一個預測的木薯衰老相關基因的獲得及其功能分析
2017-04-27 00:49:27李超萍蔡吉苗李博勛黃貴修
廣東農業科學 2017年2期
時 濤,李超萍,蔡吉苗,李博勛,黃貴修
(中國熱帶農業科學院環境與植物保護研究所/農業部熱帶作物有害生物綜合治理重點實驗室/海南省熱帶農業有害生物監測與控制重點實驗室,海南 海口 571101)
一個預測的木薯衰老相關基因的獲得及其功能分析
時 濤,李超萍,蔡吉苗,李博勛,黃貴修
(中國熱帶農業科學院環境與植物保護研究所/農業部熱帶作物有害生物綜合治理重點實驗室/海南省熱帶農業有害生物監測與控制重點實驗室,海南 海口 571101)
根據水稻LRR類抗病基因Xa21的保守結構域設計1對簡并引物,提取木薯抗/感細菌性萎蔫病種質E1340和GR911的基因組DNA為模板,通過PCR擴增均獲得大小約0.5 kb的擴增片段。兩個片段克隆、測序后獲得完全一致的474 nt序列,比對發現木薯種質AM560帶有與該序列高度同源的核苷酸片段。提取包括同源片段上下游各約2.2 kb的大片段序列進行基因預測,發現該片段位于1個預測基因內,命名為SLR1。該預測基因ORF全長1 641 nt,編碼546 aa,與已報道的衰老相關蛋白具有較高的同源性,可能在木薯衰老相關反應中發揮重要作用。
木薯;衰老相關基因;功能分析
木薯(Manihot esculenta crantz)為大戟科(Euphorbiaceae)木薯屬灌木狀多年生作物,起源于熱帶美洲地區[1]。木薯具有粗生易長的特點,目前已在全球100多個國家和地區廣泛種植,是約10億人口的主食。據世界糧農組織(FAO)統計,近年來國際木薯總產量穩步上升,接近3億t[2]。1820年前后,木薯從東南亞傳入我國廣東地區,目前已在華南地區廣泛種植,北京、河北、山東地區也進行了引種試驗。木薯在我國最初供食用或做飼料,目前主要用作工業原料,可生產變性淀粉、乙醇等大量產品[3]。據國家木薯產業體系的統計,2014年我國木薯種植面積約39.27萬hm2,鮮薯總產量893.63萬t[4]。隨著我國木薯加工業的發展,國內木薯產量已經不能滿足產業所需,每年均需從東南亞和非洲等地區進口大量的木薯干片及淀粉,我國是世界上最大的木薯進口國[5]。
