旁路信號通路激活介導的EML4-ALK融合基因陽性肺癌細胞株H3122對alectinib繼發耐藥的研究*
2017-05-18 12:49:29李婭妮梁柳丹陳燕瓊宋向群周韶璋
中國病理生理雜志 2017年5期
李婭妮, 梁柳丹, 陳燕瓊, 宋向群, 周韶璋△
(廣西醫科大學 1研究生院, 2附屬腫瘤醫院呼吸腫瘤內科,廣西 南寧 530021)
·論 著·
旁路信號通路激活介導的EML4-ALK融合基因陽性肺癌細胞株H3122對alectinib繼發耐藥的研究*
李婭妮1, 梁柳丹1, 陳燕瓊1, 宋向群2, 周韶璋2△
(廣西醫科大學1研究生院,2附屬腫瘤醫院呼吸腫瘤內科,廣西 南寧 530021)
目的: 本研究旨在探討肝細胞生長因子(HGF)、表皮生長因子(EGF)及轉化生長因子α(TGF-α)是否以旁路激活的方式誘導EML4-ALK融合基因陽性肺癌細胞株H3122對alectinib的耐藥,并進一步探討旁路信號激活在alectinib耐藥中的作用。方法: 用不同濃度的alectinib、克唑替尼(crizotinib)、17-DMAG或(和)HGF(50 μg/L)、EGF(100 μg/L)、TGF-α(100 μg/L)處理EML4-ALK陽性肺癌細胞株H3122,采用CCK-8法檢測細胞活力,流式細胞術檢測細胞凋亡,應用Western blot技術檢測細胞中ALK、c-Met、EGFR及相應磷酸化蛋白的表達,觀察其下游通路關鍵蛋白AKT、ERK、p-AKT和p-ERK水平。結果: Alectinib作用72 h后,H3122細胞株的活力隨著alectinib藥物濃度的增加而逐漸下降,呈劑量依賴性。HGF、EGF和TGF-α誘導后,alectinib抑制H3122細胞的生長曲線往右移,HGF、EGF和TGF-α處理能夠降低alectinib對肺癌細胞活力的抑制作用。0.05 μmol/L alectinib作用H3122細胞株48 h后的凋亡率為(20.12±1.36)%,而alectinib聯合HGF、EGF和TGF-α后的凋亡率分別為(7.85±1.03)%、(5.60±0.79)%和(4.58±1.00)%,顯著低于alectinib單藥處理(P<0.05)。Alectinib單藥成功抑制p-ALK及其下游信號通路,HGF明顯增加細胞中p-Met及其下游p-AKT、p-ERK的蛋白水平,EGF和TGF-α明顯增加細胞中p-EGFR及其下游p-AKT、p-ERK的表達,alectinib抑制p-ALK,但不能抑制HGF、EGF和TGF-α誘導的p-AKT和p-ERK的蛋白表達。此外,聯合應用crizotinib和17-DMAG可以抑制因HGF和EGFR配體而導致的H3122耐藥細胞的活力。結論: HGF、EGF和TGF-α可通過旁路激活的方式誘導EML4-ALK陽性肺癌細胞H3122對alectinib耐藥,其機制可能與HGF激活c-Met磷酸化、EGF和TGF-α激活EGFR磷酸化有關。
EML4-ALK融合基因; Alectinib; 肝細胞生長因子; 表皮生長因子受體; 耐藥性
棘皮動物微管相關蛋白樣蛋白4-間變性淋巴瘤激酶(echinoderm microtubule-associated protein-like 4-anaplastic lymphoma kinase,EML4-ALK)融合基因是非小細胞肺癌(non-small cell lung cancer,NSCLC)領域中一個重要的治療靶點。ALK包含許多重要的生物學信號通路,影響腫瘤細胞的增殖、分化與凋亡[1]。 Alectinib是一種強效、選擇性的二代ALK抑制劑,其對EML4-ALK陽性NSCLC有強的抑制作用,不僅抑制大多數與克唑替尼(crizotinib)耐藥相關的ALK突變,并且對有中樞神經系統轉移的患者有顯著療效[2],目前已被FDA批準用于克唑替尼耐藥的ALK陽性NSCLC的治療。然而最近在一些基礎實驗及ALK陽性患者中發現alectinib的耐藥,目前已知的alectinib耐藥機制中,ALK繼發耐藥突變[3]、小細胞肺癌的轉變[4-5]以及上皮間質轉化[6]等耐藥方式都有了相應的解決方案,而旁路信號激活在alectinib耐藥中的作用復雜多樣,有待進一步研究。肝細胞生長因子(hepatocyte growth factor,HGF)是原癌基因c-Met的唯一配體,表皮生長因子(epidermal growth factor,EGF)及轉化生長因子α(transforming growth factor-α,TGF-α)是表皮生長因子受體(epidermal growth factor receptor, EGFR)的配體,它們在與其相應的受體特異性結合后可激活一系列的跨膜信號通路(包括PI3K/AKT、MEK/ERK及STAT3),從而促進腫瘤的生長、侵襲和轉移,常與臨床預后差及耐藥有關[7-8]。