TET酶及其中間產物的研究進展*
2017-04-10 09:29:05程易塵吳曉明
中國病理生理雜志 2017年3期
關鍵詞:檢測
程易塵, 吳曉明, 羅 瑛
(昆明理工大學醫學院,云南 昆明 650500)
TET酶及其中間產物的研究進展*
程易塵, 吳曉明, 羅 瑛△
(昆明理工大學醫學院,云南 昆明 650500)
DNA甲基化的形式主要包括以下3種:5-甲基胞嘧啶(5mC)、N6-甲基嘌呤(N6mA)以及7-甲基鳥嘌呤(7mG)[1]。目前,研究得最為廣泛也最為透徹的就是主要存在于CpG島中胞嘧啶的甲基化修飾,即5mC。在哺乳動物基因組中大約有70%~80%的CpG島區域的胞嘧啶存在甲基化修飾[2]。而CpG島的甲基化是基因沉默的一個重要標志,在調控基因表達過程中發揮著至關重要的作用。
DNA的甲基化修飾是由DNA甲基化酶(DNA methyltransferases,DNMTs)催化形成。DNMTs將由S-腺苷甲硫氨酸提供的活性甲基基團轉移參入到堿基中從而形成甲基化修飾。DNA甲基化酶主要成員包括DNMT1、DNMT3A、DNMT3B以及DNMT3L。其中DNMT1是維持型的甲基化轉移酶:它的作用是在染色體的復制過程中維持子代染色體原有的甲基化修飾。DNMT3A和DNMT3B是從頭合成型的DNA甲基化轉移酶,可以在未發生甲基化修飾的堿基中摻入甲基基團,在胚胎發育過程中發揮著重要的作用[3-4]。DNMT3L不具有甲基化轉移酶活性,可以輔助DNMT3A和DNMT3B的催化活性[2]。然而,現階段DNA的去甲基研究得還不夠成熟。去甲基化酶TET(ten-eleven translocation)家族的發現為DNA去甲基化機制提供了新的視角。而其氧化過程的中間產物5-羥甲基胞嘧啶(5-hydroxymethylcytosine, 5hmC)、5-醛基胞嘧啶(5-formylcytosine, 5fC)和5-羧基胞嘧啶(5-carboxylcytosine, 5caC)很可能不僅僅是反應的中間體,還在生命過程中發揮著重要的作用[5-7]。
1 TET酶家族
1.1 TET酶介導的DNA去甲基化機制 DNA的去甲基主要有2種機制,一種是被動的去甲基化,在DNA復制過程中由于維護甲基化修飾機制的缺失(如維持性甲基化酶DNMT1的缺失或功能喪失)而失去這一修飾;另一種是主動的去甲基化,在去甲基化酶的作用下去除DNA的甲基化修飾。其中被動型的去甲基化機制已經被人們廣泛接受,而主動型的去甲基化的發生機制仍有爭議[8]。
TET酶家族的發現為主動型的去甲基化機制提供的全新的視角和方向。TET甲基胞嘧啶雙加氧酶是分子量為180~230 kD的多亞基蛋白,包含TET1、TET2和TET3 3個成員。所有的TET蛋白在C末端都有一個由雙鏈β螺旋結構域、富含半胱氨酸區域、輔酶因子Fe和2-氧化戊二酸結合區組成的催化中心。研究表明這個催化中心優先與CpG島的胞嘧啶結合但與周圍的堿基沒有相互作用,并且沒有或者幾乎沒有顯示出序列特異性[9-11]。除了催化中心,TET1和TET3在N末端還包含有一個可與DNA結合的CXXC鋅指結構域[12]。它們的作用是氧化5mC形成5hmC,進一步的氧化作用形成5fC和5caC。5fC和5caC最終通過由胸腺嘧啶DNA糖基化酶(thymine-DNA glycosylase,TDG)介導的堿基切除修復(base excision repair,BRE)機制轉化為未修飾的C,從而完成去甲基的過程[6, 13]。
1.2 TET酶與腫瘤的關系及最新的研究進展 在眾多的腫瘤中,TET酶的異變與惡性造血細胞瘤的關系最為密切,并且得到許多實驗數據的支持。最近數據顯示TET1與混合系白血病(mixed lineage leukemia,MLL)是密切相關的。TET1可以直接靶向MLL融合蛋白,并且在MLL重排白血病中它的表達量顯著上調,相應引起5hmC升高[14]。在各種惡性血癌中常會發現TET2的突變,包括:急性髓性白血病(acute myeloid leukemia,AML)、骨髓異常增生綜合征(myelodysplastic syndrome,MDS)、慢性粒單核細胞白血病(chronic myelomonocytic leukemia,CMML)和骨髓增殖性腫瘤(myeloproliferative neoplasms, MPN)[15]。并且研究發現單倍劑量不足是TET突變后引起腫瘤發生的一個最主要的機制[16]。至于TET3,目前還沒有發現腫瘤與體細胞中TET3的突變直接相關[17]。而在實體瘤中,雖然也發現了TET酶的突變,但是其突變率卻非常低。然而,研究表明在各種實體瘤中如胃癌、前列腺癌、肝癌、肺癌和乳腺癌以及膠質母細胞瘤和黑色素瘤中經常會觀察到TET酶的表達下調,而相應的5hmC含量也會降低[18-20]。而這一調控機制目前還不甚明了,仍然有待進一步的探索。研究表明microRNAs可能在其中發揮著一定的作用[21-22]。例如,miR-22可以直接與3種TET酶的mRNA結合并促進其降解過程,從而下調其表達[23]。