病害防控是木薯種植中的重要問題之一。世界范圍內木薯病害有30多種[6],我國有3類9種,其中細菌性萎蔫病為害最為嚴重,其次為褐斑病和疫霉根腐病[7-9]。選用抗病品種能夠有效地減輕病害發生,是常用的防控措施之一。目前國內外有關木薯抗細菌性萎蔫病方面的研究較少,也并未獲得可用的基因資源,嚴重影響了抗病育種工作的進行。在植物抗病反應中,相關基因調控著一系列級聯反應,發揮著核心作用,大量抗病基因已得到克隆和功能鑒定[10]。研究發現,絕大多數已克隆的抗病基因編碼產物均具有保守結構域,根據該類保守域設計引物進行抗病基因類似序列(Resistance gene analogs,RGAs)的分離,再從中篩選潛在的抗病基因,稱為類似序列法,近年來在相關研究中得到廣泛應用。本研究根據水稻抗病基因Xa21的保守區設計簡并引物,從木薯中分離了一個預測的衰老相關基因。
1 材料與方法
1.1 試驗材料
1.1.1 供試木薯品種 木薯種質E1340和GR911,由中國熱帶農業科學院熱帶作物品種資源研究所木薯種質資源圃提供,盧昕等[11]已證明E1340為抗細菌性萎蔫病種質,GR911為高感種質。
1.1.2 培養基和試劑 Taq酶、dNTPs、大腸桿菌JM109感受態細胞、DNA 片段膠回收試劑盒、pMD19-T 載體,購自寶生物工程(大連)有限公司。引物由北京六合華大基因科技股份有限公司合成。大腸桿菌培養采用LB培養基(參照方中達[12]的方法制備),其他試劑均為國產分析純。
1.2 試驗方法
1.2.1 木薯基因組DNA的提取及目的序列擴增 田間采集健康木薯葉片,參照閆慶祥等[13]的方法提取基因組DNA。根據水稻抗白葉枯基因Xa21的保守結構域設計1對簡并引物Pl-F (SYGTWGGAYARGWRGGAG-3′)和Pl-R (5′- AACARTCCRR YMYWTGGT-3′),參照王潔[14]的反應體系和參數進行PCR擴增。擴增產物經電泳檢測后,按說明書進行產物的回收和克隆,隨機挑選3個陽性克隆送北京六合華大基因科技股份有限公司進行測序。