已有研究證實HGF通過激活c-Met信號通路誘導EGFR突變患者對EGFR-TKI耐藥[9],不同于克唑替尼(為ALK及c-Met的雙靶點抑制劑),alectinib對c-Met并沒有抑制作用,c-Met信號通路可能是alectinib的耐藥機制之一。另有研究報道EGFR信號旁路與克唑替尼耐藥相關[10],但是它在alectinib耐藥中的作用目前研究有待明確。本研究擬通過外源性加入HGF、EGF及TGF-α,比較和分析其對alectinib作用下肺癌細胞的生長和凋亡的影響,觀察ALK、c-Met和EGFR信號通路的變化,旨在研究HGF、EGF及TGF-α旁路激活方式在誘導EMA4-ALK陽性NSCLC對alectinib耐藥中的作用,并探索alectinib聯合應用crizotinib和17-DMAG對H3122細胞活力的影響。
材 料 和 方 法
1 材料
1.1 細胞EML4-ALK融合基因陽性肺癌細胞株H3122購自科佰生物科技有限公司,含EML4-ALK融合基因變體1。
1.2 主要試劑與儀器 Alectinib和17-DMAG購于Selleckchem;crizotinib粉末制劑購于Cell Signaling Technology;胎牛血清和RPMI-1640培養基均購自BI;胰酶替代物購自Gibco;CCK-8細胞活力檢測試劑盒購自Dojindo;Annexin V-PE/7-AAD細胞凋亡檢測試劑盒購自BD;HGF、EGF和TGF-α購自PeproTech;抗GAPDH、ALK、p-ALK、c-Met、p-Met (Tyr1234/1235)、EGFR、p-EGFR(Tyr1068)、AKT、p-AKT(Ser473)、ERK、p-ERK(Thr202/Tyr204)單克隆抗體以及兔源和鼠源 II 抗均購自Cell Signaling Technology;Western blot實驗儀器設備購自Bio-Rad。
2 方法
2.1 細胞培養及藥物配制 將H3122細胞使用含10%滅活新生牛血清的RPMI-1640培養液,5% CO2、37 ℃的條件下培養,每3~4 d傳代1次。Alectinib、crizotinib和17-DMAG原料用DMSO溶解制成106nmol/L的母液儲存于-80 ℃冰箱,用藥時用新鮮的RPMI-1640培養基稀釋,并使DMSO的終濃度小于0.1%。HGF、EGF和TGF-α用無菌雙離子水稀釋成10 mg/L、20 mg/L和20 mg/L,分裝儲存于-20 ℃冰箱。
2.2 CCK-8實驗檢測細胞活力 取生長良好的對數期H3122細胞,每100 μL含細胞數為4×103個的細胞懸液接種于96孔板。待細胞貼壁后,吸去培養基,加入不同濃度的alectinib、crizotinib、17-DMAG或(和)50 μg/L HGF、100 μg/L EGF、100 μg/L TGF-α,以不加藥孔為空白對照孔,以不含細胞,只含對應濃度藥物培養基的孔為空白調零孔,設置4個復孔。作用72 h后,吸去培養基,加入100 μL含1/10體積的CCK-8溶液的培養基,繼續孵育2~3 h后,用酶標儀測量波長450 nm處吸光度(A)值。細胞存活率(%)=(處理組平均A值-空白調零組平均A值)/(空白對照組平均A值-空白調零組平均A值)×100%。
2.3 流式細胞術凋亡檢測 取生長良好的對數期H3122細胞,以每孔4×105個細胞數接種于6孔板中,待細胞貼壁后棄原培養基,實驗分為4組:空白對照(control)組、alectinib處理組、重組細胞因子(HGF/EGF/TGF-α)處理組和alectinib聯合重組細胞因子(HGF/EGF/TGF-α)處理組,按分組加入含0.05 μmol/L的alectinib或(和)50 μg/L HGF、100 μg/L EGF、100 μg/L TGF-α的培養基作用48 h。胰酶消化并分別收集各孔全部細胞,離心、棄上清液,冷生理鹽水洗滌2次,按照細胞凋亡試劑盒說明書對細胞進行染色,避光、室溫反應15 min后加入緩沖液,用流式細胞儀檢測細胞凋亡。實驗重復3次。
