GDF11在棕櫚酸誘導骨骼肌細胞胰島素抵抗中的作用研究
2017-06-22 14:50:17敬媛媛
中國藥理學通報 2017年6期
敬媛媛,吳 凡,李 蓉
(重慶醫科大學附屬第一醫院內分泌科,重慶 400016)
GDF11在棕櫚酸誘導骨骼肌細胞胰島素抵抗中的作用研究
敬媛媛,吳 凡,李 蓉
(重慶醫科大學附屬第一醫院內分泌科,重慶 400016)
GDF11;骨骼肌細胞;胰島素抵抗;PGC-1α;棕櫚酸;葡萄糖攝取
胰島素抵抗增加2型糖尿病、心血管疾病和高血壓的發病風險[1-2]。胰島素抵抗的特征為胰島素作用的敏感性或反應性降低,主要表現在骨骼肌、肝臟和脂肪組織。骨骼肌胰島素抵抗與線粒體數目減少或功能紊亂密切相關。既往研究發現肥胖伴胰島素抵抗人群骨骼肌內線粒體數目減少[3]。骨骼肌線粒體減少或功能紊亂可降低其氧化脂肪酸的能力,致肌細胞內脂質沉積最終導致胰島素抵抗[4-5]。
GDF11(growth differential factor 11)是新近發現的一種具有逆轉衰老作用的分泌性蛋白,近年來備受關注。研究發現血循環中GDF11隨年齡的增長而下降,并且GDF11有明顯改善骨骼肌代謝,逆轉心肌肥厚,改善大腦認知等功能[6-9],顯示出GDF11廣泛的生物學活性和潛在的應用價值。Sinha等[6]發現GDF11可改善衰老骨骼肌的線粒體的形態結構,增強線粒體功能,同時增加老年小鼠骨骼肌中PGC-1α的表達。PGC-1α為PPARs的轉錄共激活因子,參與機體線粒體生物合成[10]。研究發現過表達小鼠骨骼肌PGC-1α可改善糖代謝[11]。基于GDF11在改善衰老骨骼肌功能方面的重要作用,本研究首次從體外實驗探索GDF11是否可改善骨骼肌細胞胰島素抵抗。
1 材料與方法
1.1 材料 C2C12小鼠成肌細胞購自中國科學院上海細胞庫;棕櫚酸(PA)購自美國Sigma公司;GDF11購自美國R&D公司;2NBDG購自美國Invitrogen公司;CCK-8試劑盒購自美國Biotool公司;實時定量逆轉錄及擴增試劑盒購自日本TaKaRa公司;兔抗鼠PGC-1α一抗購自美國Abcam公司,羊抗兔二抗購自德國Millipore公司。
1.2 方法
1.2.1 細胞培養及實驗分組 C2C12小鼠成肌細胞用高糖DMEM培養基(含10%胎牛血清、1%青霉素及鏈霉素)于37℃,飽和濕度,5% CO2培養箱中培養。細胞匯合達到80%~100%時,換用誘導培養基(含2%馬血清,1%青霉素及鏈霉素)培養,每天更換培養基。經過6 d誘導分化,形成多核肌管細胞用于后續實驗研究。將實驗分為以下6組:對照組;GDF11(50 μg·L-1)干預組; GDF11(100 μg·L-1)干預組;棕櫚酸(0.5 mmol·L-1)干預組;GDF11(50 μg·L-1)聯合棕櫚酸(0.5 mmol·L-1)干預組;GDF11(100 μg·L-1)聯合棕櫚酸(0.5 mmol·L-1)干預組。