1.2.2 目的基因預測和功能分析 參照李超萍等[15]的方法進行。
2 結果與分析
2.1 木薯基因組DNA的提取及目的序列擴增
提取木薯抗病種質E1340和感病種質GR911基因組作為模板,分別用引物對Pl-F和Pl-R進行PCR擴增,均獲得大小約0.5 kb的擴增產物(圖1)。回收、克隆、測序后發現兩個擴增產物的核苷酸序列完全一致,長度均為474 nt。
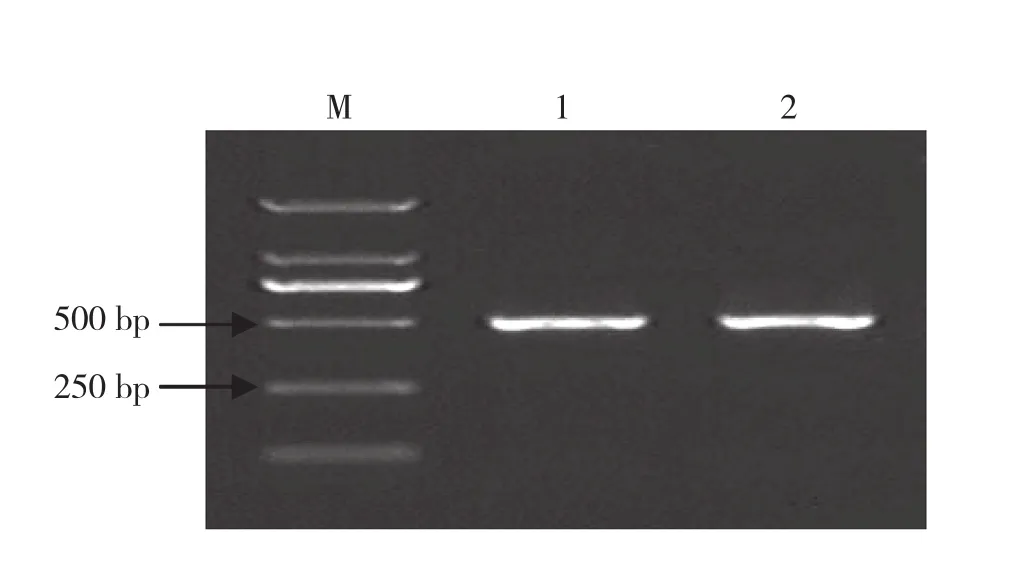
圖1 引物對Pl-F和Pl-R的PCR擴增結果
2.2 目的基因的預測和功能分析
2.2.1 序列的同源性比對 采用Blastx對所獲序列進行同源性比對,結果表明其與已報道的蒺藜苜蓿、柏樹、豌豆、百合等植物的衰老相關蛋白的同源性在50%以上(表1)。

表1 所獲序列同源性比對結果
2.2.2 目的基因預測 將所獲序列和木薯種質AM560的基因組序列進行比對,序列一致性為99%的同源片段,獲得包括該片段上下游各約2.2 kb的大片段序列,采用Softberry(http://www.softberry.com)的Hevea(橡膠樹)、Arabidopsis(擬南芥)2種模式和GENSCAN (http://genes.mit.edu/GENSCAN.html)的Maize(玉米)模式進行基因預測,結果(圖2)均表明該同源片段位于一個預測基因內部,該基因暫命名為SLR1。參照同屬大戟科作物的Hevea模式分析結果,該基因位于負鏈上,有10個外顯子,TSS(轉錄起始位點)位于第1個外顯子前約0.2 kb處,ORF全長1 641 nt,編碼546 aa。
2.2.3 目的基因保守結構域分析 利用SMART軟件(http://smart.embl-heidelberg.de/)、NCBI 中CDD數據庫和Sanger中Pfam數據庫對SLR1基因結構進行預測,發現該基因蛋白產物不具有LRR類抗病基因特有的結構域(圖3)。
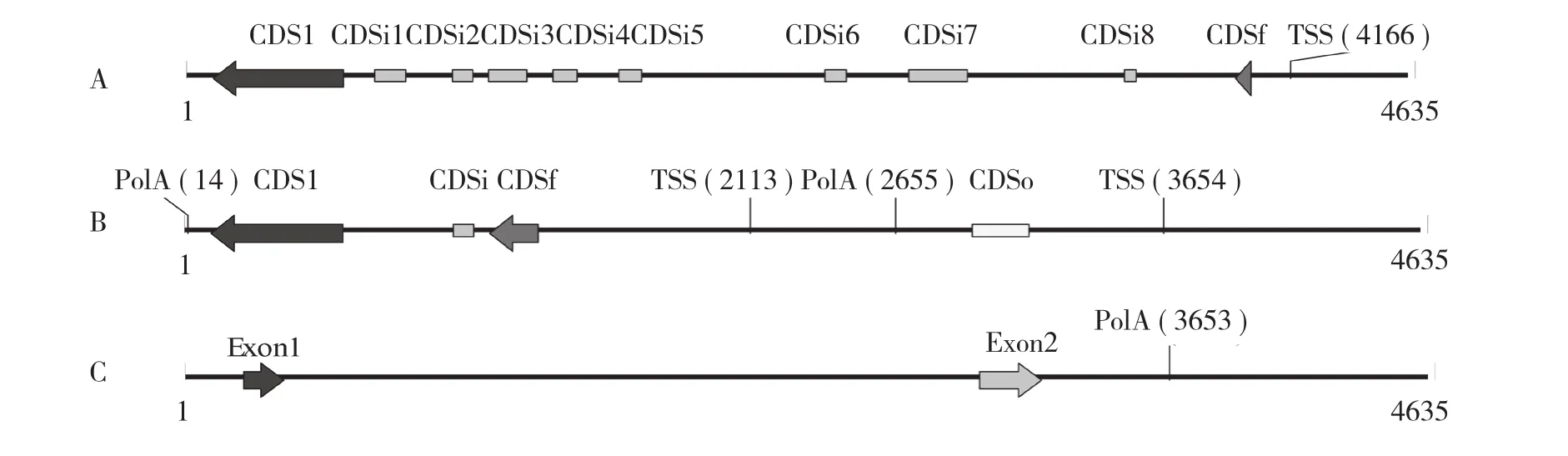
圖2 幾種預測模式對目的基因SLR1的預測結果
