病毒誘導(dǎo)的全新 tae-miR9663的沉默影響葉片發(fā)育和籽粒大小
2017-05-15 07:08:30朱妍峰趙惠賢
麥類作物學(xué)報(bào) 2017年4期
朱妍峰,王 倩,趙惠賢,2
(1.西北農(nóng)林科技大學(xué)生命科學(xué)學(xué)院,陜西楊陵 712100; 2.西北農(nóng)林科技大學(xué)旱區(qū)作物逆境生物學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室,陜西楊陵 712100)
病毒誘導(dǎo)的全新 tae-miR9663的沉默影響葉片發(fā)育和籽粒大小
朱妍峰1,王 倩1,趙惠賢1,2
(1.西北農(nóng)林科技大學(xué)生命科學(xué)學(xué)院,陜西楊陵 712100; 2.西北農(nóng)林科技大學(xué)旱區(qū)作物逆境生物學(xué)國(guó)家重點(diǎn)實(shí)驗(yàn)室,陜西楊陵 712100)
MicroRNA(miRNA)是一類長(zhǎng)度為18~24 nt的非編碼小分子RNA,參與植物各種發(fā)育進(jìn)程。 tae-miR9663是新發(fā)現(xiàn)的在小麥幼苗、旗葉和籽粒中高表達(dá)的miRNA,但其生物學(xué)功能未知。為了探索 tae-miR9663的功能,通過(guò)人工合成 tae-miR9663的小串聯(lián)模擬靶標(biāo)(short tandem target mimic, STTM),將其構(gòu)建到大麥條斑花葉病毒(barley stripe mosaic virus, BSMV)載體上,利用病毒介導(dǎo)的基因沉默(virus-inducing gene silencing, VIGS)技術(shù)轉(zhuǎn)染小麥寧春16五葉一心期的第5片葉,轉(zhuǎn)染20 d后觀察葉片表型并取旗葉進(jìn)行實(shí)時(shí)定量PCR,成熟時(shí)觀察種子大小。葉片表型觀察結(jié)果表明,與BSMV:00相比,接種BSMV:STTM- tae-miR9663的9株幼苗中出現(xiàn)4種葉片表型,即第6片葉有白點(diǎn)或白條紋,旗葉(第7片葉)有白點(diǎn)或白條紋,第6片葉邊緣有鋸齒狀,旗葉葉尖處有皺縮。實(shí)時(shí)定量PCR分析結(jié)果表明,STTM- tae-miR9663過(guò)表達(dá)植株的 tae-miR9663表達(dá)豐度下降,說(shuō)明BSMV-VIGS技術(shù)可通過(guò)過(guò)表達(dá)STTM有效地沉默內(nèi)源miRNA。成熟種子大小觀察結(jié)果表明,與BSMV:00比較,接種BSMV:STTM- tae-miR9663的植株種子的長(zhǎng)和寬均減小。
小麥; tae-miR9663;STTM;病毒誘導(dǎo)的基因沉默(VIGS)技術(shù)
miRNA是一類由18~24 nt核酸構(gòu)成的非編碼小分子RNA,通過(guò)切割降解靶基因或抑制靶基因的翻譯起到調(diào)控作用[1]。miRNA參與植物各種發(fā)育進(jìn)程,如種子萌發(fā)[2]、根發(fā)育[3]、葉的發(fā)育與極性[4]、以及花器官的發(fā)育[5]。目前,已從小麥多個(gè)組織中鑒定出大量miRNA,如豫麥18花后5、15、25、30 d籽粒的miRNA[6],根、成熟葉片、小穗的組織特異性表達(dá)的miRNA[7],葉片中參與非生物脅迫的miRNA[8]。通過(guò)對(duì)小麥品種小偃6號(hào)的幼苗、旗葉和花后5、10、20 d籽粒中的miRNA進(jìn)行高通量深度測(cè)序和分析,鑒定出109個(gè)植物間保守的已知miRNA(分屬于49個(gè)miRNA家族)和113個(gè)小麥中新發(fā)現(xiàn)的miRNA。 tae-miR9663是其中新發(fā)現(xiàn)的一個(gè)miRNA,其在幼苗、旗葉和發(fā)育的籽粒中都高表達(dá),尤其在旗葉和籽粒中表達(dá)量最高[9-10],但 tae-miR9663在小麥旗葉和籽粒生長(zhǎng)發(fā)育過(guò)程中的生物學(xué)功能尚不清楚。