1.2.2 CCK-8 法檢測細胞活力 取對數生長期細胞,以5×106·L-1細胞懸液按每孔100 μL接種于96孔板。培養24 h細胞貼壁后,吸棄培養液,分別加入含不同濃度的GDF11(0、50、100 μg·L-1)的培養基培養24 h。吸棄培養液,每孔加入100 μL含CCK-8的無血清培養基(CCK-8 ∶培養基=1 ∶10)培養30 min。在酶標儀上以450 nm波長測吸光度值。
1.2.3 2NBDG糖攝取的檢測 細胞接種于6孔板,誘導分化6 d后形成肌管。細胞經干預后加入100 nmol·L-1胰島素作用30 min。用預冷的HBSS緩沖液洗3遍后,加入含有100 μmol·L-12NBDG的HBSS緩沖液培養30 min。分別提取6孔板每孔細胞的總蛋白,操作過程需避光。將提取的總蛋白分別轉移至96孔板中,在熒光酶標儀上以激發波長/發射波長(Ex/Em,485/535 nm)測吸光度值。每組吸光度值以相應蛋白濃度進行校正。
1.2.4 實時熒光定量PCR檢測mRNA的表達 采用TRIzol法提取細胞總RNA,反轉錄為cDNA,進行實時熒光定量PCR擴增反應。具體操作步驟按照說明書進行,引物設計見Tab 1。根據擴增曲線測得內參基因及目的基因的Ct值,目的基因的表達用GAPDH進行標化,采用2-△△ct法分析。

Tab 1 Sequences of primers for RT-PCR

Fig 1 C2C12 myoblasts successfully differentiate into ±s,n=3)
A: C2C12 myoblasts(×100); B: C2C12 myotubes(×100);C:The mRNA level of desmin after differentiation;D:The mRNA level of myogenin after differentiation.*P<0.05,**P<0.01vsundifferentiation group
1.2.5 Western blot檢測蛋白的表達 提取細胞總蛋白,BCA法測定蛋白濃度。蛋白經SDS-PAGE電泳分離后轉至PVDF膜;5%脫脂奶粉封閉2 h后,加入兔抗鼠PGC-1α一抗4℃孵育過夜,TBST 緩沖液洗3次后,加入羊抗兔二抗室溫孵育1 h;TBST充分洗膜3次,ECL試劑顯影及成像。用蛋白條帶吸光度與相應內參吸光度比值進行分析。
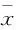
2 結果
2.1 C2C12成肌細胞分化鑒定 C2C12成肌細胞經2%馬血清完全培養基培養6 d后分化為長梭形且含有多個細胞核的肌管細胞。分化細胞較未分化細胞肌管標志基因desmin及myogenin的表達明顯升高(P<0.05,P<0.01)。提示誘導分化條件適當。見Fig 1。
