細胞鐵死亡發生與調控機制的研究進展*
2017-04-11 04:39:31康傳杰張相彤
中國病理生理雜志 2017年3期
康傳杰, 張相彤, 馬 威
(哈爾濱醫科大學附屬第一醫院神經外科急診綠色通道,黑龍江 哈爾濱 150001)
細胞鐵死亡發生與調控機制的研究進展*
康傳杰, 張相彤△, 馬 威
(哈爾濱醫科大學附屬第一醫院神經外科急診綠色通道,黑龍江 哈爾濱 150001)
細胞死亡無論在生理還是在病理條件下都是無法避免的重要環節。傳統意義上細胞死亡主要分為凋亡(apoptosis)和壞死(necrosis)兩大主要類型,但近年來卻發現在諸多病理生理情況下亦有自噬(autophagy)、脹亡(oncosis)、副凋亡(paraptosis)、焦亡(pyroptosis)等多種方式參與其中。越來越多細胞死亡方式的發現為進一步明確疾病的發生機制以及為研究人員尋找新的疾病治療方法提供科學的理論依據。2012年Dixon等[1]首次發現并報道了這一新型細胞死亡方式——鐵死亡(ferroptosis),其確切機制尚有待深入研究。
本文將對鐵死亡的特征、調控機制及與其它細胞死亡方式的區別作一綜述,以期進一步加深對這一細胞死亡方式的認識,并對相關疾病靶向治療藥物的研發提供新思路。
1 鐵死亡的提出
鐵死亡是一種依賴于脂質過氧化反應驅動的非凋亡性細胞死亡方式;其發生過程需要細胞內富含可利用的鐵。2012年Dixon等[1]首次發現并報道了這一新型細胞死亡方式。第一個鐵死亡誘導復合物erastin和RSL3是通過小分子文庫高通量篩查發現的[1-2]。這些復合物誘導的細胞死亡是非凋亡性的,因為erastin和RSL3處理后細胞發生的死亡缺乏caspase激活等凋亡特征[3]。進一步研究發現,鐵螯合劑和一些親脂性抗氧化劑(如維生素E和β-胡蘿卜素)均能強烈抑制erastin和RSL誘導的這種細胞死亡,然而這種與鐵密切相關的死亡形式在細胞形態學、生物化學等方面與壞死、凋亡、自噬等均有明顯的不同。綜合這些發現,研究者提出這種與鐵含量增多相關的細胞死亡是一種先前未被認識的類型,并命名為鐵死亡[1, 4]。
2 鐵死亡的特征及與凋亡、自噬、焦亡的區別
鐵死亡是一個全新的細胞死亡模式,它在形態學、生物化學及遺傳學方面與凋亡、焦亡、自噬等有著明顯的不同,見圖1、表1。在形態學方面,發生鐵死亡的細胞主要表現為線粒體體積縮小,雙層膜密度增加,線粒體嵴減少或消失[1, 3]。在生化方面,谷胱甘肽(γ-glutamylcysteinylglycine,GSH)耗竭,谷胱甘肽過氧化物酶4(glutathione peroxidase 4,GPX4)活性下降,脂質氧化物不能經GPX4催化的谷胱甘肽還原反應代謝,繼而二價鐵離子以類似Fenton反應的方式氧化脂質產生大量活性氧簇(reactive oxygen species,ROS),促使細胞發生鐵死亡[3, 5]。在遺傳學方面,對于鐵死亡的發生及調控機制研究尚處于起始階段,目前發現鐵死亡是多基因調控的,主要涉及鐵穩態及脂質過氧化代謝等方面的基因改變。

Figure 1.Morphological characteristics of different cell death types. A: normal; B: ferroptosis (shrunken mitochondria); C: apoptosis (chromatin condensation); D: necrosis (cytoplasmic and organelle swelling,plasma membrane rupture); E: autophagy (formation of double-membrane vesicles)[1].
