茉莉酸甲酯對茯苓三萜生物合成的調控研究
2017-07-03 15:09:37崔培梧魯耀邦張志麗張書航黃紹國易剛強
湖南中醫(yī)藥大學學報 2017年6期
陳 林,崔培梧,魯耀邦,廖 彥,張志麗,張書航,黃紹國,易剛強*
(1.湖南中醫(yī)藥大學藥學院,湖南 長沙 412008;2.湖南中醫(yī)藥大學中藥藥性與藥效實驗室,湖南 長沙 410208)
茉莉酸甲酯對茯苓三萜生物合成的調控研究
陳 林1,2,崔培梧1,2,魯耀邦1,2,廖 彥1,2,張志麗1,2,張書航1,2,黃紹國1,2,易剛強1,2*
(1.湖南中醫(yī)藥大學藥學院,湖南 長沙 412008;2.湖南中醫(yī)藥大學中藥藥性與藥效實驗室,湖南 長沙 410208)
目的 探索茉莉酸甲酯(methyl jasmonate,MeJA)對茯苓三萜生物合成的調控效應及其機制。方法 從MeJA添加量、添加時間角度考察MeJA對茯苓三萜生物合成的調控效應,確定最佳調控策略,在此基礎上采用熒光定量PCR(Real time PCR,RT-PCR)技術分析MeJA對茯苓甲羥戊酸途徑法尼基焦磷酸合成酶基因(fps)和鯊烯合酶基因(sqs)基因表達水平的影響,揭示其調控機制。結果 MeJA可顯著促進茯苓三萜的生物合成,最佳添加策略為發(fā)酵4 d添加150 μmol/L的MeJA,此時茯苓三萜得率可達20.95 mg/L,為空白組的1.55倍、吐溫-80組的1.32倍;RT-PCR結果顯示MeJA可使fps、sqs基因表達水平顯著上調,分別為吐溫-80對照組的1.17倍、788.70倍。結論 MeJA通過顯著上調sqs基因表達實現茯苓三萜生物合成的過程強化,為一種有效的促進茯苓三萜合成的外源調控因子。
茯苓;三萜;生物合成;發(fā)酵調控;茉莉酸甲酯
茯苓[Wolfiporia cocos(Schwein.)Ryvarden&Gilb.(Basidiomycota,Polyporaceae)]俗稱茯菟,其性甘味平,入心、肺、脾、腎經,具有滲濕利水、健脾和胃、寧心安神、強精益髓之功效,為我國名貴大品種菌類中藥,始載于《五十二病方》,在《神農本草經》被列為上品[1-4]。目前超過80%的中藥方劑都配伍了茯苓,如四君子湯、茯苓半夏湯、茯苓補心湯、天王補心丹、五苓散等[5];此外以茯苓為原料配伍的中成藥也有300多種[6],這使得市場對茯苓資源的需求日益擴大。
研究表明三萜為茯苓的主要活性成分之一,具有抗癌、抗炎、降糖等多種活性[7-9],且發(fā)酵菌絲體中茯苓三萜含量明顯優(yōu)于茯苓菌核[10],這進一步證實了采用發(fā)酵法生產茯苓活性成分的可行性。在真菌中,三萜類化合物主要由甲羥戊酸途徑(mevalonate pathway,MVA)合成[11],而茉莉酸甲酯(methyl Jasmonate,MeJA)作為良好的次級代謝調節(jié)因子,可通過上調 MVA 途徑 hmgs、hmgr、mvd、fps、sqs 等關鍵酶基因表達水平而實現靈芝酸的強化合成[12],因此,擬探索MeJA對液態(tài)發(fā)酵體系茯苓三萜生物合成的過程調控效應。實驗將從MeJA濃度、添加時間方面系統考察MeJA對茯苓三萜生物合成的影響,并采用熒光定量PCR(real-time PCR,RT-PCR)技術分析MeJA干預對茯苓MVA途徑關鍵酶基因sqs(編碼鯊烯合酶)、fps(編碼法尼基焦磷酸酶)的表達影響,進而揭示MeJA對茯苓三萜合成過程的調控機制。
