帶,放于1.5 mL離心管中,用200 μL 1×TE(10 mmol·L-1Tris-HCl,0.1 mmol·L-1EDTA)緩沖液洗滌兩次,并用槍頭壓碎后加入50 μL 1×TE,置于50~60 ℃水浴中孵化1~2 h左右,該溶液可作為DNA模板并使用原有的PCR擴增體系和條件進行再擴增,第二次PCR擴增產物用瓊脂糖凝膠檢測后,與原PCR產物或其他含有該等位基因的樣品同時進行SSCP電泳,以確保第二次擴增產物為新發現的等位基因。然后用PCR純化試劑盒對第二次擴增產物純化并測序[13]。
1.4 數據分析
利用GenePop和PIC生物軟件計算突變位點的遺傳參數。因體細胞數(somatic cell count,SCC)呈非正態分布,可將體細胞數轉換為體細胞評分,公式為:SCS=[log2(SCC/100)]+3[14]。
荷斯坦奶牛群均為舍飼群體,飼養環境基本一致,可忽略場效應對性狀的影響。因此,應用SPSS 23.0 統計軟件中一般線性模型分析荷斯坦奶牛Nramp1基因突變對測定日乳脂率、測定日乳蛋白率、日產奶量、305 d 產奶量和體細胞評分進行差異顯著性檢驗。不同基因型間的多重比較用LSD 法。模型如下:Yijkl=μ+Ci+Tj+Gk+Sl+eijkl,其中Yijkl為個體性狀表型值;μ為群體均值;Ci為環境的固定效應;Tj為第j胎次的固定效應;Gk為Nramp1基因第k個標記基因型的固定效應;Sl為產犢季節的固定效應;eijkl為隨機誤差效應。
2 結果與分析
2.1Nramp1基因PCR擴增產物質量
荷斯坦奶牛Nramp1基因第10、11外顯子PCR擴增產物經1%瓊脂糖凝膠(160 V,30 min)電泳檢測條帶清晰且特異性較好(圖1),大小范圍分別為200~300 bp、300~400 bp,與預期片段長度基本相符,可以用于SSCP檢測。
2.2 PCR-SSCP檢測分析和序列測定
荷斯坦奶牛Nramp1基因第10、11外顯子PCR擴增產物經SSCP分析,結果表明,荷斯坦奶牛Nramp1基因第10外顯子無多態性;第11外顯子區域檢測到2種等位基因A和B,形成AA、BB、AB共3種帶型(圖2),其中等位基因A和B均發現純合型個體。第11外顯子PCR擴增產物測序結果通過與普通牛Nramp1基因(GenBank No. DQ493965)第11外顯子序列比對,發現擴增序列的200 bp處發生了G到C的錯義突變(圖3),從而導致氨基酸由原來的半胱氨酸(Cys)變為絲氨酸(Ser)(圖4)。

M, 100 bp DNA Marker; 泳道1為Exon10 PCR產物;泳道2為Exon11 PCR產物M, 100 bp DNA marker; Lane 1, PCR product of Exon10; Lane 2, PCR product of Exon11圖1 荷斯坦奶牛Nramp1基因第10和11外顯子PCR擴增產物Fig.1 Detections of PCR products of Nramp1 gene exon 11 in Holstein cattle

圖2 荷斯坦奶牛Nramp1基因第10/11外顯子PCR產物SSCP檢測結果Fig.2 SSCP analvsis of PCR amplification of Nramp1 exon10/11 in Holstein cattle
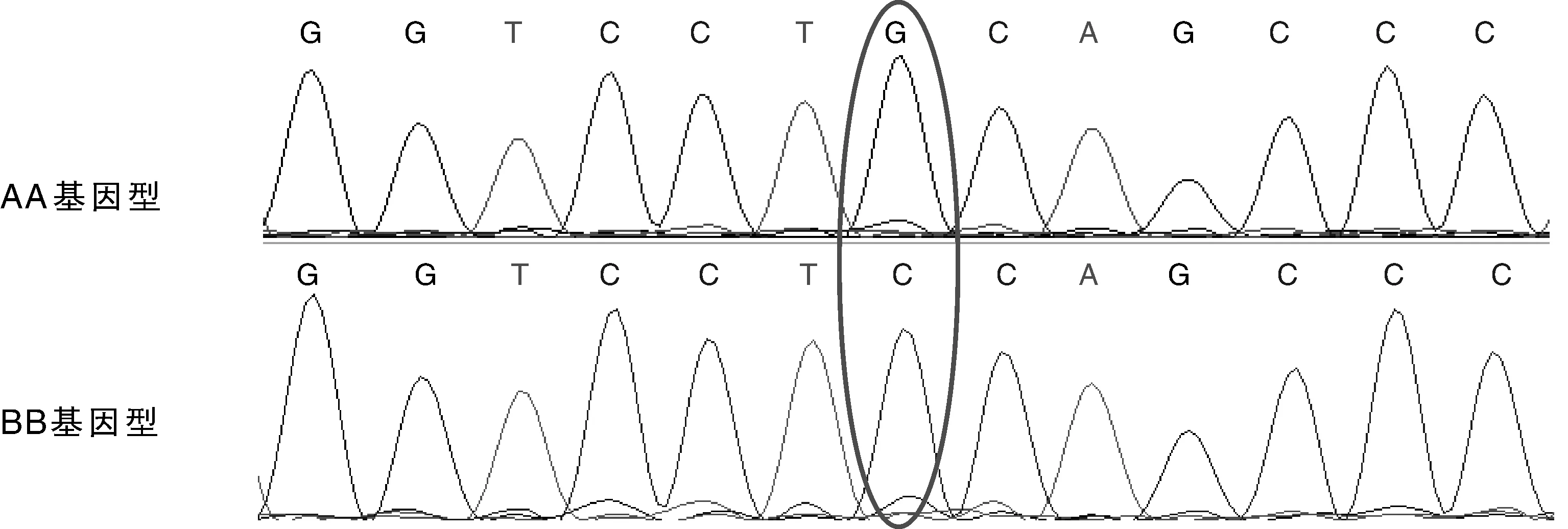
圖3 荷斯坦奶牛Nramp1基因第11外顯子區域AA/BB基因型堿基突變Fig.3 Base mutations of AA/BB genotype of Nramp1 exon11 area in Holstein cattle