2.2.4 目的基因蛋白產物結構分析 利用ProtParam(http://web.expasy.org/protparam/)分析發現該蛋白質分子量為59.16 ku,等電點為9.19,平均疏水性(GRAVY)為-0.086。各氨基酸中,絲氨酸、亮氨酸和丙氨酸的含量最高,分別為65個(9.9 %)、57個(10.84 %)和56個(7.23 %),色氨酸和谷氨酰胺含量最低,分別為7個(2.07 %)和8個(1.69 %)。TMpred程序(http://www.ch.embnet.org/software/ TMPRED_form.html )預測結果表明,SLR1所編碼蛋白在第3~24、80~100、219~241、241~262、306~326、335~359、352~373個氨基酸殘基間存在7個由內到外的跨膜螺旋區域,在第3~15、81~100、219~239、239~262、306~329、356~373、440~457個氨基酸殘基間存在7個由外到內的跨膜螺旋區域(圖4)。
2.2.5 目的基因蛋白產物的聚類分析 氨基酸序列比對結果表明,SLR1基因和來自蒺藜苜蓿的幾個預測衰老相關蛋白具有較高的同源性,其中XP_013443002.1的同源性最高,為49%。與部分近源蛋白序列進行聚類分析,結果表明來自蒺藜苜蓿的6個預測衰老相關蛋白XP_013443002.1、XP_013442995.1、XP_013442969.1、XP_013442966.1、XP_013442997.1和XP_013442963.1為一個分枝,與SLR1同源性最高;來自大豆的預測蛋白GLYMA_13G019300(KRH17828.1)和花生的預測蛋白(XP_016164677.1)為親緣關系較近的一個分枝,而來自百合的預測衰老相關蛋白(ABO20851.1)和來自柏樹的預測衰老相關蛋白(ACA30301.1)為親緣關系最遠的一個分枝(圖5)。
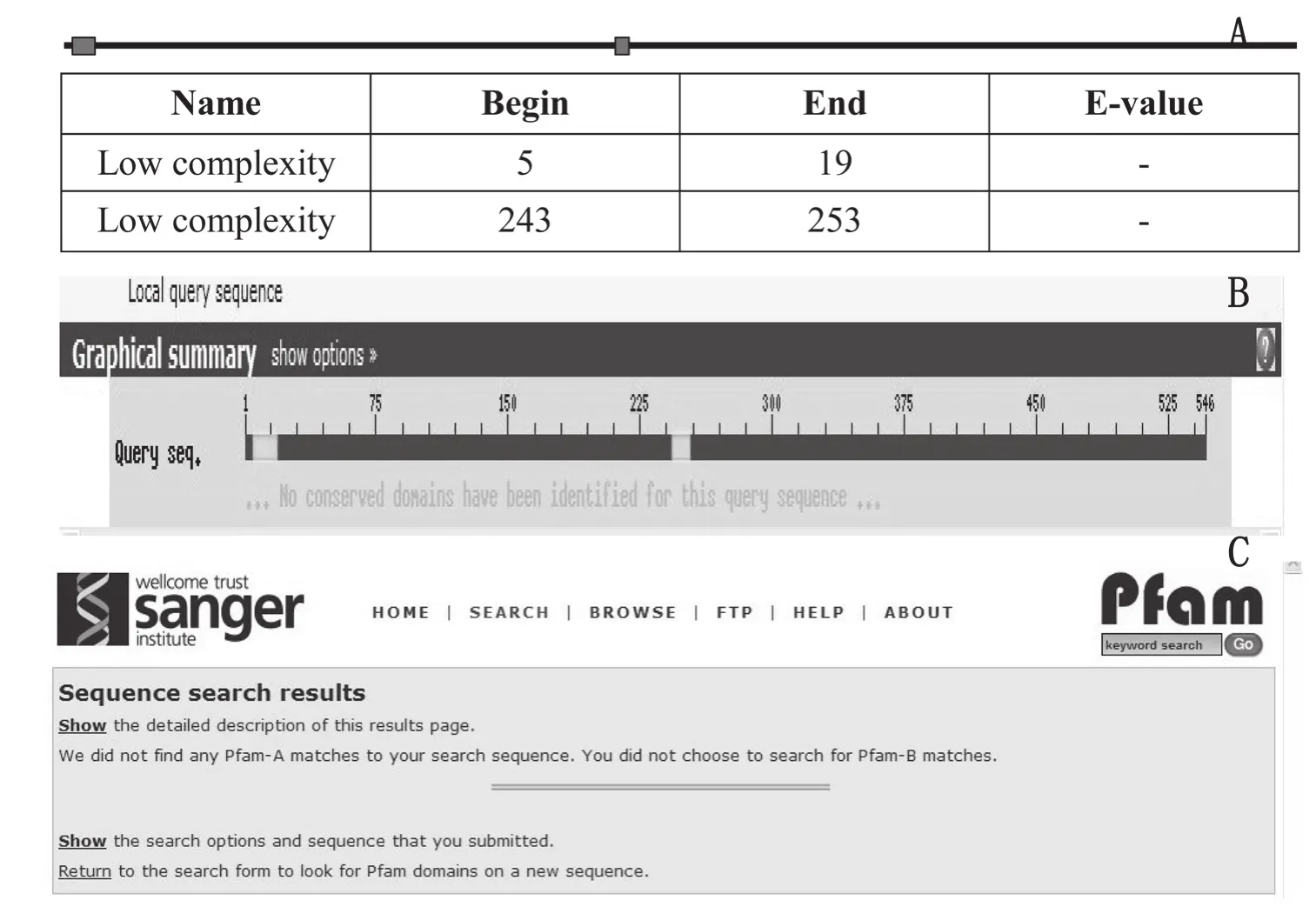
圖3 利用SMART(A)、NCBI中CDD數據庫(B)和Sanger中Pfam數據庫(C))分析SLR1基因結構結果