近期研究發現,TET酶可以通過調節亞端粒甲基化水平在胚胎干細胞的端粒維持和染色體穩定性中發揮重要作用[24]。此外,TET1可以誘導上皮間質細胞的轉化,并且作為一種輔助激活因子調控基因的轉錄,在胚胎發育過程中發揮重要的調節作用[25]。在MLO-Y4細胞中敲低TET3的表達可以解除地塞米松誘導的Akt通路的抑制,從而為激素性骨壞死(steroid-associated osteonecrosis,SAON)提供了一個新的治療靶點[26]。另外,除了作為一種酶具有的催化作用,TET酶家族還可以通過與一些表觀修飾的相互作用,在腫瘤和神經系統的發育過程發揮重要作用[27]。
2 氧化中間產物: 5hmC、5fC和5caC
近期研究表明基因組DNA中胞嘧啶的羥甲基化和低水平的乙酰化都是相對穩定的修飾形式[28]。這也就意味著大多數我們觀察到的5hmC和5fC并不是一個催化反應過程中短暫的瞬態中間體,而是會在一段時期內穩定的存在。并且,酶反應動力學分析顯示TET酶對5mC的氧化速度是對5hmC的4.9~6.3倍,當底物是5fC時這個數值升高至7.8~12.6倍[10]。早期的質譜數據分析表明在所有現經檢測的組織中都檢測到了5mC氧化產物的存在,而它們的含量具有一定的組織差異性。5mC存在于所有的組織中,基因組中大概有4%~5%的胞嘧啶通過甲基化作用修飾形成5mC。在它的氧化產物中含量最為豐富的是5hmC,在中樞神經系統中5hmC存在的比例為0.3%~0.7%,而在脾臟和睪丸中這一數值降為0.03%~0.06%,顯示出較強的組織差異性[29-30]。5mC的其它兩個氧化產物5fC和5caC的含量相對較少,大概為5hmC的1/10~1/100,或者低至難以檢測[28, 31]。
2.1 5hmC、5fC和5caC的檢測方法 目前很多研究人員都致力于繪制出基因組中5mC氧化產物的圖譜,以期揭示它們在DNA甲基化動態調控以及基因調控中的作用。下面簡要介紹一下在單堿基分辨率水平檢測5mC氧化產物的方法。基因組中5mC氧化產物豐度太低,給檢測造成了一定的技術障礙。早期主要是通過特異性的抗體和化學標記對相應的氧化產物進行親和富集,但是這種方法的分辨率很低,無法進行精確定位。為了解決這一限制,近期研究人員開發出新的實驗方法,在基因組水平對單個氧化5mC進行特異性檢測。目前各種研究方法主要是基于亞硫酸鹽測序法。首先常規的亞硫酸鹽測序法并不能區分5mC和其各種氧化形式。通過亞硫酸鹽處理和PCR擴增后,5mC和5hmC還是為胞嘧啶C,而5fC和5caC則會轉化為T,因此常規的亞硫酸鹽測序法檢測出來的是5mC和5hmC的總和。因而,我們需要在此基礎上對堿基進行修飾或者保護才能進一步區分出C的各種修飾形式。
對于5hmC而言目前主要有3種方法:氧化亞硫酸鹽測序法(oxidative bisulfite sequencing,oxBS-seq)、TET輔助亞硫酸鹽測序法(TET-assisted bisulfite sequencing,TAB-seq)和PvuRts1I內切酶聯合化學富集標記測序法(Pvu-Seal-seq)。第1種方法oxBS-seq是利用KRuO4可特異性地將5hmC氧化成5fC,結合亞硫酸鹽處理通過2次測序后進行比對從而達到區分5mC和5hmC的目的,因而需要深度測序。這種方法可以對5hmC位點進行精確定位但是工作量較大[32]。第2種方法TAB-seq首先用β-糖基化轉移酶(β-glucosyltransferase,β-GT)對5hmC進行糖基化修飾保護,再用TET酶進行處理將5mC和5fC轉化成5caC,就可以通過亞硫酸鹽測序將5hmC檢測出來[33-34]。這種方法不需要進行2次測序,但是利用酶處理條件較為苛刻,容易出現催化不完全的現象。第3種方法Pvu-Seal-seq不依賴亞硫酸鹽的處理,首先通過可以特異性識別5hmC位點的PvuRts1I內切酶對基因組DNA進行切割,切割后的片段再進行特異性富集并加上biotin標記,最后測序。Pvu-Seal-seq最大的優勢在于,相對于亞硫酸鹽測序,它非常靈敏,可以檢測到通常亞硫酸鹽處理會丟失的5hmCH(H=A、T、C),而前者通常只能檢測到CpG序列處的5hmC;缺點在于由于前期處理中有富集的步驟,因而無法進行絕對定量[35]。這種方法增加一步NaBH4處理后同樣適用于5fC,NaBH4可將5fC還原為5hmC,從而進行Pvu-Seal-seq檢測。