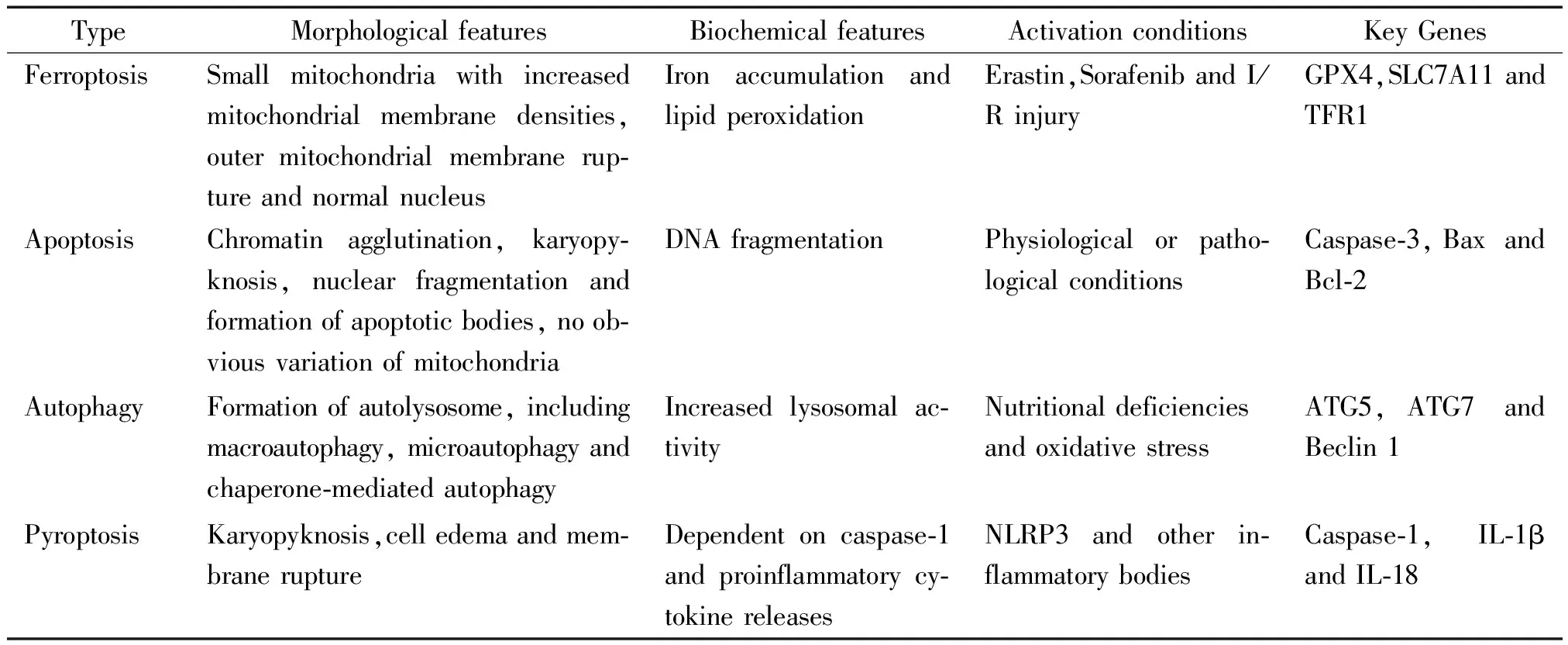
表1 鐵死亡、凋亡、自噬和焦亡的特征
凋亡是一種在生理性或病理性刺激下由基因調控的程序性細胞死亡(programmed cell death,PCD)。其形態學特征主要為:細胞皺縮,染色質濃縮,核裂解成碎片,之后細胞膜內陷將細胞分割形成多個具有膜包裹的凋亡小體。凋亡細胞及其凋亡小體由周圍吞噬細胞或鄰近的細胞清除,這一過程依賴caspase-3的活化,不發生線粒體、溶酶體及細胞膜的破裂,無內容物外溢,所以不引起炎癥反應[6]。
自噬分為大自噬、小自噬及分子伴侶介導的自噬,其形態學變化主要為:首先胞漿內包裹胞質及變性壞死細胞器的雙層膜結構形成,即自噬空泡;繼而與溶酶體膜相融合,形成自噬溶酶體;之后溶酶體中的水解酶水解自噬溶酶體內容物;最終自噬溶酶體破裂水解,水解產生的氨基酸等營養成分可被細胞再利用[7]。
焦亡是一種特殊的程序性細胞死亡,兼有凋亡和壞死的特點。其發生過程主要是由NLRP3(NACHT, LRR and PYD domains-containing protein 3)等炎癥小體活化及下游caspase-1的激活所介導[8]。
3 鐵內穩態的調控及意義
鐵參與體內多種重要的生理生化過程,包括氧的轉運,作為核糖核苷酸還原酶參與DNA的生物合成,作為線粒體電子傳遞鏈某些蛋白的輔助因子參與ATP的合成等。體內鐵穩態的維持需要一個非常復雜的調控過程,在哺乳動物體內鐵的主要儲存器官是肝臟,它能感知血清中鐵的含量,從而調節鐵代謝的核心激素——鐵調素的分泌,調控機體鐵穩態。鐵調素通過與小腸絨毛的上皮細胞、巨噬細胞和肝細胞膜上的膜鐵轉運蛋白(ferroportin,Fpn/SLC40A1)結合,介導Fpn的內化降解,進而調控十二指腸吸收鐵進入血液循環以及巨噬細胞循環再利用鐵,從而維持機體的鐵穩態[9]。Fpn是目前已知的唯一細胞內鐵流出泵。鐵調素的表達與血清及細胞內鐵的狀態有直接的關系,而細胞內鐵穩態主要受鐵代謝相關基因通過鐵反應元件-鐵調控蛋白系統進行轉錄后調控,如鐵蛋白(ferritin;包括重鏈FTH1和輕鏈FTL)、轉鐵蛋白(transferrin,TF)、轉鐵蛋白受體1、二價金屬離子轉運體1(divalent metal-ion transporter-1,DMT1/SLC11A2)等,以及近年來被發現的一些新的細胞內鐵攝取及利用的關鍵蛋白,如鋅鐵調控蛋白(ZRT/IRT-like proteins,ZIP)家族等[10]。
鐵是生命所必需的微量元素之一,鐵代謝失衡會引起多種病變的發生。如鐵能抑制富絲氨酸/精氨酸剪接因子7(serine/arginine-rich splicing factor 7,SRSF7)與細胞死亡受體Fas的mRNA前體結合,隨后外顯子6排除,導致突變的促凋亡的Fas亞型表達增加[11]。在遺傳性血色素沉著癥患者中,ZIP14可能是一個促進肝細胞死亡的因子,其通過介導鐵的轉運,增強促凋亡蛋白Fas亞型的表達發揮促凋亡作用[10]。
4 鐵死亡在相關疾病中的作用機制