圖4 SLR1跨膜區域預測
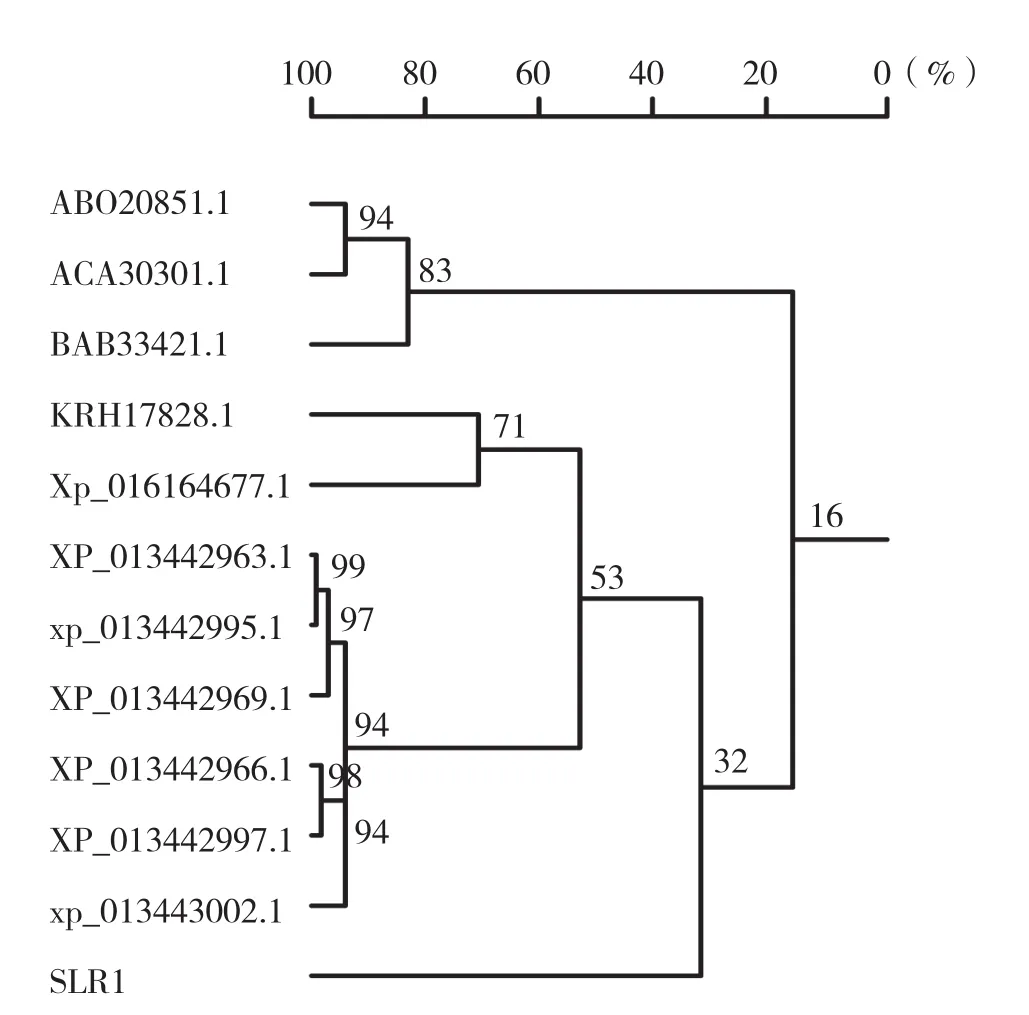
圖5 SLR1基因氨基酸序列的聚類分析
3 結論與討論
本研究根據水稻抗白葉枯基因Xa21的保守結構域設計引物,分別以抗、感細菌性萎蔫病的木薯種質E1340、GR911的DNA為模板,均擴增得到完全一致的474 nt序列。該序列和木薯種質AM560的基因組序列比對后獲得一個預測基因SLR1,其ORF全長1 641 nt,編碼546 aa。該基因蛋白產物不具有LRR類抗病基因特有的結構域,絲氨酸、亮氨酸和丙氨酸的含量最高,色氨酸和谷氨酰胺含量最低,預測有14個跨膜結構域。氨基酸序列比對和聚類分析表明,該基因和預測的衰老相關蛋白有較高的同源性。本研究發現3個木薯種質中均帶有SLR1或其同源基因,表明該基因可能在衰老相關反應中發揮重要作用。
目前,采用類似序列法從植株中分離獲得大量的RGAs,很多是抗病基因的一部分或者與其是緊密連鎖的。例如亞麻的4個RGAs位于抗條誘病復合N基因簇[16],是抗病基因的一部分,而核桃中的部分NBS類RGAs僅存在于抗炭疽病核桃品種中,與抗病性相關[17]。Elízabeth等[18]從木薯中分離得到兩個RGAs,與其木薯抗細菌性萎蔫病QTL連鎖,其中1個在受到病原侵染5 d后表達,而另外1個是組成性表達的。李超萍等[15]也從木薯中獲得1個具有NBS類抗病基因保守區的預測抗病基因。有些RGAs和抗病反應不相關,本研究所獲預測基因SLR1不具備抗病基因的結構特征,而是和衰老相關蛋白具有較高的同源性。
衰老是植物生長發育的最后一個階段,是植株在細胞、組織、器官或整株水平上生長衰退的過程,受基因調控并受內外環境因素影響[19]。葉片衰老是植物衰老的主要表現形式,是由基因控制的細胞自主有序死亡的過程[20],該階段葉片的主要功能是將生物大分子降解形成的營養元素輸送至新生器官,供進一步生長發育或儲存[21-22]。衰老和抗病反應之間是密切相關的,例如衰老相關蛋白在菊花受白銹病[23]、玉米受立枯絲核菌[24]、小麥受條銹病[25]、苧麻受根腐線蟲[26]侵染后,表達量均出現了差異變化。SLR1基因是否通過參與衰老相關反應而調控木薯和細菌性萎蔫病的互作機理有待進一步深入研究。