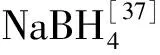
總的來說以上的幾種檢測方式都具有各自的優缺點,目前也沒有一個檢測5mC氧化產物的黃金標準。我們仍然需要摸索新的方法以及進行技術的改進。值得一提的是目前嶄露頭角的三代測序技術,無需前期處理可進行直接測序讀出各種修飾形式,但是這種方法的精密度和通量仍有待提升[38-39]。這也是我們可以期待的一個方向。
2.2 5hmC、5fC和5caC在基因組中的分布 盡管這些技術還存在一些缺陷,由于對各種修飾后的胞嘧啶轉化不完全或者對這種修飾保護機制的欠缺,結果可能會出現一些偏差,但是仍然提供了一個量化的比較直觀的視角。在胚胎干細胞中繪制的5hmC全基因組圖譜顯示:胚胎干細胞中大約有200萬個5hmC修飾位點,其中大概有20%以5hmC的形式存在,并且這一修飾經常發生在CpG含量較低的區域,通常處于低表達基因的啟動子區域、基因中、間隔區以及遠端調控區,如DNase I 超敏位點、增強子和絕緣子區域[33]。相較而言,另外2個氧化產物5fC和5caC在基因組中的修飾位點較少,大概只有33萬,其中8%~10%發生相應修飾[40]。5fC和5caC主要存在于遠端調控元件中,如DNase I 超敏位點、增強子和絕緣子區域,但是在活躍基因的起始轉錄位點也檢測到了它們的存在[40-41]。5fC/5caC在開放染色質和轉錄因子結合區的存在提示我們,染色質重塑和轉錄因子的結合可能在介導TET參與的5mC的氧化去甲基化過程中發揮一定的作用。值得一提的是,絕大多數5hmC修飾位點和5fC/5caC修飾位點是不重疊的,在對大約2 200萬個CpG位點的檢測中只有7.5%的5hmC修飾位點與18.8%的5fC/5caC修飾位點是重合的[42]。這也就意味著大多數5hmC不會轉化為更高的氧化形式而是作為一種獨立的存在,繼而可能行使某些功能。當然我們不能排除一種情況就是這些檢測位點進行了快速的氧化去甲基化過程,以及現階段實驗技術的缺陷,導致我們沒有檢測到。
2.3 5hmC、5fC和5caC功能的研究進展 5mC作為一種穩定的可遺傳的表觀遺傳學標記,它的衍生物除了作為去甲基化過程的中間產物外,可能還具有其它的功能。例如,在腦組織中高峰度的5hmC可能是一個潛在的表觀遺傳標記。近期研究表明DNMT1結合蛋白UHRF1和UHRF2可以綁定5-hmC[13,43]。因此,這2種蛋白可能通過將DNMT1募集到5-hmC上進而維持DNA的甲基化狀態。然而,通過系統地對可與包含5mC、5hmC、5fC或5caC的DNA序列相互作用的蛋白檢測顯示,能與5hmC特異性結合的蛋白很少,而能與5fC/5caC相互作用的蛋白很多,包括TDG、p53、DNA修復因子、染色質重塑因子和叉頭框轉錄因子[13,44]。這其中部分蛋白與DNA的修復和主動去甲基化過程相關,但是這種相互作用的生理生化意義還不是非常清楚。此外,雖然胞嘧啶的修飾形式具有一定的序列特異性,但是大多數DNA結合蛋白無法識別基因組中含量較豐富的5hmC的現象提示我們:5mC修飾狀態的改變可能會在一定程度上抑制蛋白與染色體的結合。例如,MBD1、MBD2和MBD4這些可以與5mC結合的蛋白無法識別5hmC,或者與5hmC的結合能力非常弱[45]。此外,在1型腎母細胞瘤WT1中胞嘧啶的氧化修飾形式(5hmC和5fC)可以顯著干擾轉錄因子與DNA的結合[11],而它們是否對其它DNA結合蛋白有干擾作用還有待進一步的研究。
大量的體外研究數據表明5fC/5caC很可能不僅僅是一種去甲基化過程的中間產物,并且在細胞生命活動中承擔著其它重要的功能。5fC/5caC可以通過對RNA聚合酶Ⅱ(RNA polymerase Ⅱ,Pol Ⅱ)的作用干擾基因轉錄。實驗表明在轉錄過程中,Pol Ⅱ可以直接識別5fC/5caC并且引起瞬時的轉錄減慢[46-47]。鑒于5fC/5caC在基因組和不同細胞之間分布的差異性,這一短暫的轉錄停頓可能參與了基因表達的調控,進一步對細胞生命活動的進程產生影響。例如,在調控大腦中一些長基因的優先表達以及維持神經元的完整性中,這一瞬時延遲的積累可能發揮著重要的作用。Pol Ⅱ在5fC/5caC位點處的轉錄阻滯還有可能為轉錄延伸因子、染色體重塑復合物、TDG和堿基切除修復復合物在氧化胞嘧啶上的定位提供信號[47]。