圖4 荷斯坦奶牛Nramp1基因第11外顯子區域AA/BB基因型編碼蛋白序列Fig.4 Encoding protein sequence of AA/BB genotype of Nramp1 exon11 area in Holstein cattle
2.3 荷斯坦奶牛Nramp1基因群體遺傳特性分析
對荷斯坦奶牛Nramp1基因第11外顯子的各遺傳參數指標進行了分析,其中等位基因A的頻率為0.566 0,為優勢等位基因;基因型AA的頻率為0.445 6,為優勢基因型。等位基因B的頻率為0.434 0,基因型BB和AB的頻率分別為0.313 5和0.240 9。純合度(Ho)、雜合度(He)、有效等位基因數(Ne)分別為0.507 9、0.492 1和1.965 7;多態信息含量(PIC)為0.370 6,即0.25
2.4 荷斯坦奶牛Nramp1基因第11外顯子突變與泌乳性狀和產奶量的關聯分析
303頭荷斯坦奶牛群體的Nramp1基因第11外顯子AA、BB、AB基因型對日產奶量、測定日乳脂率、測定日乳蛋白率、SCS值及305 d產奶量5個生產性狀的影響見表2。方差分析結果表明,荷斯坦奶牛的乳脂率、日產奶量、體細胞評分在基因型間有顯著或極顯著差異(P<0.05,P<0.01)。多重比較結果表明,BB基因型乳脂率、日產奶量分別為(3.83±0.08)%、(35.38±1.42)kg,顯著(P<0. 05)和極顯著(P<0.01)高于AA基因型個體相應性狀(3.57±0.07)%、(28.73±1.16)kg,BB基因型體細胞評分(2.35±0.10)顯著低于AA基因型(P<0.05)。測定日乳蛋白率和305 d產奶量在不同基因型間無顯著差異,但AB型個體介于AA型和BB型兩者之間。
表2 荷斯坦奶牛Nramp1基因第11外顯子基因型對泌乳性狀及體細胞評分的關聯分析
Table 2 Influence ofNramp1 gene exon11 genotype on milk traits and SCS of Holstein cattle

基因型Genotype數量Number乳脂率Fatpercentage/%乳蛋白率Proteinpercentage/%日產奶量Dailymilkyield/kg305d產奶量305dmilkyield/kg體細胞評分SCSAA1353.57±0.07b3.17±0.0528.73±1.16B8412.66±393.052.90±0.20aAB953.83±0.09a3.27±0.0632.58±1.79AB8304.04±646.282.86±0.18abBB733.83±0.08a3.22±0.0535.38±1.42A7851.42±512.922.35±0.10b
同列數據后無相同大寫字母表示差異極顯著(P<0.01),無相同小寫字母表示差異顯著(P<0.05)。
Values in each column without the same capital or lowercase letters are significantly different at the probability level of 0.01 or 0.05, respectively.
3 討論
3.1 荷斯坦奶牛Nramp1基因多態性分析