病毒誘導(dǎo)的基因沉默(virus induced gene silencing, VIGS)是指攜帶目的基因的重組病毒載體侵染植物后,可誘導(dǎo)植物內(nèi)源基因發(fā)生沉默而引起表型變化,從而有助于研究目的基因的功能[11]。研究表明,大麥條紋花葉病毒(barley stripe mosaic virus, BSMV)可作為VIGS的有效載體,它的基因組包含α、β、γ三個(gè)部分,其中經(jīng)修飾的γ分子是最常用的載體[12]。與傳統(tǒng)的基因功能分析方法相比,VIGS可以在植物當(dāng)代對(duì)目標(biāo)基因進(jìn)行沉默,具有簡(jiǎn)單、有效、高通量等優(yōu)點(diǎn)[13],已被廣泛用于小麥基因功能分析[12, 14-16],但是利用VIGS技術(shù)進(jìn)行小麥miRNA研究的報(bào)道較少[17]。
鑒于此,本研究首先人工合成新發(fā)現(xiàn)的小麥 tae-miR9663的小串聯(lián)模擬靶標(biāo)(short tandem target mimic, STTM),然后插入BSMV-γ載體上獲得BSMV:STTM- tae-miR9663,體外轉(zhuǎn)錄后轉(zhuǎn)染小麥葉片,實(shí)施BSMV誘導(dǎo)的通過(guò)過(guò)表達(dá)STTM而沉默小麥內(nèi)源 tae-miR9663,以快速分析 tae-miR9663對(duì)小麥旗葉和籽粒生長(zhǎng)發(fā)育的作用,為小麥超高產(chǎn)分子育種奠定理論基礎(chǔ)。
1 材料與方法
1.1 材 料
本研究所用材料為春小麥品種寧春16,將發(fā)芽一周的小麥幼苗在4 ℃春化2周后,轉(zhuǎn)移至溫室培養(yǎng)。溫室條件為24 ℃(晝)/20 ℃(夜),光照16 h·d-1。
BSMV基因組由a、β、γ三條RNA組成,BSMV病毒載體α、β、γ質(zhì)粒由美國(guó)蒙塔那州立大學(xué)Huang L博士惠贈(zèng)。
1.2 目的基因片段的克隆
根據(jù)miRBase(http://www.mirbase.org/)中小麥 tae-miR9663的成熟序列和Yan等[18]的方法設(shè)計(jì) tae-miR9663 STTM序列,并由上海生工生物工程股份有限公司合成。以合成的質(zhì)粒為模板,利用引物STTM- tae-miR9663-F/STTM- tae-miR9663-R(STTM- tae-miR9663-F:5′-GTTGTG TGGAATGTATGGAGC-3′,STTM- tae-miR9663-R:5′-GCTGTAATCACACTGGCTCA-3′,均由上海英俊生物技術(shù)有限公司合成。)進(jìn)行目的片段STTM- tae-miR9663的擴(kuò)增。擴(kuò)增產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳分離后,利用膠回收試劑盒(天根)回收目的片段。
1.3 BSMV載體的構(gòu)建
Ma等[19]將Huang L.博士惠贈(zèng)的γ載體改造后,使其序列含有兩個(gè)XcmⅠ酶切位點(diǎn),被XcmⅠ(NEB)切割后使其線性化,然后通過(guò)TA克隆方法連接膠回收的STTM- tae-miR9663,轉(zhuǎn)化DH5α感受態(tài)細(xì)胞(天根),菌落PCR檢測(cè),引物為γ-F:5′-GTGATCAACTGCCAATCGTG-3′,γ-R:5′-GTTTCCAATTCAGGCATCGT-3′。測(cè)序正確的重組載體命名為γ-STTM- tae-miR9663。
1.4 BSMV轉(zhuǎn)染小麥
用MluⅠ線性化BSMV-α、γ質(zhì)粒,用SpeⅠ線性化BSMV-β質(zhì)粒,用BssHⅡ線性化γ-STTM- tae-miR9663質(zhì)粒。參照試劑盒說(shuō)明書進(jìn)行體外轉(zhuǎn)錄(NEB),按a、β、γ各10 μL轉(zhuǎn)錄物等量混合,共30 μL,命名為BSMV:00;按a、β、γ-STTM- tae-miR9663各10 μL轉(zhuǎn)錄物等量混合,共30 μL,命名為BSMV:STTM- tae-miR9663。將配好的病毒混合液置于冰上,及時(shí)接種。將要接種前再向30 μL混合病毒液中加220 μL的GKPbuffer(50 mmol·L-1Gly、30 mmol·L-1磷酸氫二鉀、1%膨潤(rùn)土、1%硅藻土),制得250 μL的接種病毒混合液。轉(zhuǎn)染時(shí)用剛開封的乳膠手套,摩擦接種于寧春16小麥五葉一心期的第5片葉。病毒轉(zhuǎn)染小麥后,每天觀察、記錄轉(zhuǎn)染植株葉片和穗部的表型變化。
1.5 BSMV轉(zhuǎn)染小麥后的qRT-PCR分析