目前已發現鐵死亡參與多種疾病的發生發展,且機制復雜,某些化合物如erastin或臨床藥物如柳氮磺胺吡啶、索拉菲尼均能誘導癌細胞和某些正常的細胞(如腎小管細胞、神經元、纖維母細胞、T細胞)觸發鐵死亡[12]。此過程是在鐵的參與下脂質過氧化產物和致命性反應ROS積聚導致的,在藥理學上能被鐵螯合劑去鐵胺以及脂質過氧化抑制劑ferrostatin-1、liproxstatin-1或zileuton所抑制(圖2)。GPX4、熱休克蛋白β-1(heat shock protein β-1,HSPB1)和核因子E2相關因子2(nuclear factor E2-related factor 2,NRF2)分別通過限制ROS產物和降低細胞內鐵的攝取對ferroptosis發揮負性調控作用(圖2)。
4.1 腫瘤 索拉菲尼廣泛用于晚期肝癌的治療,研究發現Rb蛋白狀態是索拉菲尼誘發肝癌細胞發生鐵死亡敏感性的一個重要參數,Rb蛋白缺失使癌細胞更容易發生鐵死亡[13]。另有研究表明核轉錄因子NRF2在肝細胞癌中有抑制鐵死亡發生的作用[14]。 NRF2的表達促進抗氧化相關基因NAD(P)H:醌氧化還原酶1[NAD(P)H:quinone oxidoreductase 1, NQO1]和血紅素加氧酶1(heme oxygenase-1,HO-1),以及鐵代謝相關基因(如FTH1)表達上調,繼而調控鐵代謝及脂質過氧化反應,抑制鐵死亡的發生(圖2)。而敲除NRF2及其靶基因能增強索拉菲尼的活性,抑制肝癌細胞增殖[14]。另外,對乙酰氨基酚過量是目前引起急性肝衰竭最常見的原因。另外對乙酰氨基酚已被證明可以誘導原代肝癌細胞發生鐵死亡,ferrostatin-1能抑制對乙酰氨基酚誘導的這種死亡[15]。
癌基因K-Ras重新編碼使胰導管腺癌細胞對凋亡產生強烈抵抗作用。在胰腺癌細胞株中青蒿酯特異性誘導ROS和溶酶體鐵依賴的死亡。而且通過分析胰腺癌病人mRNA表達情況發現癌細胞內存在抗氧化失衡以及細胞對游離的細胞內鐵敏感性增加。癌細胞獲得最強的細胞毒性需組成性激活型K-Ras的存在,但青蒿酯對非瘤型人類胰腺導管腺癌細胞株沒有影響[16]。
在鐵死亡敏感的纖維母細胞瘤細胞中轉鐵蛋白受體1表達上調,而鐵蛋白重鏈和鐵蛋白輕鏈表達下調[3](圖2),表明鐵貯備對鐵死亡的發生也有明顯影響。HO-1啟動血紅素分解代謝,釋放一氧化碳、鐵、膽綠素,用erastin孵育瘤細胞時,HO-1缺失的纖維肉瘤細胞死亡減少,而HO-1過表達加速癌細胞的死亡,進一步研究發現在erastin誘導的鐵死亡發生過程中HO-1可能通過增加細胞內的鐵及脂質過氧化發揮作用[17](圖2)。
NADPH氧化酶(NADPH oxidases,NOX)是細胞內一組具有氧化活性的蛋白,其主要的生物學功能是產生ROS。NOX抑制劑二甲基苯碘在人肺癌中能部分抑制erastin誘導的鐵死亡的發生[1]。磷酸戊糖途徑是與糖酵解產生NADPH和戊糖相平行的代謝途徑。利用6-氨基煙酰胺抑制磷酸戊糖途徑或者敲除磷酸戊糖途徑的2個關鍵酶葡萄糖-6-磷酸脫氫酶和磷酸甘油酸脫氫酶也能部分保護人肺癌細胞避免erastin誘導的鐵死亡的發生[1]。