2.4 Western blot實驗 實驗分為4組:空白對照(control)組、alectinib處理組、重組細胞因子(HGF/EGF/TGF-α)處理組和alectinib聯合重組細胞因子(HGF/EGF/TGF-α)處理組,先以0.05 μmol/L alectinib處理2 h,繼而加入50 μg/L HGF、100 μg/L EGF或100 μg/L TGF-α刺激15 min,收集長滿瓶的經不同處理的H3122細胞,按蛋白提取試劑說明書提取細胞總蛋白,所得總蛋白經10%聚丙烯酰胺凝膠電泳分離后,轉移至PVDF膜,5%脫脂奶粉室溫封閉1 h。分別使用稀釋度為1∶1 000的ALK、1∶1 000的p-ALK、1∶1 000的c-Met、1∶1 000的p-Met(Tyr1234/1235)、1∶1 000的EGFR、1∶1 000的p-EGFR(Tyr1068)、1∶2 000的AKT 、1∶2 000的p-AKT(Ser473)、1∶1 000的ERK和1∶1 000的p-ERK(Thr202/Tyr204)抗體,4 ℃孵育過夜,TBST洗膜10 min,3次后用稀釋度1∶2 000的兔鼠II抗室溫下孵育1~2 h,ECL發光試劑盒顯色,計算機掃描蛋白質條帶,以條帶灰度值確定蛋白表達,實驗重復3次。
3 統計學處理
應用SPSS 17.0統計學軟件,數值以均數±標準差(mean±SD)表示,兩組間比較采用t檢驗, 多組數據比較采用單因素方差分析,以P<0.05表示差異有統計學意義。
結 果
1 CCK-8法檢測alectinib單獨或聯合HGF/EGF/TGF-α處理對H3122細胞活力的影響
Alectinib作用72 h后,H3122細胞的活力隨著alectinib藥物濃度的升高相應下降,呈現顯著的濃度依賴性抑制,alectinib作用H3122細胞72 h的IC50值為0.042 μmol/L。50 μg/L HGF處理后,alectinib作用于H3122細胞的IC50顯著升高(P<0.05),而100 μg/L EGF或100 μg/L TGF-α處理后,H3122細胞對alectinib不敏感,給予100倍于IC50值的藥物濃度仍未求出IC50,估計值遠大于3 μmol/L。HGF(50 μg/L)、EGF(100 μg/L)和TGF-α(100 μg/L)誘導H3122細胞的藥物濃度-細胞存活率曲線與非誘導曲線相比明顯往右側移。重組細胞生長因子單獨處理H3122細胞出現不同程度促進細胞活力的作用,尤其是TGF-α對H3122細胞的促生長作用顯著(P<0.05),見圖1。
Figure 1.H3122 cell viability after treatment with various concentrations of alectinib alone or combined with HGF/EGF/TGF-α for 72 h detected by CCK-8 assay. Mean±SD.n=3.*P<0.05vsalectinib group.
圖1 CCK-8法檢測不同濃度alectinib單獨或聯合HGF/EGF/TGF-α作用72 h后H3122細胞的存活率
2 流式細胞術檢測細胞凋亡
根據CCK-8法檢測H3122細胞生長的情況,選擇0.05 μmol/L alectinib單獨或聯合HGF/EGF/TGF-α處理H3122細胞48 h,檢測細胞凋亡率。結果0.05 μmol/L alectinib單藥處理48 h后H3122細胞的凋亡率為(20.12±1.36)%,50 μg/L HGF、100 μg/L EGF和100 μg/L TGF-α聯合0.05 μmol/L alectinib處理48 h后的凋亡率分別為(7.85±1.03)%、(5.60±0.79)%和(4.58±1.00)%,顯著低于alectinib單藥處理(P<0.05)。實驗結果表明alectinib對H3122細胞有促凋亡作用,HGF、EGF和TGF-α誘導作用可減少alectinib導致的H3122細胞凋亡,見圖2。
3 Western blot檢測alectinib單獨或聯合HGF/EGF/TGF-α處理對H3122細胞ALK、HGF/Met和EGFR信號通路蛋白表達的影響
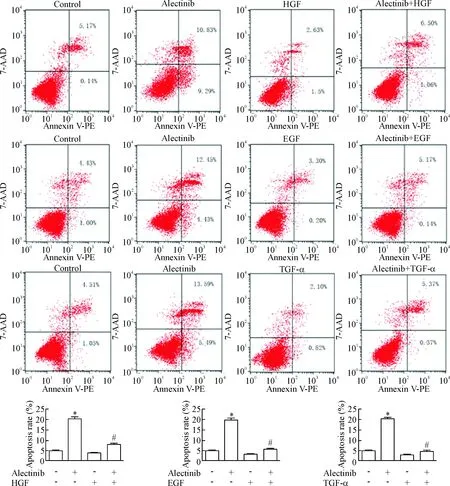
Figure 2.The apoptosis of H3122 cells treated with alectinib at 0.05 μmol/L alone or combined with HGF, EGF or TGF-α for 48 h. Mean±SD.n=3.*P<0.05vscontrol group;#P<0.05vsalectinib group.