2.2 GDF11對骨骼肌細胞葡萄糖攝取的影響 如Fig 2A所示,不同濃度GDF11(50、100 μg·L-1)干預對細胞活性無明顯影響,所以后續實驗采用這兩個濃度。2NBDG糖攝取結果顯示,與對照組相比,棕櫚酸干預組胰島素介導的葡萄糖攝取明顯降低(P<0.01),提示胰島素抵抗模型建立成功。而GDF11(50、100 μg·L-1)干預組與對照組相比葡萄糖攝取無明顯變化。與棕櫚酸干預組比較,GDF11 (50、100 μg·L-1)聯合棕櫚酸干預組葡萄糖攝取無明顯變化。棕櫚酸干預組GLUT4及IRS-1的表達較對照組明顯降低(P<0.01)。而聯合GDF11干預后與單純棕櫚酸組比較GLUT4及IRS-1表達無明顯變化。見Fig 2。
2.3 GDF11對PGC-1α表達的影響 與對照組比較,棕櫚酸干預組PGC-1α mRNA及蛋白的表達均降低(P<0.01),而GDF11(50、100 μg·L-1)干預對PGC-1α mRNA及蛋白的表達無明顯影響。與棕櫚酸干預組比較,GDF11(50、100 μg·L-1)聯合棕櫚酸干預組PGC-1α mRNA及蛋白的表達無明顯變化。見Fig 3。
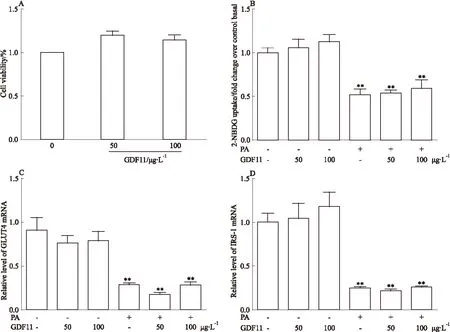
Fig 2 Effect of GDF11 on insulin resistance induced by palmitate in C2C12 ±s,n=3)
A:Cell viability was measured after different concentrations of GDF11 intervention; B:Glucose uptake in each group; C:The mRNA level of GLUT4 in each group; D:The mRNA level of IRS-1 in each group.**P<0.01vsControl group
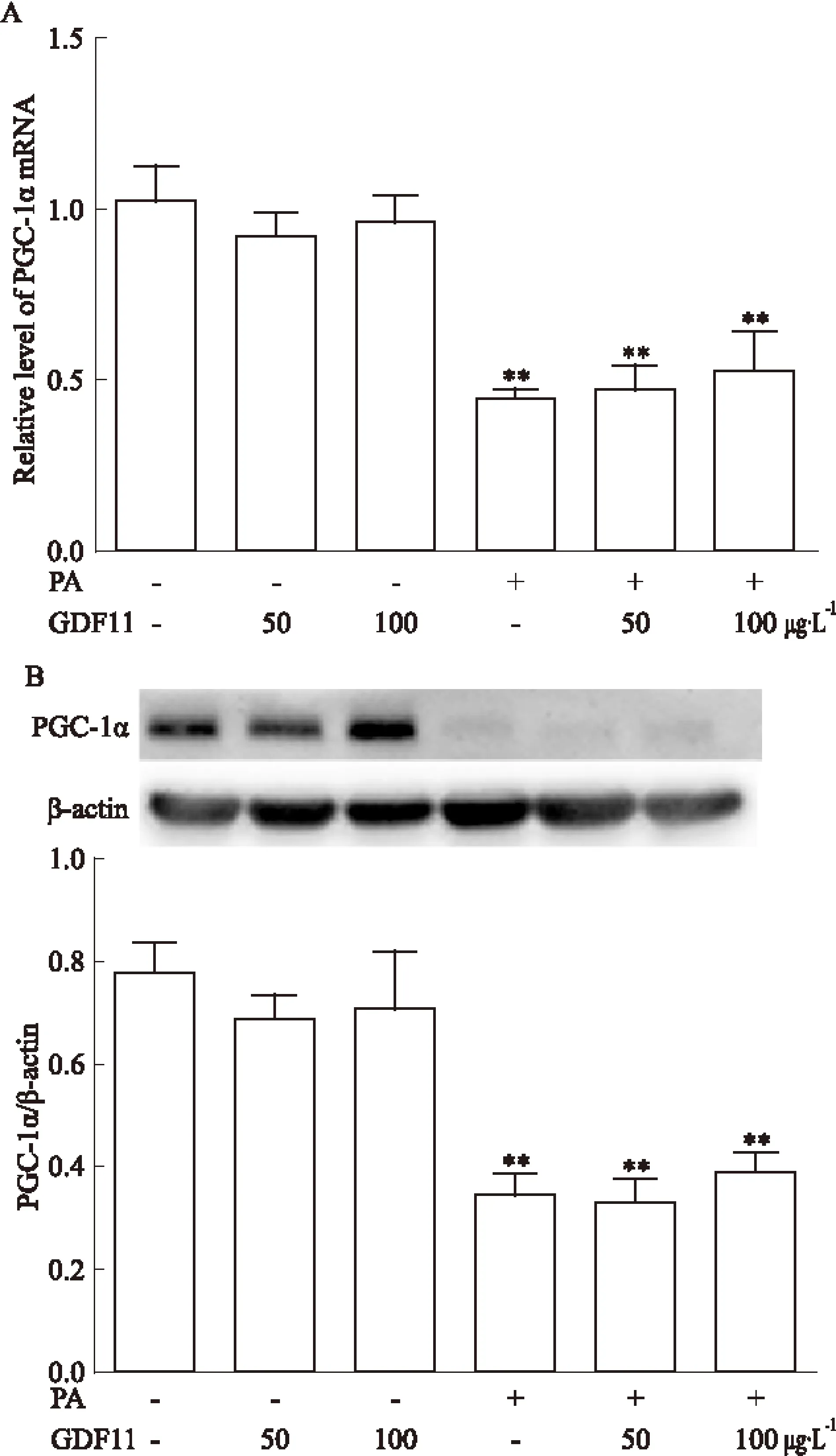
Fig 3 Effect of GDF11 on expression of PGC-1α in
A:The mRNA level of PGC-1α in each group;B:The protein level of PGC-1α in each group.**P<0.01vsControl group
3 討論