1 材料與方法
1.1 菌種、試劑與儀器
Poria cocos 5.0078,購于中國科學院微生物研究所菌種保藏中心,現保藏于湖南中醫(yī)藥大學中藥藥性與藥效實驗室。
熊果酸 (批號:110742-201220,質量分數≥98%,中國食品藥品檢定研究院);RNA反轉錄試劑盒 (iScriptTMcDNA Synthesis Kit, Bio-Rad Laboratories);Real-time PCR 擴增試劑盒 (iTaqTMUniversal SYBR?Green Supermix, Bio-Rad Laboratories);Trizol(15596026,Invitrogen);茉莉酸甲酯(392707,質量分數≥95%,Sigma);其余試劑如葡萄糖、蛋白胨、酵母浸膏、硫酸鎂、磷酸二氫鉀等均為國產分析純。
生化培養(yǎng)箱(ZSD-1270,上海智誠分析儀器制造有限公司);紫外-可見分光光度計(UV-1800,日本島津公司);高速冷凍離心機(GL-26M,長沙平凡儀器儀表有限公司);PCR儀(Mastercycler gradient,Eppendorf); 核酸蛋白分析儀 (BioPhotometer,Eppendorf);熒光定量 PCR 儀(CFX96 Real-time system,Bio-Rad)。
1.2 方法
1.2.1 菌種活化 取保藏的茯苓菌種,轉接至馬鈴薯葡萄糖瓊脂(potato dextrose agar,PDA)平板上,在28℃下培養(yǎng)10 d,然后轉接至不含瓊脂的馬鈴薯葡萄糖培養(yǎng)基中,裝液量為100 mL/250 mL,于28℃、180 r/min振蕩培養(yǎng)7 d,作為二級菌種。
1.2.2 基礎發(fā)酵培養(yǎng) 基礎發(fā)酵培養(yǎng)基為葡萄糖(2%)、蛋白胨(0.5%)、酵母浸膏(0.4%)、KH2PO4(0.1%)、MgSO4(0.05%)、初始 pH 5.5。 培養(yǎng)條件:250 mL 三角瓶裝液量90 mL,接種10 mL二級種子,于28℃、180 r/min振蕩培養(yǎng)19 d,培養(yǎng)結束后測定生物量和茯苓三萜含量。
1.2.3 MeJA對茯苓三萜合成過程的干預 以基礎發(fā)酵培養(yǎng)基組為空白對照,以添加0.2 mL/L吐溫-80的基礎培養(yǎng)基組為助溶劑對照,考察MeJA不同濃度(100、150、200、250、300 μmol/L)和不同添加時間(0、2、4、6、8 d) 對茯苓菌生長和茯苓三萜合成的影響,進而確定最佳的MeJA調控工藝。