圖2 Alectinib單獨或聯合HGF、EGF和TGF-α作用 48 h對H3122細胞凋亡的情況
為了探索HGF、EGF和TGF-α旁路激活在誘導H3122細胞對alectinib耐藥中的作用,我們采用Western blot技術檢測ALK、c-Met、EGFR及相應磷酸化蛋白的表達,同時觀察其下游通路關鍵蛋白AKT、ERK、p-AKT和p-ERK的水平。結果顯示未經處理的H3122細胞可測得ALK和下游信號通路AKT、ERK及其磷酸化蛋白,H3122也表達c-Met及EGFR蛋白,但p-Met及p-EGFR蛋白測不出。0.05 μmol/L alectinib單藥作用2 h后成功抑制ALK及其下游信號蛋白AKT、ERK的磷酸化,但對總蛋白的水平無明顯影響。當聯合重組細胞因子(50 μg/L HGF、100 μg/L EGF或100 μg/L TGF-α)同時作用于細胞時,alectinib雖然仍有效抑制ALK的磷酸化,但不能抑制下游信號蛋白AKT、ERK的磷酸化。HGF激活c-Met旁路途徑、EGF和TGF-α激活EGFR旁路途徑,可以不依賴ALK的活化而激活下游AKT、ERK的磷酸化。這些結果提示HGF通過激活c-Met旁路途徑、EGF和TGF-α通過激活EGFR旁路途徑進而激活下游通路來介導H3122細胞對alectinib的繼發耐藥,見圖3。
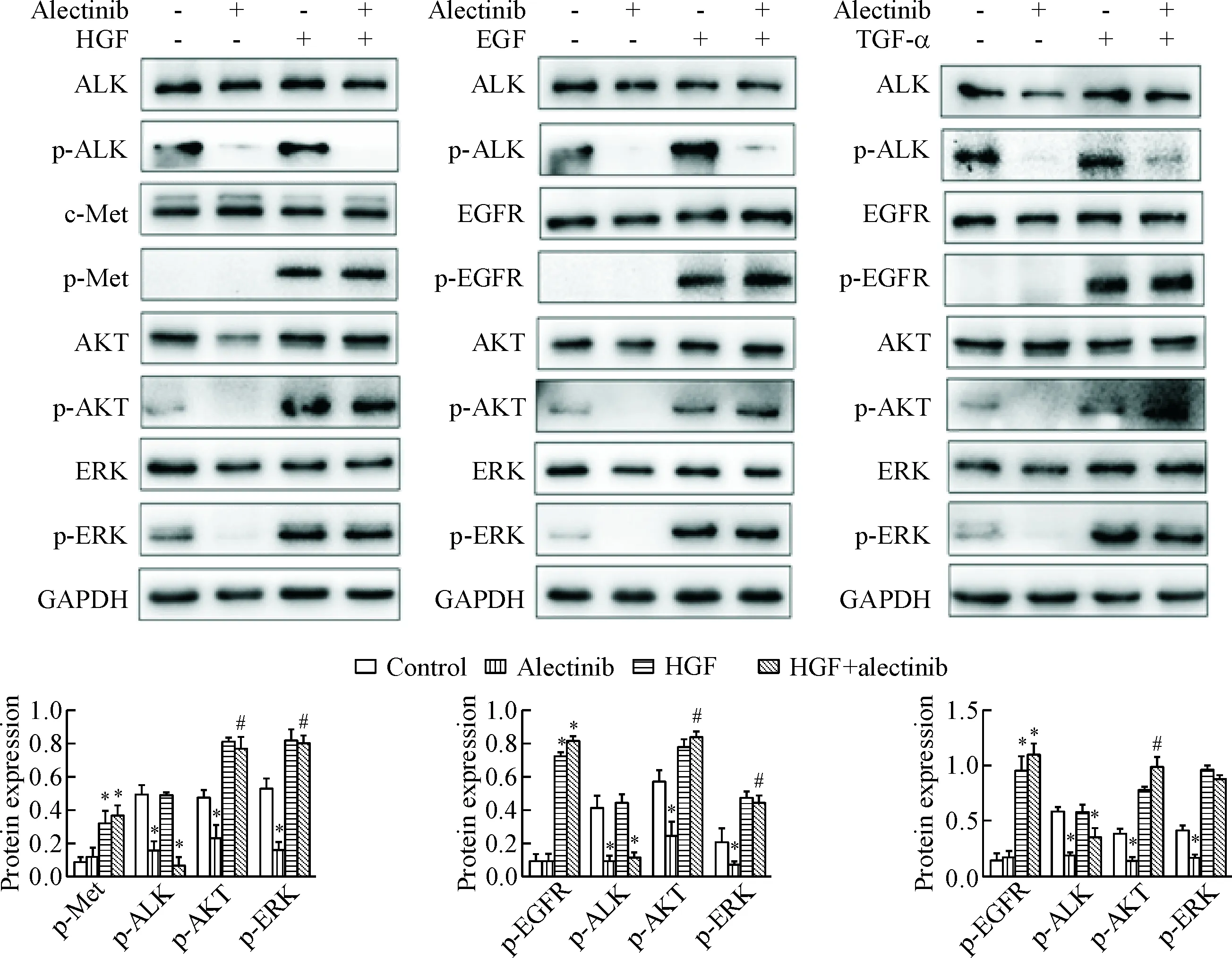
Figure 3.The protein levels in the H3122 cells treated with alectinib at 0.05 μmol/L alone or combined with HGF, EGF or TGF-α. Mean±SD.n=3.*P<0.05vscontrol group;#P<0.05vsalectinib group.
圖3 Alectinib單獨或聯合HGF、EGF和TGF-α處理后H3122細胞的蛋白水平比較
4 Alectinib聯合HGF/c-Met及熱休克蛋白 90 (heat shock protein 90, Hsp90)抑制劑對HGF/EGF/TGF-α誘導的細胞增殖的影響
為進一步研究逆轉由HGF/EGF/TGF-α介導的alectinib耐藥,我們檢測了alectinib聯合crizotinib(HGF/c-Met抑制劑)和17-DMAG(Hsp90抑制劑)對H3122細胞活力的影響。結果發現0.1 μmol/L的crizotinib可解除HGF誘發的細胞對alectinib的耐藥性變化,0.3 μmol/L 的17-DMAG 可以有效抑制H3122細胞在alectinib聯合EGF/TGF-α處理下細胞的活力,見圖4。
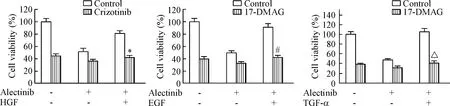