骨骼肌是胰島素刺激后攝取葡萄糖的主要部位,占全身葡萄糖攝取的80%以上[12]。既往研究發現棕櫚酸可誘導胰島素抵抗[13],本實驗采用棕櫚酸干預C2C12肌管細胞,使胰島素介導的葡萄糖攝取明顯減少,成功構建骨骼肌細胞胰島素抵抗模型。過多的游離脂肪酸進入肌細胞,線粒體氧化脂肪酸能力降低,致骨骼肌內脂質沉積[4]。骨骼肌內脂質沉積與骨骼肌胰島素抵抗密切相關[14]。本研究發現棕櫚酸同時可降低PGC-1α的表達,這與Coll等[15]的研究結果一致。PGC-1α主要通過調節線粒體的數量和功能來調節細胞能量代謝[16,17]。特異性過表達骨骼肌中PGC-1α不僅可增加線粒體生物合成,還可增強胰島素敏感性,促進葡萄糖的吸收[11]。而在胰島素抵抗患者的骨骼肌中PGC-1α的表達明顯下降[3]。
GDF11是一種分泌型生長分化因子,屬于TGF-β超家族一員,在骨骼肌、胰腺、腎臟、心臟等組織廣泛表達[18]。近年GDF11逆轉衰老的作用備受矚目,Loffredo等[7]報道恢復老年小鼠血液中GDF11到年輕小鼠的水平可逆轉年齡相關的心肌肥厚。Sinha等[6]發現血液中GDF11隨著小鼠年齡增長而降低,補充GDF11可改善衰老骨骼肌結構和功能,促進PGC-1α的表達,增加線粒體生物合成。本研究旨在探索GDF11是否可改善骨骼肌胰島素抵抗。結果發現GDF11未明顯改善棕櫚酸誘導骨骼肌細胞胰島素抵抗。GDF11對棕櫚酸誘導后下調的PGC-1α表達也沒有明顯改善作用,這與Sinha等[6]研究顯示GDF11可促進PGC-1α表達的結果不一致。其原因可能是模型不一樣,Sinha等的研究模型是老年小鼠模型,體內是一個相對較復雜的整體環境,而本研究是體外構建的骨骼肌細胞胰島素抵抗模型,環境較單一。目前的研究對GDF11逆轉衰老的作用存在爭議。與Sinha的研究結果相反,Egerman等[19]發現大鼠血液和骨骼肌中的GDF11蛋白隨著年齡增長而升高,并且補充GDF11抑制老齡化骨骼肌的再生能力和骨骼肌細胞的分化;Egerman等提出Sinha對GDF11蛋白的檢測方法難以將GDF11蛋白與Myostatin蛋白區分開來,而后者具有抑制肌肉生成的作用。Smith等[20]研究發現GDF11對老年小鼠心臟結構和功能沒有改善作用,也沒有減少心肌細胞肥大,而是誘導心肌細胞肥大。
綜上所述,本研究在體外成功構建了骨骼肌胰島素抵抗模型,棕櫚酸干預使骨骼肌細胞葡萄糖攝取減少及GLUT4和IRS-1的表達下降,而加用GDF11干預后,骨骼肌細胞葡萄糖攝取及GLUT4和IRS-1的表達無明顯改善。本研究首次探討GDF11對骨骼肌細胞胰島素抵抗的影響。可為目前GDF11的作用提供參考。
(致謝:感謝重慶醫科大學附屬第一醫院脂糖代謝實驗室提供的實驗條件和技術支持。)
[1] Cooper S A, Whaley-Connell A, Habibi J, et al. Renin-angiotensin-aldosterone system and oxidative stress in cardiovascular insulin resistance[J].AmJPhysiolHeartCircPhysiol,2007,293(4):H2009-23.