在橫紋肌肉瘤中由于致瘤性的RAS突變體異常表達抑制了抗氧化防御系統對ROS的解毒作用,使橫紋肌肉瘤RMS13細胞對氧化應激產生抵抗,細胞對erastin誘導鐵死亡發生的耐藥性增加,這涉及多種不同的抗氧化防御通路,包括GSH系統、硫氧化還原蛋白系統等[18]。
在膠質瘤中低濃度柳氮磺胺吡啶不影響膠質瘤細胞的活性,但柳氮磺胺吡啶在高濃度時能誘發膠質瘤細胞發生鐵死亡,并緩解病灶周圍水腫。同時SAS能增強標準化療和自噬誘導劑替莫唑胺的療效[19]。
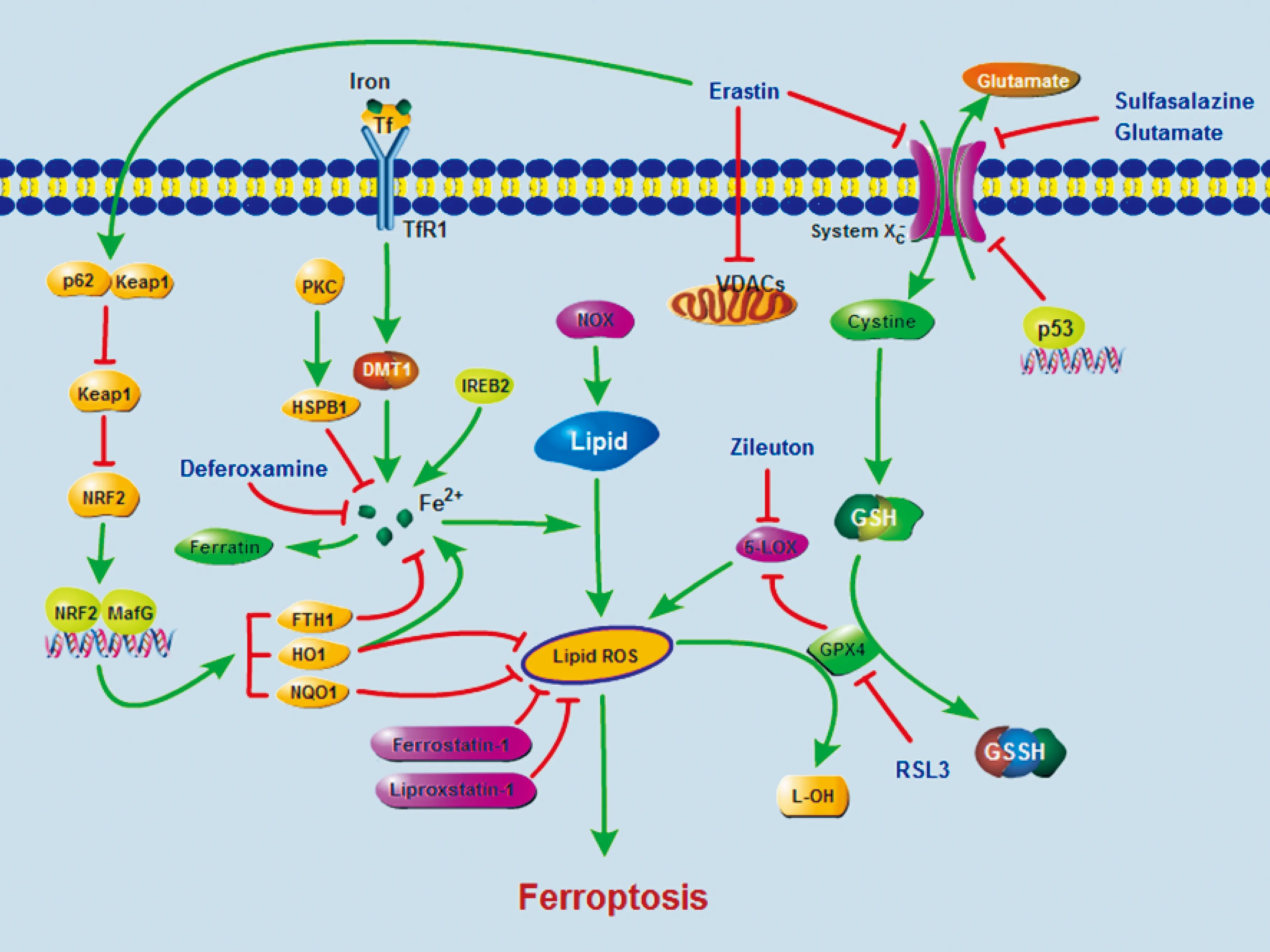
Figure 2. Ferroptosis-related signaling pathway.Ferroptosis is caused by iron-dependent lipid oxidative damage,which is mainly related to the regulation of iron metabolism and intracellular antioxidant ability. In the presence of sulfasalazine,glutamate and other conditions,the cystine /glutamate antiporter,system,is suppressed and then the content of GSH is reduced,which leads to the decrease in intracellular antioxidant capacity. In the meantime,the process of lipid peroxidation is enhanced in the existence of iron ion,and lipid ROS increases. The iron-dependent cell death,ferroptosis,is triggered eventually.
4.2 神經系統 包括膠質瘤在內的神經系統抗腫瘤藥物療效的發揮也與鐵死亡相關[19]。利用谷氨酸培養HT22海馬神經元細胞發現細胞死亡數量顯著升高,5-脂氧合酶抑制劑zileuton能部分通過抑制鐵死亡的發生而保護海馬神經元細胞免受谷氨酸氧化毒性損害[20]。
腦室周圍白質軟化通常是由于發育中的少突膠質細胞死亡所致。研究顯示在本病中少突膠質細胞的死亡是鐵依賴性的,并伴隨脂質ROS的升高[21]。Ferrostatins(如ferrostatin-1)能有效保護少突膠質細胞免受胱氨酸耗竭的損害[22]。