3 小結與展望
TET酶家族氧化5mC,逐步形成5hmC、5fC和5caC,進而通過TDG和堿基切除修復作用催化形成為修飾的胞嘧啶從而完成基因的主動去甲基化。而在此過程中形成的氧化中間產物5hmC、5fC和5caC可能作為一種潛在的遺傳標記在眾多生命活動中發揮著重要的作用。現階段關于5mC氧化中間產物以及TET作用和調控機制的研究還有一定的局限性,也存在著一些問題,但是隨著相關技術的發展和研究的深入我們將會慢慢揭示它們的存在和意義。
[1] Ramsahoye BH, Biniszkiewicz D, Lyko F, et al. Non-CpG methylation is prevalent in embryonic stem cells and may be mediated by DNA methyltransferase 3a[J]. Proc Natl Acad Sci U S A, 2000, 97(10):5237-5242.
[2] Goll MG, Bestor TH. Eukaryotic cytosine methyltransferases[J]. Annu Rev Biochem, 2005, 74:481-514.
[3] Bestor TH. The DNA methyltransferases of mammals[J]. Hum Mol Genet, 2000, 9(16):2395-2402.
[4] Okano M, Bell DW, Haber DA, et al. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development[J]. Cell, 1999, 99(3):247-257.
[5] Ito S, D’Alessio AC, Taranova OV, et al. Role of Tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification[J]. Nature, 2010, 466(7310):1129-1133.
[6] He YF, Li BZ, Li Z, et al. Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA[J]. Science, 2011, 333(6047):1303-1307.
[7] Rasmussen KD, Helin K. Role of TET enzymes in DNA methylation, development, and cancer[J]. Genes Dev, 2016, 30(7):733-750.
[8] Wu SC, Zhang Y. Active DNA demethylation: many roads lead to Rome[J]. Nat Rev Mol Cell Biol, 2010, 11(9):607-620.
[9] Hu L, Li Z, Cheng J, et al. Crystal structure of TET2-DNA complex: insight into TET-mediated 5mC oxidation[J]. Cell, 2013, 155(7):1545-1555.
[10]Hu L, Lu J, Cheng J, et al. Structural insight into substrate preference for TET-mediated oxidation[J]. Nature, 2015, 527(7576):118-122.
[11]Hashimoto H, Olanrewaju YO, Zheng Y, et al. Wilms tumor protein recognizes 5-carboxylcytosine within a specific DNA sequence[J]. Genes Dev, 2014, 28(20):2304-2313.