Figure 4.The effects of alectinib combined with crizotinib or 17-DMAG in the presence of HGF, EGF or TGF-α. Mean±SD.n=3.*P<0.05vsalectinib+HGF group;#P<0.05vsalectinib+EGF;△P<0.05vsalectinib+TGF-α group.
圖4 Alectinib聯合crizotinib或17-DMAG對HGF、EGF和TGF-α誘導的H3122細胞活力的影響
討 論
Alectinib是第2代的ALK抑制劑,藥物活性及選擇性都高于第1代的克唑替尼,它對克唑替尼耐藥的ALK陽性NSCLC及有中樞神經系統轉移的患者均有效。但同其它靶向藥物一樣,在治療一段時間后仍不可避免地發生耐藥,探索其耐藥的發生機制對于尋求克服方法具有重要意義。
ALK結合域二次突變[3]、細胞類型轉變[4-5]以及上皮-間質轉化[6]目前被認為是alectinib獲得性耐藥的重要機制。腫瘤細胞除了表達ALK外,同時還表達其它酪氨酸激酶活性的跨膜受體,稱之為ALK旁路酪氨酸激酶信號。在肺癌患者中,MET擴增[11]、表皮生長因子受體2(human epidermal growth factor receptor-2,HER2)擴增[12]、HGF/c-Met[9]、生長停滯特異性基因產物6(growth arrest-specific gene 6,GAS6) /AXL[13]與胰島素樣生長因子1受體(human insulin-like growth factor 1 receptor,IGF-1R)[14]信號通路的激活,均可導致EGFR突變非小細胞肺癌患者對EGFR-TKI 的耐藥。此外還發現HGF/c-Met[15]及IGF-1R[16]信號通路的激活參與了BRAF突變黑色素瘤患者對BRAF的耐藥。有研究證實EGFR[10]、KIT/SCF[17]旁路信號通路的激活與克唑替尼耐藥相關,但旁路信號激活在alectinib耐藥發生發展中的作用目前尚未完全明確。
在我們的實驗中,CCK-8細胞活力檢測顯示alectinib抑制EML4-ALK融合基因陽性肺癌細胞株H3122細胞的活力呈濃度依賴性,HGF、EGF及TGF-α誘導后IC50值顯著升高。HGF、EGF及TGF-α誘導后藥物濃度-細胞存活率曲線與非誘導曲線相比明顯往右側移。Alectinib促進細胞凋亡,HGF、EGF及TGF-α誘導后明顯降低其促進細胞凋亡的作用。從IC50的變化、藥物濃度-存活率曲線、細胞凋亡率的結果看出,HGF、EGF及TGF-α誘導H3122細胞對alectinib耐藥。本研究檢測出HGF誘導后增加細胞中p-Met及其下游信號通道蛋白,EGF、TGF-α誘導增加細胞中p-EGFR及其下游信號蛋白的水平,alectinib抑制p-ALK,但不能抑制由HGF、EGF和TGF-α誘導后p-AKT和p-ERK的蛋白水平。推測HGF激活c-Met磷酸化及EGF、TGF-α激活EGFR磷酸化,以旁路信號通路激活的方式誘導EML4-ALK陽性肺癌細胞株H3122對alectinib的耐藥。