大多數高等哺乳動物Nramp1基因外顯子和內含子間變異很小,但是其3’和5’非編碼區存在很大變異,因此有關荷斯坦奶牛Nramp1基因多態性的研究主要集中3’UTR區[4-8]。本研究僅在荷斯坦奶牛Nramp1基因第11外顯子中發現擴增片段的第200位發生的C→G堿基突變,該突變導致蛋白質氨基酸序列由半胱氨酸(Cys)轉變為絲氨酸(Ser),而在第10外顯子中未發現多態位點。同胡海川[12]研究對比發現,擴增片段相同卻出現了不同的結果。胡海川[12]研究發現,荷斯坦奶牛Nramp1基因第10外顯子存在2個等位基因3種基因型,第11外顯子第200位、254位分別存在C→G、T→G的堿基突變,造成這種差異可能是由于所研究的荷斯坦奶牛群體的遺傳背景和基因與環境相互作用不同而造成的差異,再者我們也應擴大樣本容量,避免偶然情況發生;另外第10外顯子突變位點的PIC低于0.25,說明該位點低度多態,遺傳變異較小,可望獲得的遺傳進展不大。
本研究中荷斯坦奶牛Nramp1基因第11外顯子多態信息含量(PIC)介于0.25和0.50之間,說明該群體多態信息含量已達到中度多態,且該群體未偏離Hardy-Weinberg平衡(P>0.05),因此荷斯坦奶牛群體在此位點的多態性相對較高,遺傳變異相對較大,可望獲得更多的遺傳進展,可以很好地利用這一位點在本試驗群體中表現出良好的多態性選擇。馬捷瓊等[15]研究表明,奶牛Nramp1基因3′UTR區具有豐富的多態性,多態信息含量為0.950,處于高度多態,該群體未偏離Hardy-Weinberg平衡,這可能與人工選擇和飼養環境有關。
3.2 荷斯坦奶牛Nramp1基因多態性與泌乳性狀及產奶量的關聯性
Nramp1基因是動物體內殘余固有免疫反應的重要調節因子。候選基因作為尋找篩選畜禽重要經濟性狀的遺傳連鎖標記有效方法,可以直接研究該基因的多態性與個體經濟性狀的關系。由于牛乳中SCC能準確反映奶牛乳腺炎的發病程度,同時SCS與乳腺炎的遺傳相關為0.6~0.8,并且SCS的增加可降低產奶量,因此本試驗以SCS為乳腺炎的參數,研究各基因型與SCS、產奶量之間的相關性。馬捷瓊等[16]研究表明,Nramp1基因3′UTR區多態性與奶牛305 d產奶量和乳房炎檢出率均達到顯著相關(P<0.05)。郭洋等[17]研究發現,乳中SCC越低,305 d產奶量較高。
本研究發現,Nramp1基因第11外顯子G→C的突變位點對荷斯坦奶牛體細胞評分的影響顯著(P<0.05),對測定日產奶量影響極顯著(P<0.01),對測定日乳脂率影響顯著(P<0.05),但對305d產奶量影響不顯著(P>0.05),因此可以把該位點作為中國荷斯坦奶牛體細胞評分、測定日產奶量和測定日乳脂率遺傳標記進行研究。中國荷斯坦奶牛Nramp1基因第11外顯子的突變與泌乳性狀的關聯分析結果表明,AA基因型個體305 d產奶量最高,為8 412.66 kg,但其乳脂率(3.57±0.07)%、乳蛋白率(3.17±0.05)%和日產奶量(28.73±1.16)kg卻是各種基因型中最低的。中國荷斯坦奶牛不同基因型個體泌乳性狀的分析結果和突變檢測情況提示,Nramp1基因第11外顯子的G→C突變對奶牛的泌乳性狀可能存在重要影響。
[1] VIDAL S, GROS P, SKAMENE E. Natural resistance to infection with intracellular parasites: molecular genetics identifies Nramp1 as the Bcg/Ity/Lsh locus[J].JournalofLeukocyteBiology,1995,58(4):382-390.
[2] SUPER F, SUPEKOVA L,NELSON H, et al. A yeast manganese transporter related to the macrophage protein involved in con-ferring resistance to mycobacteria[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,1996,93(10):5105-5110.