[2] Reaven G M, Chen Y D. Insulin resistance, its consequences, and coronary heart disease. Must we choose one culprit[J]?Circulation,1996,93(10):1780-3.
[3] Patti M E, Butte A J, Crunkhorn S, et al. Coordinated reduction of genes of oxidative metabolism in humans with insulin resistance and diabetes: Potential role of PGC1 and NRF1[J].ProcNatlAcadSciUSA,2003,100(14):8466-71.
[4] Lowell B B, Shulman G I. Mitochondrial dysfunction and type 2 diabetes[J].Science,2005,307(5708):384-7.
[5] Kelley D E, He J, Menshikova E V, et al. Dysfunction of mitochondria in human skeletal muscle in type 2 diabetes[J].Diabetes,2002,51(10):2944-50.
[6] Sinha M, Jang Y C, Oh J, et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle[J].Science,2014,344(6184):649-52.
[7] Loffredo F S,Steinhauser M L,Jay S M, et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy[J].Cell,2013,153(4):828-39.
[8] Olson K A, Beatty A L, Heidecker B, et al. Association of growth differentiation factor 11/8, putative anti-ageing factor, with cardiovascular outcomes and overall mortality in humans: analysis of the Heart and Soul and HUNT3 cohorts[J].EurHeartJ, 2015,36(48):3426-34.
[9] Katsimpardi L, Litterman N K, Schein P A, et al. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors[J].Science,2014,344(6184):630-4.
[10]Wu Z, Puigserver P, Andersson U, et al. Mechanisms controlling nitochondrial biogenesis and respiration through the Thermogenic coactivator PGC-1[J].Cell, 1999,98(1):115-24.
[11]Summermatter S, Shui G, Maag D, et al. PGC-1alpha improves glucose homeostasis in skeletal muscle in an activity-dependent manner[J].Diabetes, 2013,62(1):85-95.
[12]Abdul-Ghani M A, DeFronzo R A. Pathogenesis of insulin resistance in skeletal muscle[J].JBiomedBiotechnol, 2010,2010:476279.
[13]吳文君,湯孫寅炎,時俊鋒,等. 二甲雙胍抑制SREBP-1c改善高脂誘導的骨骼肌胰島素抵抗[J].中國藥理學通報,2016,32(1):55-9.
[13]Wu W J, Tang S Y Y, Shi J F, et al. Metformin ameliorates PA-induced skeletal muscle insulin resistance by suppressing SREBP-1c[J].ChinPharmacolBull,2016,32(1):55-9.
[14]Krssak M, Falk Petersen K, Dresner A, et al. Intramyocellular lipid concentrations are correlated with insulin sensitivity in humans: a 1H NMR spectroscopy study[J].Diabetologia, 1999,42(1):113-6.
[15]Coll T, Jove M, Rodriguez-Calvo R, et al. Palmitate-mediated downregulation of peroxisome proliferator-activated receptor-gamma coactivator 1alpha in skeletal muscle cells involves MEK1/2 and nuclear factor-kappaB activation[J].Diabetes, 2006,55(10):2779-87.
[16]Arany Z. PGC-1 coactivators and skeletal muscle adaptations in health and disease[J].CurrOpinGenetDev, 2008,18(5):426-34.
[17]Buler M, Aatsinki S M, Izzi V, et al.SIRT5 is under the control of PGC-1alpha and AMPK and is involved in regulation of mitochondrial energy metabolism[J].FASEBJ, 2014,28(7):3225-37.
[18]McPherron A C. Metabolic functions of myostatin and Gdf11[J].ImmunolEndocrMetabAgentsMedChem, 2010,10(4):217-31.