[1]黃潔,周建國. 木薯間套作與高效利用技術[M]. 海口:海南出版社,2015.
[2]Food and agriculture organization of the united Nations. Food outlook,biannual report on global food markets[Z]. 2014.
[3]陳麗珍,葉劍秋. 我國木薯加工業的發展現狀與展望[J]. 園藝與種苗,2011(3):87-90.
[4]國家木薯產業技術體系. 木薯產業技術發展報告[Z]. 2014.
[5]盤歡. 亞洲11 國木薯生產概況[J]. 廣西熱帶農業,2009(5):29-31.
[6]李開綿,黃貴修. 木薯主要病蟲害[M]. 北京:中國農業科學技術出版社,2008.
[7]李超萍,時濤,劉先寶,等. 國內木薯病害普查及細菌性萎蔫病安全性評估[J]. 熱帶作物學報,2011,32(1):116-121.
[8]盧昕,李超萍,裴月令,等. 木薯疫霉根腐病病原初步鑒定及其生物學特性測定[J]. 熱帶農業科學,2014,34(8):59-62,85.
[9]陳奕鵬,時濤,蔡吉苗,等. 木薯新發藻斑病在中國的發生調查及病原鑒定[J]. 熱帶作物學報,2016,37(9):1787-1792.
[10]董繼新,董海濤,李德葆. 植物抗病基因研究進展[J]. 植物病理學報,2001,31(1):1-9.
[11]盧昕,李超萍,時濤,等. 國內603份木薯種質對細菌性枯萎病抗性評價[J]. 熱帶農業科學,2013,33(4):67-70,90.
[12]方中達. 植病研究方法[M]. 第3版. 北京:中國農業出版社,1998.
[13]閆慶祥,黃東益,李開綿,等. 利用改良CTAB法提取木薯基因組DNA[J]. 中國農學通報,2010,26(4):30-32.
[14]王潔. 水稻白葉枯病抗性相關基因及抗病基因類似序列的克隆[D]. 北京:中國農業大學,2005.