采用莖環(huán)法[20]設(shè)計(jì) tae-miR9663的反轉(zhuǎn)錄及后續(xù)qRT-PCR分析所用引物(表1)。利用RNAiso Plus(TAKARA)提取小麥葉片總RNA,經(jīng)DNA酶消化后,通過(guò)cDNA第一鏈合成試劑盒(康為世紀(jì))合成cDNA。以看家基因GADPH為內(nèi)參,采用BioRad-IQ5型實(shí)時(shí)熒光定量PCR儀進(jìn)行擴(kuò)增。經(jīng)過(guò)3次生物學(xué)重復(fù)和3次技術(shù)重復(fù),得到 tae-miR9663的Ct值,然后對(duì)數(shù)據(jù)進(jìn)行2-ΔΔCt相對(duì)定量分析,檢測(cè)目的基因的表達(dá)水平。

表1 反轉(zhuǎn)錄和qRT-PCR分析所用引物
2 結(jié)果與分析
2.1 重組BSMV:STTM- tae-miR9663載體的構(gòu)建
以上海生工生物工程股份有限公司合成的含有 tae-miR9663 STTM序列的質(zhì)粒為模板,采用引物STTM- tae-miR9663-F/STTM- tae-miR9663-R進(jìn)行 PCR擴(kuò)增,得到長(zhǎng)度為96 bp的STTM- tae-miR9663序列,經(jīng)純化、回收后連接到γ載體的TA克隆位點(diǎn)(TACS)上,轉(zhuǎn)化DH5α(圖1)。用引物γ-F和γ-R進(jìn)行菌落PCR,鑒定重組轉(zhuǎn)化子。測(cè)序證實(shí)獲得正確的重組載體γ-STTM- tae-miR9663。

A:BSMV由a、β、γ三條鏈組成,用長(zhǎng)方框表示其開放閱讀框,標(biāo)尺為300 bp;TACS為連接插入片段的TA克隆位點(diǎn)。B:STTM包含2個(gè)重復(fù)的模擬靶標(biāo)和一段48 nt的間隔序列。
A:Genomic organization of three BSMV components, including α, β and γ, and open reading frames are indicated by boxes with scale of 300 bp; TACS is TA cloning site designed for direct cloning of inserts. B:STTM structure contains two tandem target mimics separated by a 48 nt stem-loop linker.
圖1 BSMV基因組結(jié)構(gòu)及插入片段STTM- tae-miR9663的示意圖
Fig.1 Schematic organization of the BSMV genomes and STTM- tae-miR9663
2.2 重組BSMV:STTM- tae-miR9663轉(zhuǎn)染后小麥葉片表型及 tae-miR9663表達(dá)豐度的變化
待寧春16小麥生長(zhǎng)至五葉一心期時(shí),第五片葉分別接種空病毒BSMV:00和含有外源目的片段的重組病毒BSMV:STTM- tae-miR9663,各接種9株。接種BSMV:STTM- tae-miR9663的9個(gè)植株分別編號(hào)為9663-1、9663-2、9663-3、9663-4、9663-5、9663-6、9663-7、9663-8和9663-9。接種20 d后,將接種BSMV:STTM- tae-miR9663的9個(gè)植株與接種BSMV:00的植株對(duì)比,產(chǎn)生四種不同的表型,即第6片葉有白點(diǎn)或白條紋,第6片葉邊緣有鋸齒狀,旗葉(即第7片葉)有白點(diǎn)或白條紋,旗葉葉尖處有皺縮(圖2、表2)。同時(shí),取接種小麥植株的旗葉,以GADPH基因作為內(nèi)參,通過(guò)實(shí)時(shí)定量PCR檢測(cè) tae-miR9663的表達(dá)豐度,發(fā)現(xiàn)與對(duì)照小麥植株相比, tae-miR9663的表達(dá)豐度均下降(表2)。
2.3 重組BSMV:STTM- tae-miR9663轉(zhuǎn)染后小麥籽粒的變化
與BSMV:00相比,BSMV:STTM- tae-miR9663轉(zhuǎn)染后的小麥成熟籽粒變小(圖3),籽粒的平均長(zhǎng)度、寬度極顯著降低,平均粒重顯著降低(表3、圖4)。
A:第6片葉有白點(diǎn)或白條紋;B:第6片葉邊緣有鋸齒狀;C:旗葉有白點(diǎn)或白條紋;D:旗葉葉尖處有皺縮。
A:White dots or white stripes on the sixth leaf; B:Serrate at the sixth leaf margins; C:White dots or white stripes on flag leaf; D:Shrinking on flag leaf blade tips.