既往有研究在藥物濃度梯度遞增法誘導的alectinib耐藥株中檢測出HGF、NRG1、EGF、IGF及TGF-α的mRNA高表達[18-20],提示HGF、NRG1,EGF、IGF及TGF-α可能是alectinib耐藥的驅動因子,而本研究通過外源性加入生長因子的方法亦成功誘導H3122細胞對alectinib耐藥,進一步證實了旁路激活在耐藥中的作用。此外,NRG1/HER3的激活及IGF-1R也與alectinib耐藥相關[18, 20],可見旁路信號激活誘導alectinib耐藥的機制是復雜多樣的,還需不斷探索發現新的機制。
HGF、EGF及TGF-α雖然是本研究中介入的外源性生長因子,但在腫瘤細胞生長的微環境中也存在HGF、EGFR及IGF-1R等的配體,如內皮細胞產生的EGFR配體激活EGFR誘導克唑替尼及TAE684耐藥,纖維細胞產生的HGF激活MET信號通路誘導TAE684的耐藥[10]。有研究[20]報道了1例經alectinib治療后繼發耐藥的患者,采用IHC檢測患者用藥前及耐藥后的腫瘤活檢標本,結果提示耐藥后的標本中HGF呈現高表達,這些結果為我們的實驗結果提供了臨床依據。
有意思的是,我們的研究發現HGF/c-Met和EGFR信號通路的活化能夠介導H3122細胞對alectinib的耐藥,而克唑替尼作為ALK和c-Met的雙靶點抑制劑可抑制因HGF誘發的耐藥細胞的生長。EGFR是熱休克蛋白90(HSP90)顧客蛋白之一,Hsp90是一種分子伴侶,它在調節許多顧客蛋白構象與功能的維持和調控方面發揮重要作用[21]。17-DMAG作為HSP90的抑制劑,對多種 NSCLC 細胞株具有抗增殖效應[22]。本研究中觀察到alectinib聯合應用17-DMAG可抑制由EGF、TGF-α誘導的H3122耐藥細胞的增殖。這揭示了在ALK-TKI耐藥患者中聯合使用一代的ALK-TKI或其它TKI可能是潛在的克服耐藥的方法,另有相關的研究也報道了L1198F突變導致的三代ALK-TKI Lorlatinib耐藥患者再次用回克唑替尼并從中獲益[23],這似乎在演繹著ALK-TKI的輪回,如果在多線進展的腫瘤細胞中發現新的耐藥機制,聯合使用相關信號通路抑制劑可能仍敏感有效。我們研究alectinib的耐藥機制,為聯合應用靶向受體配體抑制劑克服alectinib耐藥提供實驗室依據,為臨床耐藥患者提供了新的治療策略。
本研究證實外源性加入HGF、EGF和TGF-α可以誘導EML4-ALK陽性細胞H3122對alectinib的耐藥,并揭示了HGF/c-Met和EGFR旁路信號通路的活化可能是其耐藥機制,這為今后深入研究針對EML4-ALK靶點的靶向耐藥和逆轉耐藥機制提供分子生物學基礎。然而,該研究僅限在細胞水平,仍需更多的臨床耐藥患者組織標本證實,可望更好闡明 HGF/c-Met和EGFR旁路信號通路激活在alectinib耐藥中的作用。
[1] 戴 輝, 宋向群, 潘星辰, 等. mTOR 信號通路在克唑替尼誘導的EML4-ALK融合基因陽性肺癌細胞株 H2228凋亡中的作用[J]. 中國病理生理雜志, 2014, 30(6):1103-1109.