在1-甲基-4-苯基-1,2,3,6-四氫吡啶(1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine,MPTP)誘導的帕金森病動物模型中,MPTP對多巴胺能神經元的毒性作用能被ferrostatin-1抑制,進一步研究發現在多巴胺能神經元中蛋白激酶Cα(protein kinase Cα,PKCα)激活,繼而活化絲裂原激活蛋白激酶(mitogen-activated protein kinase,MAPK),并以Ras非依賴的方式觸發鐵死亡。在神經退行性變中鐵螯合劑、ferrostatin-1和PKC抑制劑對鐵死亡發生的信號通路均具有強效抑制作用[23]。
而在周圍神經系統,敲除GPX4能引起脂質過氧化及線粒體功能紊亂從而使快速運動神經元發生鐵死亡,導致小鼠癱瘓,而對成年大鼠皮質神經元的影響相對較小[24]。
4.3 缺血再灌注損傷 Ferrostatins在急性腎損傷模型新鮮孤立腎小管中起到保護作用,阻止發生致命性損害[22],這表明在急性腎衰竭中可能存在鐵死亡介導的細胞死亡。利用第3代ferrostatins抑制鐵死亡能削弱缺血再灌注損傷引起的急性腎衰[25]。
谷氨酰胺代謝被認為是鐵死亡發生的重要因素之一,利用野生型小鼠離體心臟模型使其發生缺血再灌注應激反應,在再灌注開始時用去鐵胺處理離體心臟,發現與對照組相比去鐵胺組心臟的心功能有明顯增強。再灌注期間去鐵胺能顯著抑制心臟損傷另一指標乳酸脫氫酶的釋放。這些數據表明通過控制谷氨酰胺代謝進而抑制鐵死亡的發生緩解缺血再灌注損傷對離體心臟起保護作用[26]。
另外小鼠肝缺血損害也與鐵死亡相關,鐵死亡抑制劑liproxstatin-1可完全阻斷脂質過氧化作用,改善小鼠缺血再灌注引起的肝損傷[5]。
5 展望
目前研究發現許多疾病的發生發展都涉及多種不同的細胞死亡類型,如壞死、凋亡、自噬等。然而針對這些過程的靶向治療效果卻不盡如人意,表明其它未被發現的死亡過程在這些病理過程可能同樣起到十分重要的作用。研究結果顯示鐵死亡在多種癌癥、神經毒性、肝腎損害等病變的演變過程中發揮重要作用。鐵死亡的研究尚處于初期階段,尚存在大量未解決的問題,如具體哪些脂類物質被氧化?體內其它代謝過程是否存在鐵死亡的發生等都需要更進一步的研究。對鐵死亡的深入研究將有助于更加充分認識多種疾病的發生進展機理并為臨床防治提供新思路。
[1] Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death[J]. Cell, 2012, 149(5):1060-1072.
[2] Dolma S, Lessnick SL, Hahn WC, et al. Identification of genotype-selective antitumor agents using synthetic lethal chemical screening in engineered human tumor cells[J]. Cancer Cell, 2003, 3(3):285-296.
[3] Yang WS, Stockwell BR. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells[J]. Chem Biol, 2008, 15(3):234-245.