圖2 BSMV:STTM- tae-miR9663轉(zhuǎn)染后小麥葉片出現(xiàn)的四種表型
“+”表示該植株出現(xiàn)此表型;“-”表示該植株未出現(xiàn)此表型。
“+”represents the phenotypes occurred on the plant; “-”represents that the phenotypes did not occur on the plant.

圖3 BSMV:00和BSMV:STTM- tae-miR9663轉(zhuǎn)染后的小麥成熟籽粒

植株P(guān)lant長(zhǎng)度Length/mm寬度Width/mm粒重Grainweight/mgBSMV:006.13±0.09A3.37±0.13A41.80±3.22aBSMV:STTM?tae?miR96635.90±0.17B3.16±0.07B39.10±2.92b
表中數(shù)據(jù)為從3個(gè)重復(fù)中的3個(gè)植株中隨機(jī)挑選的10粒種子的平均值;數(shù)據(jù)后不同大、小寫字母表示差異在0.01和0.05水平上顯著。
The data in the table was obtained by measuring ten seeds randomly from three plants in triplicates;Different capital or low-case letters following values indicated significant difference at 0.01 and 0.05 levels,respectively.
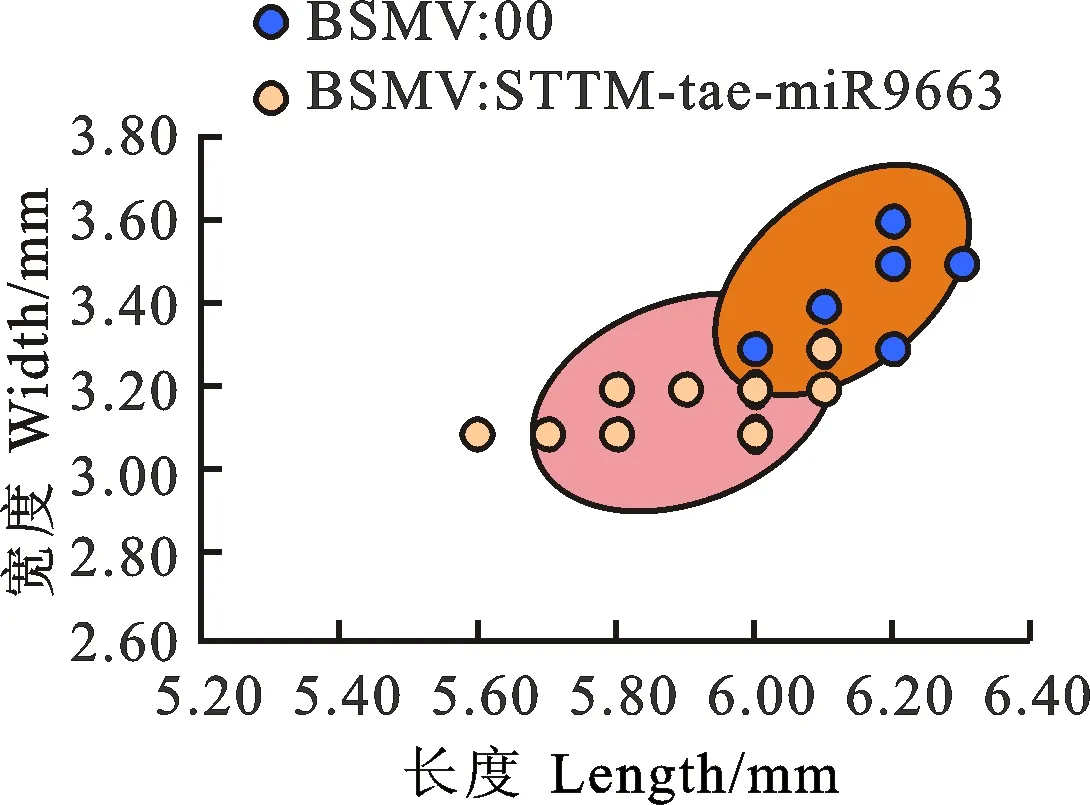
圖中的點(diǎn)代表從3個(gè)重復(fù)中的3個(gè)植株中隨機(jī)挑選的成熟籽粒。
The dots in the figure represent mature seeds chosen randomly from three plants in triplicates.