[12]Xu Y, Xu C, Kato A, et al. Tet3 CXXC domain and dioxygenase activity cooperatively regulate key genes for Xenopus eye and neural development[J]. Cell, 2012, 151(6):1200-1213.
[13]Spruijt CG, Gnerlich F, Smits AH, et al. Dynamic readers for 5-(hydroxy)methylcytosine and its oxidized derivatives[J]. Cell, 2013, 152(5): 1146-1159.
[14]Huang H, Jiang X, Li Z, et al. TET1 plays an essential oncogenic role in MLL-rearranged leukemia[J]. Proc Natl Acad Sci U S A, 2013, 110(29): 11994-11999.
[15]Shih AH, Abdel-Wahab O, Patel JP, et al. The role of mutations in epigenetic regulators in myeloid malignancies[J]. Nat Rev Cancer, 2012, 12(9): 599-612.
[16]Mariani CJ, Madzo J, Moen EL, et al. Alterations of 5-hydroxymethylcytosine in human cancers[J]. Cancers, 2013, 5(3):786-814.
[17]Zhao H, Chen T. Tet family of 5-methylcytosine dioxyge-nases in mammalian development[J]. J Hum Genet, 2013, 58(7):421-427.
[18]Yang H, Liu Y, Bai F, et al. Tumor development is associated with decrease of TET gene expression and 5-methylcytosine hydroxylation[J]. Oncogene, 2013, 32(5):663-669.
[19]Liu C, Liu L, Chen X, et al. Decrease of 5-hydroxymethylcytosine is associated with progression of hepatocellular carcinoma through downregulation of TET1[J]. PLoS One, 2013, 8(5):e62828.
[20]Lian CG, Xu Y, Ceol C, et al. Loss of 5-hydroxymethylcytosine is an epigenetic hallmark of melanoma[J]. Cell, 2012, 150(6):1135-1146.
[21]Chuang KH, Whitney-Miller CL, Chu CY, et al. Micro-RNA-494 is a master epigenetic regulator of multiple invasion-suppressor microRNAs by targeting ten eleven translocation 1 in invasive human hepatocellular carcinoma tumors[J]. Hepatology, 2015, 62(2):466-480.
[22]Cheng J, Guo S, Chen S, et al. An extensive network of TET2-targeting microRNAs regulates malignant hematopoiesis[J]. Cell Rep, 2013, 5(2): 471-481.
[23]Song SJ, Poliseno L, Song MS, et al. MicroRNA-antagonism regulates breast cancer stemness and metastasis via TET-family-dependent chromatin remodeling[J]. Cell, 2013, 154(2):311-324.
[24]Yang J, Guo R, Wang H, et al. Tet enzymes regulate telomere maintenance and chromosomal stability of mouse ESCs[J]. Cell Rep, 2016, 15(8):1809-1821.
[25]Tian YP, Zhu YM, Sun XH, et al. Multiple functions of ten-eleven translocation 1 during tumorigenesis[J]. Chin Med J (Engl), 2016, 129(14):1744-1751.
[26]Zhao J, Ma XL, Ma JX, et al. TET3 mediates alterations in the epigenetic marker 5hmC and Akt pathway in ste-roid-associated osteonecrosis[J]. J Bone Miner Res, 2017, 32(2):319-322.
[27]Lian H, Li WB, Jin WL. The emerging insights into catalytic or non-catalytic roles of TET proteins in tumors and neural development[J]. Oncotarget, 2016, 7(39):64512-64525.
[28]Bachman M, Uribe-Lewis S, Yang X, et al. 5-Formylcytosine can be a stable DNA modification in mammals[J]. Nat Chem Biol, 2015, 11(8):555-557.
[29]Kriaucionis S, Heintz N. The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain[J]. Science, 2009, 324(5929): 929-930.
[30]Globisch D, Munzel M, Muller M, et al. Tissue distribution of 5-hydroxymethylcytosine and search for active demethylation intermediates[J]. PLoS One, 2010, 5(12):e15367.
[31]Ito S, Shen L, Dai Q, et al. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine[J]. Science, 2011, 333(6047): 1300-1303.
[32]Booth MJ, Branco MR, Ficz G, et al. Quantitative sequencing of 5-methylcytosine and 5-hydroxymethylcytosine at single-base resolution[J]. Science, 2012, 336(6083):934-937.
[33]Yu M, Hon GC, Szulwach KE, et al. Base-resolution analysis of 5-hydroxymethylcytosine in the mammalian genome[J]. Cell, 2012, 149(6): 1368-1380.