[4] Yang WS, Stockwell BR. Ferroptosis: death by lipid peroxidation[J]. Trends Cell Biol, 2016, 26(3):165-176.
[5] Friedmann Angeli JP, Schneider M, Proneth B, et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice[J]. Nat Cell Biol, 2014, 16(12):1180-1191.
[6] 周彩虹, 黃啟福. 凋亡與腫瘤及其治療進展[J]. 中國病理生理雜志,2004,20(11):2124-2133.
[7] 洪 遠, 郭松雪, 張建民. 自噬在創傷性腦損傷中的作用[J]. 中國病理生理雜志,2011,27(8):1652-1656.
[8] Galluzzi L, Vitale I, Abrams JM, et al. Molecular definitions of cell death subroutines: recommendations of the Nomenclature Committee on Cell Death 2012[J]. Cell Death Differ, 2012, 19(1):107-120.
[9] 安 鵬. 鐵代謝穩態失衡Hepcidin調控機制研究[D]. 上海:華東理工大學, 2014.
[10]Bogdan AR, Miyazawa M, Hashimoto K, et al. Regulators of iron homeostasis: new players in metabolism, cell death, and disease[J]. Trends Biochem Sci, 2016, 41(3):274-286.
[11]Tejedor JR, Papasaikas P, Valcarcel J. Genome-wide identification of Fas/CD95 alternative splicing regulators reveals links with iron homeostasis[J]. Mol Cell, 2015, 57(1):23-38.
[12]Xie Y, Hou W, Song X, et al. Ferroptosis: process and function[J]. Cell Death Differ, 2016, 23(3):369-379.
[13]Louandre C, Marcq I, Bouhlal H, et al. The retinoblastoma (Rb) protein regulates ferroptosis induced by sorafenib in human hepatocellular carcinoma cells[J]. Cancer Lett, 2015, 356(2 Pt B):971-977.