圖4 比較BSMV:00和BSMV:STTM- tae-miR9663轉(zhuǎn)染后的小麥成熟籽粒的長(zhǎng)和寬
Fig.4 Comparison of length and width of mature seeds from wheat inoculated by BSMV:00 and BSMV:STTM- tae-miR9663
3 討 論
miRNA調(diào)控植物生長(zhǎng)、發(fā)育,參與生物或非生物脅迫[21-23]。通過(guò)高通量測(cè)序已鑒定出大量的小麥miRNA[10, 24-25],其中新發(fā)現(xiàn)的一部分miRNA的表達(dá)量很高,如 tae-miR9663在小麥幼苗、旗葉和發(fā)育的籽粒中的表達(dá)量分別為1 634、5 562和10 441 RPM[10],但它們的功能尚不清楚。高表達(dá)量表明它們可能在小麥生長(zhǎng)發(fā)育過(guò)程中起到重要作用,因此鑒定它們的功能就顯得非常重要。然而穩(wěn)定的轉(zhuǎn)基因小麥的獲得費(fèi)力且效率低,而BSMV-VIGS技術(shù)具有操作簡(jiǎn)單、周期短、不需要遺傳轉(zhuǎn)化等優(yōu)點(diǎn),被廣泛應(yīng)用于小麥基因功能的快速分析[26-27],為后期轉(zhuǎn)基因小麥的篩選奠定基礎(chǔ)。
近年來(lái),STTM的應(yīng)用為植物miRNA功能研究提供了一種高效的沉默機(jī)制[17],但利用VIGS技術(shù),通過(guò)過(guò)表達(dá)STTM而有效沉默植物內(nèi)源miRNA的報(bào)道較少,而在煙草、棉花和小麥中用病毒載體分別將miR171c、miR165/166和小麥內(nèi)源miRNA的STTM序列過(guò)表達(dá)后均獲得良好的沉默效果[28-29, 17]。本研究通過(guò)構(gòu)建BSMV:STTM- tae-miR9663,利用VIGS技術(shù)轉(zhuǎn)染小麥五葉一心期的第5片葉,轉(zhuǎn)染20 d后取旗葉進(jìn)行實(shí)時(shí)定量PCR檢測(cè),結(jié)果發(fā)現(xiàn),與BSMV:00相比,STTM- tae-miR9663過(guò)表達(dá)植株 tae-miR9663的表達(dá)量下降45%~97%。這表明利用STTM沉默小麥miRNA的方法很有效,可被推廣應(yīng)用。STTM序列包含兩個(gè)TM和一段間隔序列。研究表明,48 nt間隔序列的STTM效果較好且應(yīng)用較廣泛[17-18, 29-31]。而張 力等利用間隔序列為31 nt的STTM也有效地阻止煙草microRNA171c對(duì)其靶基因的切割而沉默內(nèi)源miRNA[28]。本研究利用VIGS技術(shù),通過(guò)過(guò)表達(dá)48 nt間隔序列的STTM有效沉默小麥內(nèi)源 tae-miR9663。除STTM外,還可使用MIMICS和SPONGES等方法獲得miRNA缺失功能的植物。研究表明,MIMICS和STTM的沉默效率會(huì)變化。不同的miRNA家族對(duì)這3種不同的方法的反應(yīng)不同,沒(méi)有一個(gè)方法對(duì)所有的檢測(cè)的miRNA工作得都好[32]。因此用STTM沉默植物內(nèi)源miRNA的效率不理想時(shí),可采取嘗試其他兩種方法。
tae-miR9663在小麥旗葉中高表達(dá)[10],而且通過(guò)過(guò)表達(dá)其STTM沉默后引起葉片出現(xiàn)白點(diǎn)或白條紋、葉邊緣鋸齒狀和葉尖處皺縮的表型。miRNA是通過(guò)抑制其靶基因的表達(dá)來(lái)行使其生物學(xué)功能的。我們的降解組數(shù)據(jù)里沒(méi)有 tae-miR9663的靶基因的相關(guān)信息(數(shù)據(jù)未發(fā)表)。利用patmatch軟件預(yù)測(cè) tae-miR9663的靶基因?yàn)镸CTP2(Multiple C2 and transmembrane domain-containing protein 2)。MCTP是在進(jìn)化上保守的Ca2+依賴型C2結(jié)構(gòu)域蛋白的一個(gè)新家族,由一個(gè)可變的N端序列、三個(gè)C2結(jié)構(gòu)域、兩個(gè)跨膜域和一個(gè)短的C端序列組成。C2結(jié)構(gòu)域蛋白通過(guò)兩個(gè)緊密間隔的跨膜域被錨定在膜上,是僅次于EF-手型結(jié)構(gòu)的第二大類Ca2+感受器[33]。然而該蛋白在植物的生長(zhǎng)發(fā)育過(guò)程中的生物學(xué)功能尚未報(bào)道。葉片出現(xiàn)白點(diǎn)或白條紋可能是接種病毒后出現(xiàn)的比對(duì)照更明顯的病斑,也可能是過(guò)表達(dá) tae-miR9663的STTM后,Ca2+結(jié)合蛋白鈣調(diào)素(calmodulin, CaM)增多,而CaM與光合作用有關(guān)[34],可能抑制了光合色素的產(chǎn)生。