[34]Wang L, Zhang J, Duan J, et al. Programming and inhe-ritance of parental DNA methylomes in mammals[J]. Cell, 2014, 157(4):979-991.
[35]Sun Z, Dai N, Borgaro JG, et al. A sensitive approach to map genome-wide 5-hydroxymethylcytosine and 5-formylcytosine at single-base resolution[J]. Mol Cell, 2015, 57(4):750-761.
[36]Song CX, Szulwach KE, Dai Q, et al. Genome-wide profiling of 5-formylcytosine reveals its roles in epigenetic priming[J]. Cell, 2013, 153(3): 678-691.
[37]Booth MJ, Marsico G, Bachman M, et al. Quantitative sequencing of 5-formylcytosine in DNA at single-base resolution[J]. Nat Chem, 2014, 6(5):435-440.
[38]Plongthongkum N, Diep DH, Zhang K. Advances in the profiling of DNA modifications: cytosine methylation and beyond[J]. Nat Rev Genet, 2014, 15(10):647-661.
[39]Booth MJ, Raiber EA, Balasubramanian S. Chemical methods for decoding cytosine modifications in DNA[J]. Chem Rev, 2015, 115(6):2240-2254.
[40]Neri F, Incarnato D, Krepelova A, et al. Single-base resolution analysis of 5-formyl and 5-carboxyl cytosine reveals promoter DNA methylation dynamics[J]. Cell Rep, 2015, 10(5):674-683.
[41]Wu H, Zhang Y. Reversing DNA methylation: mechanisms, genomics, and biological functions[J]. Cell, 2014, 156(1-2):45-68.
[42]Wu H, Wu X, Shen L, et al. Single-base resolution ana-lysis of active DNA demethylation using methylase-assisted bisulfite sequencing[J]. Nat Biotechnol, 2014, 32(12):1231-1240.
[43]Zhou T, Xiong J, Wang M, et al. Structural basis for hydroxymethylcytosine recognition by the SRA domain of UHRF2[J]. Mol Cell, 2014, 54(5):879-886.
[44]Iurlaro M, Ficz G, Oxley D, et al. A screen for hydroxymethylcytosine and formylcytosine binding proteins suggests functions in transcription and chromatin regulation[J]. Genome Biol, 2013, 14(10):R119.
[45]Hashimoto H, Liu Y, Upadhyay AK, et al. Recognition and potential mechanisms for replication and erasure of cytosine hydroxymethylation[J]. Nucleic Acids Res, 2012, 40(11):4841-4849.
[46]Kellinger MW, Song CX, Chong J, et al. 5-formylcytosine and 5-carboxylcytosine reduce the rate and substrate specificity of RNA polymerase II transcription[J]. Nat Struct Mol Biol, 2012, 19(8):831-833.
[47]Wang L, Zhou Y, Xu L, et al. Molecular basis for 5-carboxycytosine recognition by RNA polymerase II elongation complex[J]. Nature, 2015, 523(7562): 621-625.
(責任編輯: 陳妙玲, 羅 森)
Progress in TET enzymes and their intermediate products
CHENG Yi-chen, WU Xiao-ming, LUO Ying
(MedicalFaculty,KunmingUniversityofScienceandTechnology,Kunming650500,China.E-mail:luoyingabc@yahoo.com)
DNA methylation is an important epigenetic modification mode, which plays a crucial role in gene expression, genome stability and development. DNA methylation is catalyzed and maintained in cell proliferation by the family of DNA methyltransferases. The ten-eleven translocation (TET) enzymes oxidize 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC), and 5-carboxylcytosine (5caC). Here, we briefly describe the TET enzymes and their role in cancer, and the distribution, the role and detection method of those three oxidation products of cytosine in genome.
TET酶; DNA去甲基化; 5-甲基胞嘧啶; 5-羥甲基胞嘧啶; 5-醛基胞嘧啶; 5-羧基胞嘧啶
TET enzymes; DNA demethylation; 5-methylcytosine; 5-hydroxymethylcytosine; 5-formylcytosine; 5-carboxylcytosine
1000- 4718(2017)03- 0572- 05
2016- 09- 18
2016- 10- 28
國家自然科學基金資助項目(No. KKGD201460063); 云南省教育廳科學研究基金資助項目(No.KKJA201560039)
△通訊作者 Tel: 13987631893; E-mail: luoyingabc@yahoo.com
R363.2+1
A
10.3969/j.issn.1000- 4718.2017.03.033
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