1.2.4 相關分析方法 生物量 (DCW)測定:取20 mL發(fā)酵液,用干燥至恒重的濾紙抽濾,菌體經蒸餾水洗滌抽濾3次后,于60°C烘箱干燥至恒重,稱量后計算菌體干重。
茯苓三萜(PA)含量測定:取0.1 g于60°C烘箱干燥至恒重并研磨均勻的茯苓菌絲體粉末,加入6 mL異丙醇靜置提取 24 h,于 8 000×g離心10 min,所得上清液即為茯苓三萜。以熊果酸為標準品,采用香草醛-高氯酸顯色法[10]測定茯苓菌絲體中茯苓三萜的含量。
茯苓三萜生物合成途徑關鍵酶基因表達水平的分析:以單因素篩選出的MeJA最佳添加條件進行培養(yǎng),并以吐溫-80組為空白對照,濾紙過濾收集發(fā)酵菌絲體,DEPC水沖洗數次后取100 mg進行液氮研磨,參照Trizol試劑說明書提取茯苓菌總RNA,然后采用2%瓊脂糖凝膠電泳檢測樣品完整性,并采用核酸蛋白分析其純度和濃度。以所得RNA為模板,逆轉錄成cDNA,用于茯苓三萜生物合成途徑關鍵酶基因sqs和fps的實時定量PCR。逆轉錄體系:5 ×iScript reaction mix 4 μL、iScript 逆 轉 錄 酶1 μL、模板 RNA 2 μL、Rnase-free ddH2O 13 μL;逆轉錄反應條件:25℃ 5 min,42℃ 30 min,85℃ min,4℃保存。RT-PCR反應體系:2×iTaqTMUniversal SYBR? Green Supermix 5 μL、上下游引物各 1 μL(見表1)、模板 1 μL、 Rnase-free ddH2O 2 μL。RT-PCR 反應條件:95 ℃預變性3 min,95 ℃10 s,59℃ 20 s,39個循環(huán)。實驗設置3個生物學重復。 采用 2-ΔΔCt法[13]進行相對定量分析,其中 ΔΔCt=[(Ctgene-Ct18s)]實驗組-[(Ctgene-Ct18s)]對照組。
2 結果與分析
2.1 MeJA添加量對茯苓菌細胞生長和三萜生物合成的影響
MeJA是茉莉屬(Jasminum)中素馨花中香精油的重要組成成分,水溶性很差,因此若將其用于水相培養(yǎng)基必須加入助溶劑。本研究中選取吐溫-80作為MeJA的助溶劑,在發(fā)酵0 d分別考察了100、150、200、250、300 μmol/L MeJA 對茯苓菌細胞生長和茯苓三萜合成的影響,結果如圖1所示。與空白組相比,助溶劑對照組(按2 mL/L比例添加吐溫-80)的生物量顯著提高,但對胞內三萜合成的增強效應不明顯,推測應該為吐溫-80強化茯苓菌液態(tài)發(fā)酵體系溶氧傳遞效率所致[14];MeJA調控組細胞生長量增幅不再明顯,但茯苓三萜濃度顯著提高,在150 μmol/L MeJA調控條件下可獲得最大胞內三萜得率,為 20.77 mg/L,是空白組的 2.18 倍、吐溫-80組的 1.80倍;300 μmol/L MeJA 調控條件下生物量雖然可達最大值3.67 g/L,但茯苓三萜得率較低。因此,150 μmol/L MeJA為最佳的調節(jié)濃度。