[3] FLEMING M, TRENOR C, SU M A, et al. Microcytic anaemia mice have a mutation in Nramp2, a candidate iron transporter gene[J].NatureGenetics,1997,16(4):383-386.
[5] BARTHEL R, FENG J, PIEDRAHITA J A, et al. Stable transfection of the bovine NARMP1 gene into murine RAW264.7 cells: Effect onBrucellaabortussurvival[J].InfectionandImmunity, 2001, 69(5): 3110-3119.
[6] KUMAR N, MITRA A, GANGULY I, et al. Lack of association of brucellosis resistance with(GT)(13)microsatellite allele at 3’UTR of NRAMP1 gene in Indian zebu (Bosindicus) and crossbred (Bosindicus×Bostaurus) cattle[J].VeterinaryMicrobiology,2005,111(l):139-143.
[7] PAIXAO T A, FERREIRA C, BORGES A M, et al. Frequency of bovine Nramp1 alleles in Holstein and Zebu breeds[J].VeterinaryImmunologyandImmunopathology, 2006, 109(1): 37-42.
[9] 吳宏梅, 王立賢, 程篤學, 等. 豬Nramp1基因多態性與免疫功能的相關性[J].中國農業科學, 2008, 41(1): 215-220. WU H M,WANG L X,CHENG D X, et al. Relationship between polymorphisms ofNramp1 gene and immune function of pig[J].ScientiaAgriculturaSinica, 2008,41(1):215-220.(in Chinese with English abstract)
[10] HéBERT A, SAYASITH K, SéNéCHAL S, et al. Demonstration of intracellularStaphylococcusaureusin bovine mastitis alveolar cells and macrophages isolated from naturally infected cow milk[J].FEMSMicrobiologyLetters, 2000, 193(1): 57-62.
[11] JOO Y S, MOON J S, FOX L K, et al. Comparison of natural resistance-associated macrophage protein (NRAMP) 1 expression between cows with high and low milk somatic cells counts[J].AsianAustralasianJournalofAnimalSciences, 2003, 16(12): 1830-1836.