[2] Gadgeel SM, Gandhi L, Riely GJ, et al. Safety and acti-vity of alectinib against systemic disease and brain metastases in patients with crizotinib-resistant ALK-rearranged non-small-cell lung cancer (AF-002JG): results from the dose-finding portion of a phase 1/2 study[J]. Lancet Oncol, 2014, 15(10):1119-1128.
[3] Katayama R, Friboulet L, Koike S, et al. Two novel ALK mutations mediate acquired resistance to the next-generation ALK inhibitor alectinib[J]. Clin Cancer Res, 2014, 20(22):5686-5696.
[4] Takegawa N, Hayashi H, Iizuka N, et al. Transformation of ALK rearrangement-positive adenocarcinoma to small-cell lung cancer in association with acquired resistance to alectinib[J]. Ann Oncol, 2016, 27(5):953-955.
[5] Fujita S, Masago K, Katakami N, et al. Transformation to SCLC after treatment with the ALK inhibitor alectinib[J]. J Thorac Oncol, 2016, 11(6):e67-e72.
[6] Kogita A, Togashi Y, Hayashi H, et al. Hypoxia induces resistance to ALK inhibitors in the H3122 non-small cell lung cancer cell line with an ALK rearrangement via epithelial-mesenchymal transition[J]. Int J Oncol, 2014, 45(4):1430-1436.
[7] Corso S, Giordano S. Cell-autonomous and non-cell-autonomous mechanisms of HGF/MET-driven resistance to targeted therapies: from basic research to a clinical perspective[J]. Cancer Discov, 2013, 3(9):978-992.
[8] Sadiq AA, Salgia R. MET as a possible target for non-small-cell lung cancer[J]. J Clin Oncol, 2013, 31(8):1089-1096.
[9] Yano S, Wang W, Li Q, et al. Hepatocyte growth factor induces gefitinib resistance of lung adenocarcinoma with epidermal growth factor receptor-activating mutations[J]. Cancer Res, 2008, 68(22):9479-9487.
[10]Yamada T, Takeuchi S, Nakade J, et al. Paracrine receptor activation by microenvironment triggers bypass survival signals and ALK inhibitor resistance in EML4-ALK lung cancer cells[J]. Clin Cancer Res, 2012, 18(13):3592-3602.
[11]Engelman JA, Zejnullahu K, Mitsudomi T, et al. MET amplification leads to gefitinib resistance in lung cancer by activating ERBB3 signaling[J]. Science, 2007, 316(5827):1039-1043.
[12]Takezawa K, Pirazzoli V, Arcila ME, et al. HER2 amplification: a potential mechanism of acquired resistance to EGFR inhibition in EGFR-mutant lung cancers that lack the second-site EGFRT790M mutation[J]. Cancer Discov, 2012, 2(10):922-933.
[13]Zhang Z, Lee JC, Lin L, et al. Activation of the AXL kinase causes resistance to EGFR-targeted therapy in lung cancer[J]. Nat Genet, 2012, 44(8):8528-8560.
[14]Yeo CD, Park KH, Park CK, et al. Expression of insulin-like growth factor 1 receptor (IGF-1R) predicts poor responses to epidermal growth factor receptor (EGFR) tyrosine kinase inhibitors in non-small cell lung cancer patients harboring activating EGFR mutations[J]. Lung Cancer, 2015, 87(3):311-317.
[15]Straussman R, Morikawa T, Shee K, et al. Tumour micro-environment elicits innate resistance to RAF inhibitors through HGF secretion[J]. Nature, 2012, 487(7408):500-504.
[16]Hilmi C, Larribere L, Giuliano S, et al. IGF1 promotes resistance to apoptosis in melanoma cells through an increased expression of BCL2, BCL-X(L), and survivin[J]. J Invest Dermatol, 2008, 128(6):1499-1505.
[17]Katayama R, Shaw AT, Khan TM, et al. Mechanisms of acquired crizotinib resistance in ALK-rearranged lung Cancers[J]. Sci Transl Med, 2012, 4(120):120ra17.
[18]Dong X, Fernandez-Salas E, Li E, et al. Elucidation of resistance mechanisms to second-generation ALK inhibitors alectinib and ceritinib in non-small cell lung cancer cells[J]. Neoplasia, 2016, 18(3):162-171.