[14]Sun X, Ou Z, Chen R, et al. Activation of the p62-Keap1-NRF2 pathway protects against ferroptosis in hepatocellular carcinoma cells[J]. Hepatology, 2016, 63(1):173-184.
[15]Lorincz T, Jemnitz K, Kardon T, et al. Ferroptosis is involved in acetaminophen induced cell death[J]. Pathol Oncol Res, 2015, 21(4):1115-1121.
[16]Eling N, Reuter L, Hazin J, et al. Identification of artesunate as a specific activator of ferroptosis in pancreatic cancer cells[J]. Oncoscience, 2015, 2(5):517-532.
[17]Kwon MY, Park E, Lee SJ, et al. Heme oxygenase-1 accelerates erastin-induced ferroptotic cell death[J]. Oncotarget, 2015, 6(27):24393-24403.
[18]Schott C, Graab U, Cuvelier N, et al. Oncogenic RAS mutants confer resistance of RMS13 rhabdomyosarcoma cells to oxidative stress-induced ferroptotic cell death[J]. Front Oncol, 2015, 5:131.
[19]Sehm T, Fan Z, Ghoochani A, et al. Sulfasalazine impacts on ferroptotic cell death and alleviates the tumor microenvironment and glioma-induced brain edema[J]. Oncotarget, 2016, 7(24):36021-36033.
[20]Liu Y, Wang W, Li Y, et al. The 5-lipoxygenase inhibitor zileuton confers neuroprotection against glutamate oxidative damage by inhibiting ferroptosis[J]. Biol Pharm Bull, 2015, 38(8):1234-1239.
[21]Welin AK, Svedin P, Lapatto R, et al. Melatonin reduces inflammation and cell death in white matter in the mid-gestation fetal sheep following umbilical cord occlusion[J]. Pediatr Res, 2007, 61(2):153-158.
[22]Skouta R, Dixon SJ, Wang J, et al. Ferrostatins inhibit oxidative lipid damage and cell death in diverse disease models[J]. J Am Chem Soc, 2014, 136(12):4551-4556.
[23]Do Van B, Gouel F, Jonneaux A, et al. Ferroptosis, a newly characterized form of cell death in Parkinson’s di-sease that is regulated by PKC[J]. Neurobiol Dis, 2016, 94:169-178.
[24]Chen L, Hambright WS, Na R, et al. Ablation of the ferroptosis inhibitor glutathione peroxidase 4 in neurons results in rapid motor neuron degeneration and paralysis[J]. J Biol Chem, 2015, 290(47):28097-28106.
[25]Linkermann A, Skouta R, Himmerkus N, et al. Synchronized renal tubular cell death involves ferroptosis[J]. Proc Natl Acad Sci U S A, 2014, 111(47):16836-16841.
[26]Gao M, Monian P, Quadri N, et al. Glutaminolysis and transferrin regulate ferroptosis[J]. Mol Cell, 2015, 59(2):298-308.
(責任編輯: 陳妙玲, 羅 森)
Progress in occurrence and development of ferroptosis
KANG Chuan-jie, ZHANG Xiang-tong, MA Wei
(DepartmentofEmergencyandNeurosurgery,TheFirstAffiliatedHospital,HarbinMedicalUniversity,Harbin150001,China.E-mail:zgxgtg@sina.com)
With the development of modern medical research, it has been found that there are many forms of cell death under lots of physiological or pathological conditions. Among them, necrosis and apoptosis are the 2 main forms of cell death. In recent years, varieties of new cell death patterns, such as autophagy, oncosis and paraptosis, have been discovered. In 2012, Dixon et al first discovered and reported a new form of iron-dependent cell death, ferroptosis. In this review, we try to expound the morphological and biochemical features of ferroptosis and its related diseases in detail, so as to deepen our understanding of cell death, thus providing new ideas for the prevention and treatment of clinically relevant diseases.
細胞死亡; 鐵死亡; 鐵; 脂質; 活性氧簇
Cell death; Ferroptosis; Iron; Lipids; Reactive oxygen species
1000- 4718(2017)03- 0567- 05
2016- 09- 05
2016- 11- 03
國家自然科學基金資助項目(No. 81571108)
△通訊作者 Tel: 0451-85555109; E-mail: zgxgtg@sina.com
R363
A
10.3969/j.issn.1000- 4718.2017.03.032