[15]李超萍,樊春俊,時濤,等. 木薯一個假定的NBS類抗病基因的獲得及其功能分析[J]. 熱帶農業科學,2013,33(12):35-40.
[16]Dodds P N,Lawrence C X J,Ellis J C X. Contrasting modes of evolution acting on the complex N locus for rust resistance in flax[J]. Plant J. ,2001,27 (5):439-453.
[17]安海山,楊克強. 核桃NBS類抗病基因類似物的序列特征及其與炭疽病的抗性[J]. 中國農業科學,2014,47(2):344-356.
[18]E lí zabeth C N,Camilo L C. Expression of two resistance gene candidates against cassava bacterial blight in cassava[J]. Acta biol. Colomb.,2008,13(2):175-188.
[19]Norbert Brugiè re,Frédéric Dubois,Céline Masclaux,et al. Immunolocalization of glutamine synthetase in senescing tobacco(Nicotiana tabacum L.)leaves suggests that ammonia assimilation is progressively shifted to the mesophyll cytosol[J]. Planta,2000,211(4):519-527.
[20]楊征,蔡陳崚,宋運淳.植物細胞凋亡研究進展[J].生物化學與生物物理進展,1999,26 (5):439-443.
[21]Izumi M,Hidema J,Makino A,et al. Autophagy contributes to nighttime energy availability for growth in Arabidopsis[J]. Plant Physiol,2013,161(4):1682-1693.
[22]Tegeder M. Transporters involved in source to sink partitioning of amino acids and ureides:opportunities for crop improvement[J]. J Exp Bot,2014,65(7):1865-1878.
[23]王順利. 菊花白銹病綜合防治及菊花抗病相關基因研究[D]. 北京:北京林業大學,2008.
[24]馬永毅. 立枯絲核菌AG1-IA誘導玉米基因差異表達分析及防衛酶活性檢測[D]. 雅安:四川農業大學,2007.
[25]萬翠萍. 小麥應答條銹病菌冬孢子產生的蛋白質組學分析[D]. 楊凌:西北農林科技大學,2016.
[26]劉慧玲. 苧麻根腐線蟲誘導的SSH文庫構建及EST分析[D]. 北京:中國農業科學院,2013.
(責任編輯 崔建勛)
Obtaining and function analysis of predicted senescence-associated gene from cassava
SHI Tao,LI Chao-ping,CAI Ji-miao,LI Bo-xun,HUANG Gui-xiu
(Environment and Plant Protection Institute,Chinese Academy of Tropical Agricultural Sciences/ Key Laboratory of Integrated Pest Management on Tropical Crops,Ministry of Agriculture/Hainan Key Laboratory for Monitoring and Control of Tropical Agricultural Pests,Haikou 571101,China)
One pair of degenerated primers was designed based on the conserved domain of resistance gene Xa21,which had LRR-liked domain and came from rice. The genome DNA of cassava germplasm E1340 and GR911 was extracted and used as template,which was resistant and susceptible to cassava bacterial blight separately. Two products of about 0.5 kb were obtained by PCR reaction. These two products were purified,cloned and sequenced,and then the same sequences of 474 nt were got. This sequence was highly matched with the homology segment anchored on the genome of cassava germplasm AM560. The long segment including each about 2.2 kb sequences from upstream and downstream of the homology segment was obtained,then one gene was predicted and named SLR1. Sequence analysis results of SLR1 showed that the ORF included 1 641 nucleotides encoding 546 amino acids. This predicted protein was higher consistent with senescence-associated protein reported,may play an important role in senescence mechanism of cassava.
cassava;senescence-associated gene;function analysis
Q781
A
1004-874X(2017)02-0006-06
2016-10-21
農業部現代農業人才支撐計劃項目(0316001);國家木薯產業技術體系建設項目(CARS-12-hnhgx);海南省自然科學基金(20153046)
時濤(1977-),男,博士,副研究員,E-mail:shitaofly2008@163.com
黃貴修(1968-),男,博士,研究員,E-mail:hgxiu@vip.163.com
時濤,李超萍,蔡吉苗,等.一個預測的木薯衰老相關基因的獲得及其功能分析[J].廣東農業科學,2017,44(2):6-11.