葉片出現(xiàn)鋸齒狀和皺縮的表型可能是由于靶基因增多使得植物為了防御刺激而產(chǎn)生的結(jié)構(gòu)。通過(guò)比較BSMV:00和BSMV:STTM- tae-miR9663接種植株的成熟種子,我們發(fā)現(xiàn)BSMV:STTM- tae-miR9663接種植株種子的長(zhǎng)和寬均減小。從接種BSMV開始30 d內(nèi)病毒發(fā)揮作用,本研究接種BSMV 25 d植株開始孕穗,35 d開始抽穗,因此BSMV:STTM- tae-miR9663接種植株的種子變小可能是由于上述原因影響了旗葉發(fā)育,而旗葉為籽粒提供營(yíng)養(yǎng),進(jìn)而影響了籽粒發(fā)育,也可能是 tae-miR9663 STTM的過(guò)表達(dá)直接影響籽粒發(fā)育。綜上所述, tae-miR9663對(duì)小麥旗葉和籽粒發(fā)育起重要調(diào)控作用,它的發(fā)現(xiàn)具有重要意義,但還有許多尚未解決的問(wèn)題,如 tae-miR9663 STTM過(guò)表達(dá)的轉(zhuǎn)基因小麥的表型和 tae-miR9663靶基因的生物學(xué)功能的鑒定等,值得進(jìn)一步深入研究。
[1] YANG X J,ZHAO Y M,XIE D Y,etal.Identification and functional analysis of microRNAs involved in the anther development in cotton genic male sterile line Yu98-8A [J].InternationalJournalofMolecularSciences,2016,17(10):1677.
[2] WANG L W,LIU H H,LI D T,etal.Identification and characterization of maize microRNAs involved in the very early stage of seed germination [J].BMCGenomics,2011,12:154.
[3] HEWEZI T,MAIER T R,NETTLETON D,etal.TheArabidopsismicroRNA396-GRF1/GRF3 regulatory module acts as a developmental regulator in the reprogramming of root cells during cyst nematode infection [J].PlantPhysiology,2012,159(1):321.
[4] LIU Q L,YAO X Z,PI L M,etal.The ARGONAUTE10 gene modulates shoot apical meristem maintenance and establishment of leaf polarity by repressing miR165/166 inArabidopsis[J].ThePlantJournal,2009,58(1):27.
[5] ZHU Q H,UPADHYAYA N M,GUBLER F,etal.Over-expression of miR172 causes loss of spikelet determinacy and floral organ abnormalities in rice(Oryzasativa) [J].BMCPlantBiology,2009,9:149.
[6] MENG F R,LIU H,WANG K T,etal.Development-associated microRNAs in grains of wheat(TriticumaestivumL.) [J].BMCPlantBiology,2013,13:140.
[7] PANDEY R,JOSHI G,BHARDWAJ A R,etal.A comprehensive genome-wide study on tissue-specific and abiotic stress-specific miRNAs inTriticumaestivum[J].PLoSOne,2014,9(4):e95800.
[8] MA X L,ZIN Z Y,YANG Q H,etal.Identification and comparative analysis of differentially expressed miRNAs in leaves of two wheat(TriticumaestivumL.) genotypes during dehydration stress [J].BMCPlantBiology,2015,15:21.
[9] 閆 妍,韓 冉,趙惠賢.4條小麥保守microRNAs的表達(dá)譜分析及其靶基因預(yù)測(cè)[J].麥類作物學(xué)報(bào),2012,32(6):1041.