[19]Egerman Marc A, Cadena Samuel M, et al. GDF11 increases with age and inhibits skeletal muscle regeneration[J].CellMetabolism,2015,22(1):164-74.
[20]Smith S C,Zhang X, Zhang X, et al. GDF11 does not rescue aging-related pathological hypertrophy[J].CircRes,2015,117(11):926-32.
Role of GDF11 in skeletal muscle cell insulin resistance induced by palmitate
JING Yuan-yuan, WU Fan, LI Rong
(DeptofEndocrinology,theFirstAffiliatedHospitalofChongqingMedicalUniversity,Chongqing400016,China)
Aim To investigate the role of GDF11 in palmitate induced skeletal muscle insulin resistance.Methods The C2C12 cells were sorted into control group, GDF11 intervention group, palmitate group and GDF11 combined with palmitate group. Cell viability was measured by CCK-8, and the glucose uptake was determined by 2NBDG. The mRNA level of myotube marker genes(desmin,myogenin), insulin mediate glucose uptake related genes(GLUT-4,IRS-1) and PGC-1α were tested by RT-PCR. The protein expression of PGC-1α was detected by western blot.Results GDF11 had little effect on cell viability of skeletal muscle cells. Compared with control group, the glucose uptake and the expression of GLUT-4,IRS-1,PGC-1α were significantly decreased by palmitate intervention. Compared with palmitate group, the glucose uptake and the expression of GLUT-4,IRS-1,PGC-1α were not significantly changed by GDF11.Conclusion Palmitate can induce skeletal muscle cell insulin resistance, but GDF11 may not significantly improve the skeletal muscle cell insulin resistance.
GDF11;skeletal muscle cell;insulin resistance;PGC-1α;palmitate;glucose uptake
時間:2017-5-25 17:44 網絡出版地址:http://kns.cnki.net/kcms/detail/34.1086.R.20170525.1744.014.html
2017-01-18,
2017-03-16
國家自然科學基金資助項目(No 81200588);國家臨床重點專科建設項目(No 2011-170)
敬媛媛(1991-),女,碩士生,研究方向:內分泌及代謝,E-mail:776839843@qq.com; 李 蓉(1972-),女,博士,教授,主任醫師,碩士生導師,研究方向:內分泌及代謝,通訊作者,E-mail:rongli232006@163.com
10.3969/j.issn.1001-1978.2017.06.007
A
1001-1978(2017)06-0767-05
R322.74;R329.24; R347.8;R458.5;R977.6摘要:目的 探討GDF11對棕櫚酸誘導骨骼肌細胞胰島素抵抗的影響。方法 用棕櫚酸構建骨骼肌細胞胰島素抵抗模型,分為對照組、GDF11干預組、棕櫚酸干預組和GDF11聯合棕櫚酸干預組。CCK-8檢測細胞活力,2NBDG檢測細胞葡萄糖攝取。實時熒光定量PCR檢測肌管標志基因(desmin、myogenin),胰島素介導葡萄糖攝取相關基因(GLUT-4、IRS-1)及PGC-1α的表達。Western blot檢測PGC-1α蛋白水平的表達。結果 不同濃度GDF11對骨骼肌細胞活力無明顯影響。與對照組相比,棕櫚酸干預組葡萄糖攝取及GLUT-4、IRS-1、PGC-1α的表達明顯降低(P<0.05)。與棕櫚酸干預組相比,GDF11聯合棕櫚酸干預組葡萄糖攝取及GLUT-4、IRS-1、PGC-1α的表達無明顯變化。結論 棕櫚酸可成功誘導骨骼肌細胞胰島素抵抗,而GDF11對骨骼肌細胞胰島素抵抗沒有明顯改善作用。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
人人健康(2023年26期)2023-12-07 03:55:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年2期)2019-08-23 08:12:10
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
中國醫藥科學(2015年15期)2015-02-27 12:32:27
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國全科醫學(2013年36期)2013-01-25 06:20:58