[12] 胡海川.中國荷斯坦牛Nramp1基因部分序列分析及其與乳腺炎關聯研究[D].雅安:四川農業大學, 2008. HU H C. Sequence analysis of partial NramP1 gene and the association with mastitis in Chinese Holstein cattle[D].Ya’an: Sichuan Agricultural University, 2008. (in Chinese with English abstract)
[13] HU J, ZHOU H, SMYTH A, et al. Polymorphism of the bovine ADRB3 gene[J].MolecularBiologyReports, 2010, 37(7): 3389-3392.
[14] SHOOK G E. Genetic improvement of mastitis through selection on somatic cell count[J].TheVeterinaryClinicsofNorthAmericanFoodAnimalPractice,1993,9(3):563-581.
[15] 馬捷瓊, 陳宏, 陳宗芳, 等. 徐州荷斯坦牛Nramp1基因多態性與乳腺炎通徑分析[J].黑龍江畜牧獸醫, 2007,42(4):1-2. MA J Q, CHEN H,CHEN Z F, et al. Path Analysis of the polymorism ofNramp1 gene and mastitis in Xuzhou Holstein cattle[J].HeilongjiangAnimalScienceandVeterinaryMedicine, 2007,42(4):1-2. (in Chinese)
[16] 馬捷瓊,劉纏民,陳宏,等. 徐州荷斯坦牛Nramp1基因多態性與產奶量通徑分析[J].江蘇農業科學, 2007(1):116-118. MA J Q, LIU C M, CHEN H, et al. Path analysis ofNramp1 gene polymorphism and milk productivity of Holstein cattle in Xuzhou, Jiangsu Province[J].JiangsuAgriculturalSciences, 2007(1):116-118. (in Chinese with English abstract)
[17] 郭洋, 王洪梅, 侯明海, 等. 中國荷斯坦牛SLC11A1基因多態性與乳腺炎的相關性研究[J]. 中國農業科學, 2010, 44(19): 4072-4080. GUO Y, WANG H M, HOU M H, et al. Polymorphisms ofSLC11A1 gene and their association with mastitis in Chinese Holstein[J].ScientiaAgriculturaSinica,2010, 44(19): 4072-4080. (in Chinese with English abstract)
(責任編輯 張 韻)
Genetic polymorphismNramp1 gene and its associations with milk traits and milk yield in Chinese Holstein cattle
ZHANG Li, LIU Lixia, CHEN Hong, LI Qiangzi, LIU Wuxin
(CollegeofLifeScienceandEngineering,NorthwestMinzuUniversity,Lanzhou730030,China)
To search the molecular markers that are associated with mastitis and to provide a theoretical basis for the Holstein cattle breeding and disease resistance, the polymoprhisms ofNramp1 gene in 303 Holstein cattle in Ningxia were detected by the PCR-SSCP technique and the effect ofNramp1 gene c.200C>G locus mutation on milk traits and milk yield was analyzed using a general linear model. The results showed that there was a significant relationship betweenNramp1 gene c.200C>G locus mutation and milk fat, milk yield and somatic cell score at the probability level of 0.05 or 0.01. Therefore we can put the site as cattle somatic cell score, daily milk yield measurement and daily milk fat genetic markers in Chinese Holstein.
Holstein cattle;Nramp1 gene; genetic polymorphism; milk traits; milk yield
10.3969/j.issn.1004-1524.2017.03.06
2016-09-16
西北民族大學引進人才科研項目(xbmuyjrc201316);西北民族大學基本科研業務費專項資金資助項目(31920140077)
張麗(1980—),女,寧夏石嘴山人,博士,副教授,主要從事奶牛分子育種研究。E-mail: zhangli2008@aliyun.com
S823.2
A
1004-1524(2017)03-0389-06
浙江農業學報ActaAgriculturaeZhejiangensis, 2017,29(3): 389-394
http://www.zjnyxb.cn
張麗,劉麗霞,陳紅,等. 中國荷斯坦奶牛Nramp1基因多態性與泌乳性狀及產奶量的關聯性分析[J].浙江農業學報,2017,29(3): 389-394.