YAN Y,HAN R,ZHAO H X.Expression profile analysis and target gene prediction of four conserved microRNAs in wheat [J].JournalofTriticeaeCrops,2012,32(6):1041.
[10] HAN R,JIAN C,LV J Y,etal.Identification and characterization of microRNAs in the flag leaf and developing seed of wheat(TriticumaestivumL.)[J].BMCGenomics,2014,15:289.
[11] UNVER T,BUDAK H.Virus-induced gene silencing,a post transcriptional gene silencing method [J].InternationalJournalofPlantGenomics,2009,2009:198680.
[12] SCOFIELD S R,HUANG L,BRANDT A S,etal.Development of a virus-induced gene-silencing system for hexaploid wheat and its use in functional analysis of the Lr21-mediated leaf rust resistance pathway [J].PlantPhysiology,2005,138(4):2166.
[13] BENNYPAUL H S,MUTTI J S,RUSTGI S,etal.Virus-induced gene silencing(VIGS) of genes expressed in root,leaf,and meiotic tissues of wheat [J].Functional&IntegrativeGenomics,2012,12(1):144.
[14] 趙 丹,趙繼榮,黃 茜,等.利用BSMV-VIGS技術(shù)快速分析小麥 TNBL1基因的抗黃矮病功能[J].作物學(xué)報(bào),2011,37(11):2106.
ZHAO D,ZHAO J R,HUANG X,etal.Functional analysis of TNBL1 gene in wheat defense response to barley yellow dwarf virus using BSMV-VIGS technique [J].ActaAgronomicaSinica,2011,37(11):2106.
[15] CHEN K M,LI H W,CHEN Y F,etal. TaSCL14,a novel wheat(TriticumaestivumL.) GRAS gene,regulates plant growth,photosynthesis,tolerance to photooxidative stress,and senescence [J].JournalofGeneticsandGenomics,2015,42(1):21.
[16] BUHROW L M,CLARK S M,LOEWEN M C.Identification of an attenuated barley stripe mosaic virus for the virus-induced gene silencing of pathogenesis-related wheat genes [J].PlantMethods,2016,12:12.
[17] JIAO J,WANG Y C,SELVARAJ J N,etal.Barley stripe mosaic virus(BSMV) induced microRNA silencing in common wheat(TriticumaestivumL.) [J].PLoSOne,2015,10(5):e0126621.
[18] YAN J,GU Y Y,JIA X Y,etal.Effective small RNA destruction by the expression of a short tandem target mimic inArabidopsis[J].PlantCell,2012,24(2):415.
[19] MA M,YAN Y,HUANG L,etal.Virus-induced gene-silencing in wheat spikes and grains and its application in functional analysis of HMW-GS-encoding genes [J].BMCPlantBiology,2012,12:141.
[20] CHEN C F,RIDZON D A,BROOMER A J,etal.Real-time quantification of microRNAs by stem-loop RT-PCR [J].NucleicAcidsResearch,2005,33(20):e179.
[21] FENG H,WANG B,ZHANG Q,etal.Exploration of microRNAs and their targets engaging in the resistance interaction between wheat and stripe rust [J].FrontiersinPlantScience,2015,6:469.
[22] KAUR A,GUPTA O P,MEENA N L,etal.Comparative temporal expression analysis of micrornas and their target genes in contrasting wheat genotypes during osmotic stress [J].AppliedBiochemistryandBiotechnology,2017,181(2):613.
[23] DENG P C,BIAN J X,YUE H,etal.Characterization of microRNAs and their targets in wild barley(Hordeumvulgaresubsp.spontaneum)using deep sequencing [J].Genome,2016,59(5):339.
[24] CHEN F,ZHANG X F,ZHANG N,etal.Combined small RNA and degradome sequencing reveals novel MiRNAs and their targets in the high-yield mutant wheat strain Yunong 3114 [J].PLoSOne,2015,10(9):1.
[25] LIU H P,SEARLE L R,WATSON-HAIGH N S,etal.Genome-wide identification of microRNAs in leaves and the developing head of four durum genotypes during water deficit stress [J].PLoSOne,2015,10(11):1.
[26] 李建嫄,張立榮,張 娜,等.利用VIGS技術(shù)分析TcLr19中 TaSTKC8基因的抗葉銹性功能[J].河北農(nóng)業(yè)大學(xué)學(xué)報(bào),2015,38(5):12.
LI J Y,ZHANG L R,ZHANG N,etal.Functional analysis of TaSTKC8 gene on wheat leaf rust resistance in TcLr19 by virus induced gene silencing(VIGS) [J].JournalofAgriculturalUniversityofHebei,2015,38(5):12.