[19]Tani T, Yasuda H, Hamamoto J, et al. Activation of EGFR bypass signaling by TGFα overexpression induces acquired resistance to alectinib in ALK-translocated lung cancer cells[J]. Mol Cancer Ther, 2016,15(1):162-171.
[20]Isozaki H, Ichihara E, Takigawa N, et al. Non-small cell lung cancer cells acquire resistance to the ALK inhibitor alectinib by activating alternative receptor tyrosine kinases[J]. Cancer Res, 2016,76(6):1506-1516.
[21]Taipale M, Jarosz DF, Lindquist S. HSP90 at the hub of protein homeostasis: emerging mechanistic insights[J]. Nat Rev Mol Cell Biol, 2010, 11(7):515-528.
[22]Kobayashi N, Toyooka S, Soh J, et al. The anti-proliferative effect of heat shock protein 90 inhibitor, 17-DMAG, on non-small-cell lung cancers being resistant to EGFR tyrosine kinase inhibitor[J]. Lung Cancer, 2012, 75(2):161-166.
[23]Shaw AT, Friboulet L, Leshchiner I, et al. Resensitization to crizotinib by the lorlatinib ALK resistance mutation L1198F[J]. N Engl J Med, 2016, 374(1):54-61.
(責任編輯: 陳妙玲, 羅 森)
Bypass signaling pathway activation mediates resistance ofEML4-ALKfusion gene positive lung cancer cell line H3122 to alectinib
LI Ya-ni1, LIANG Liu-dan1, CHEN Yan-qiong1, SONG Xiang-qun2, ZHOU Shao-zhang2
(1PostgraduateCollege,2DepartmentofRespiratoryOncology,AffiliatedTumorHospital,GuangxiMedicalUniversity,Nanning530021,China.E-mail:zhoushaozhang@qq.com)
AIM: To detect the changes of active status of bypass signaling pathways inEML4-ALKpositive lung cancer cell line H3122 treated with alectinib, hepatocyte growth factor (HGF), epidermal growth factor (EGF) and transforming growth factor-α (TGF-α), and to explore the potential mechanisms. METHODS:EML4-ALKpositive cell line H3122 was treated with increasing concentrations of alectinib or/and induced by HGF, EGF and TGF-α. The cell viability was measured by CCK-8 assay. The cell apoptosis was analyzed by flow cytometry. The protein levels and phosphorylation status of ALK, c-Met and EGFR, and the downstream molecules AKT, ERK, p-AKT and p-ERK were examined by Western blot. RESULTS: The viability of the H3122 cells was inhibited by alectinib in a dose-dependent manner after administrated for 72 h, and the IC50value was 0.042 μmol/L. The concentration-growth curves of the H3122 cells shifted to the right after induced by HGF, EGF and TGF-α. After treatment with alectinib at 0.05 μmol/L for 48 h, the apoptotic rate of H3122 cells was (20.12±1.36)%, while the apoptotic rates of the cells in the groups of alectinib combined with HGF, EGF or TGF-α were (7.85±1.03)%, (5.60±0.79)% and (4.58±1.00)%, respectively. Those values were remarkably lower than those in alectinib single treatment group (P<0.05). Alectinib inhibited the protein levels of p-ALK and its downstream signaling pathway molecules, while HGF significantly up-regulated the protein levels of p-Met and its downstream p-AKT and p-ERK. Besides, EGF and TGF-α remarkablely up-regulated the protein levels of p-EGFR and its downstream p-AKT and p-ERK. Combined treatment with crizotinib and 17-DMAG successfully inhibited the viability of the H3122 cells even in the presence of the HGF and EGFR ligands, respectively. CONCLUSION: Bypass signaling pathways are activated by HGF, EGF and TGF-α inEML4-ALKpositive lung cancer cell line H3122, which may be linked to alectinib resistance.
EML4-ALKfusion gene; Alectinib; Hepatocyte growth factor; Epidermal growth factor receptor; Drug resistance
1000- 4718(2017)05- 0769- 07
2016- 11- 28
2017- 03- 17
國家自然科學基金資助項目(No. 81260357; No. 81060188); 廣西自然科學基金資助項目(No. 2015GXNSFAA139162)
R730.23
A
10.3969/j.issn.1000- 4718.2017.05.001
雜志網址: http://www.cjpp.net
△通訊作者 Tel: 0771-5334955; E-mail: zhoushaozhang@qq.com
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
保健醫苑(2022年5期)2022-06-10 07:46:38
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國當代醫藥(2015年17期)2015-03-01 02:03:58