[27] LEE W-S,RUDD J J,KANYUKA K.Virus induced gene silencing(VIGS) for functional analysis of wheat genes involved inZymoseptoriatriticisusceptibility and resistance [J].FungalGeneticBiology,2015,79:84.
[28] 張 力,沙愛(ài)華.煙草microRNA171c的功能分析[J].植物科學(xué)學(xué)報(bào),2016,34(5):775.
ZHANG L,SHA A H.Functional analysis of microRNA171c in tobacco [J].PlantScienceJournal,2016,34(5):775.
[29] GU Z H,HUANG C J,LI F F,etal.A versatile system for functional analysis of genes and microRNAs in cotton [J].PlantBiotechnologyJournal,2014,12(5):638.
[30] SHA A H,ZHAO J P,YIN K Q,etal.Virus-based microRNA silencing in plants [J].PlantPhysiology,2014,164(1):37.
[31] TANG G L,YAN J,GU Y Y,etal.Construction of short tandem target mimic(STTM) to block the functions of plant and animal microRNAs [J].Methods,2012,58(2):124.
[32] REICHEL M,LI Y J,LI J Y,etal.Inhibiting plant microRNA activity:molecular SPONGEs,target MIMICs and STTMs all display variable effcacies against target microRNAs [J].PlantBiotechnologyJournal,2015,13(7):915.
[33] SHIN O H,HAN W P,WANG Y,etal.Evolutionarily conserved multiple C2 domain proteins with two transmembrane regions(MCTPs) and unusual Ca2+binding properties [J].TheJournalofBiologicalChemistry,2005,280(2):1641.
[34] 周 衛(wèi),汪 洪.植物鈣吸收、轉(zhuǎn)運(yùn)及代謝的生理和分子機(jī)制[J].植物學(xué)通報(bào),2007,24(6):771.
ZHOU W,WANG H.The physiological and molecular mechanisms of calcium uptake,transport,and metabolism in plants [J].ChineseBulletinofBotany,2007,24(6):771.
Silencing of tae-miR9663 in Wheat by VIGS Affects Leaf Development and Seed Size
ZHU Yanfeng1, WANG Qian1, ZHAO Huixian1, 2
(1.College of Life Science, Northwest A&F University, Yangling, Shaanxi 712100, China; 2.Northwest A&F University, State Key Laboratory of Crop Stress Biology for Arid Areas, Yangling, Shaanxi 712100, China)
MicroRNA (miRNA), which is a class of non-coding small RNA, with 18 to 24 nt in length, regulates various developmental processes in plants. tae-miR9663 is a novel miRNA highly expressing in seedling, flag leaf and seed in wheat, but its biological function is unknown. In order to explore the function of tae-miR9663, STTM (short tandem target mimic) of tae-miR9663 was synthesized, which was connected to BSMV (barley stripe mosaic virus) vector. The fifth leaves were inoculated by VIGS (virus-inducing gene silencing) in Ningchun 16. The phenotypes of leaves were observed at 20 d post-inoculation. tae-miR9663 expression in flag leaves was detected by real-time quantitative PCR and size of mature seeds was observed. Phenotypic observation results on leaves indicated that compared with BSMV:00,there are four phenotypes including dots or white stripes appeared on the sixth leaf,dots or white stripes appeared on flag leaf(the seventh leaf),serrate margins appeared in the sixth leaf,and shrinking existed on flag leaf blade tips.Real-time quantitative PCR results indicated that the expression of tae-miR9663 was decreased, showing that BSMV-VIGS could silence endogenous miRNA by overexpression of STTM effectively. Comparison of seeds from BSMV:STTM- tae-miR9663 with those from control plants, the lengths and widths of seeds from BSMV:STTM- tae-miR9663 inoculated plants were decreased.
Wheat; tae-miR9663; STTM; Virus-induced gene silencing (VIGS) technology
時(shí)間:2017-04-07
2016-12-06
2017-02-16 基金項(xiàng)目:國(guó)家自然科學(xué)基金青年基金項(xiàng)目(31501295);國(guó)家自然科學(xué)基金面上項(xiàng)目(31471482) 第一作者E-mail:xnzyf2014@163.com 通訊作者:王 倩(E-mail:ramese1021@126.com);趙惠賢(E-mail:hxzhao212@nwsuaf.edu.cn)
S512.1;S336
A
1009-1041(2017)04-0465-07
網(wǎng)絡(luò)出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170407.1020.012.html