2.2 MeJA添加時間對茯苓菌細胞生長和三萜生物合成的影響
三萜為茯苓菌的次級代謝產物,與茯苓細胞生長階段和生長環(huán)境都有很大關系,因此外源調控因子的添加時間也是影響其調控效應的重要因素。本研 究 分 別 考 察 了 在 發(fā) 酵 0、2、4、6、8 d 添 加150 μmol/L MeJA對茯苓三萜合成的調控效應,結果顯示(圖2)在發(fā)酵4 d添加MeJA對茯苓三萜合成的誘導效應最好,可達20.95 mg/L,分別是空白組的 1.55倍、吐溫-80組的1.32倍;而此時的茯苓菌絲生物量卻低于吐溫-80調控組,進一步說明茯苓三萜的合成與茯苓菌絲生長對營養(yǎng)條件及環(huán)境需求存在差異。

表1用于RT-PCR分析的相關引物
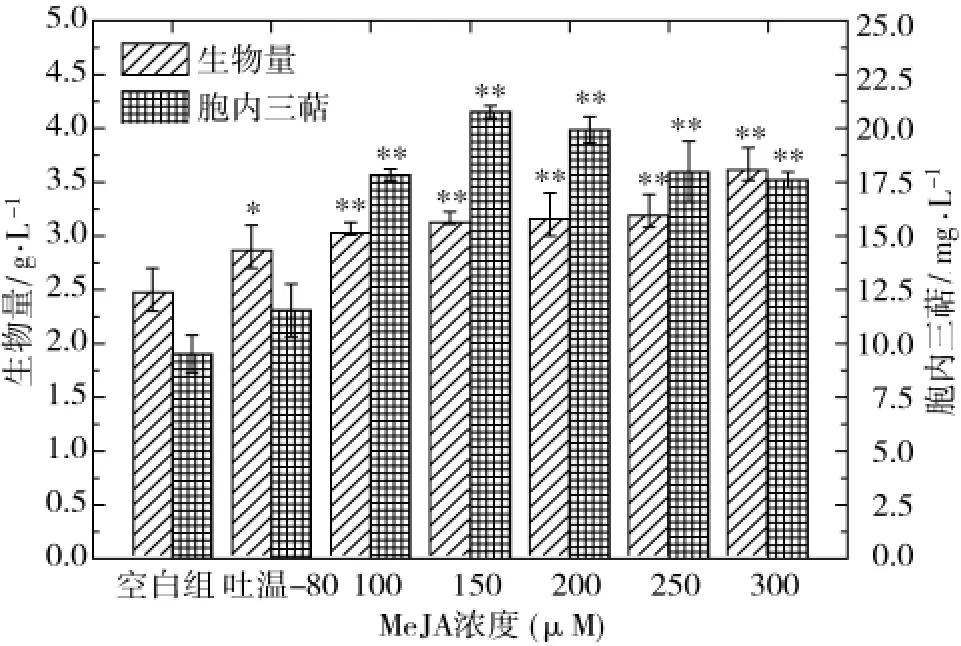
圖1 MeJA添加量對茯苓菌菌體生長和三萜合成的影響
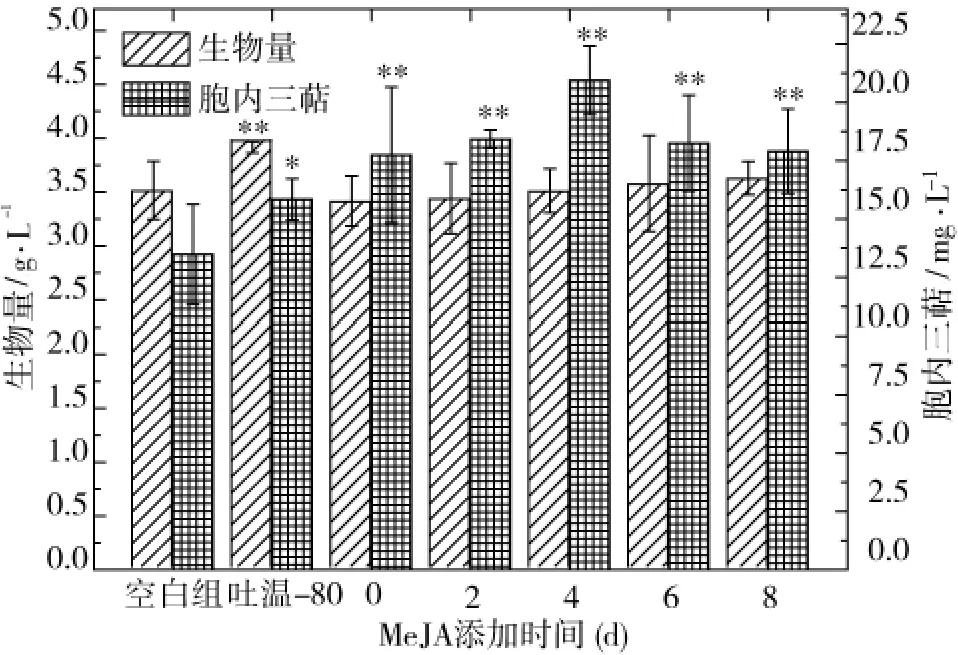
圖2 MeJA添加時間對茯苓菌菌體生長和三萜合成的影響
2.3 MeJA對茯苓菌MVA途徑關鍵酶基因表達水平的影響
研究表明,茯苓三萜由MVA途徑合成[11],MVA途徑中法尼基焦磷酸合成酶 (farnesyl diphosphate synthase,FPS)催化異戊烯基焦磷酸和二甲基丙烯焦磷酸合成法尼基焦磷酸,接著在鯊烯合酶(squalene synthase,SQS)的催化下合成鯊烯,進而轉化為茯苓三萜。由上述實驗結果可知在發(fā)酵4 d添加150 μmol/LMeJA可獲得最大的茯苓三萜積累水平,因此,為探索MeJA的調控機制,采用RT-PCR技術分析了MeJA調控組和吐溫-80組茯苓菌MVA途徑關鍵酶fps和sqs的表達水平差異。提取的RNA質量及純度、濃度分析分別見圖3、表2。電泳圖顯示5、18、28S RNA的條帶均非常清晰,且無彌散現象,同時28S RNA片段亮度是18S RNA片段的2倍,說明RNA完整性良好;RNA質量分析顯示OD260/OD280在1.8左右,意味著RNA純度良好,符合RT-PCR的要求,因此可以以此RNA為模板進行RT-PCR分析。RT-PCR分析結果如圖4所示,MeJA組的fps、sqs表達水平分別是吐溫-80組的1.17倍及788.70倍,意味著sqs在茯苓三萜生物合成途徑中發(fā)揮著非常關鍵的調控作用,因此可被認為茯苓三萜生物合成途徑的關鍵調控節(jié)點。
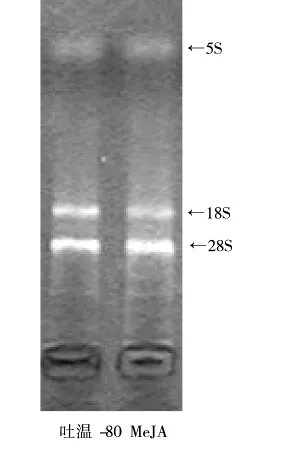
圖3茯苓菌絲體RNA電泳圖

表2茯苓菌絲體RNA的質量分析
3 結論
MeJA被認為調控次級代謝產物合成的重要外源調控因子,已被成功應用于靈芝[12]、白木香[15]、軟紫草[16]等多種植物、真菌次級代謝產物的強化合成研究,但在茯苓三萜生物合成的過程強化領域的應用還鮮見報道。本研究從MeJA添加量、添加時間角度考察了其對茯苓三萜生物合成的調控效應,結果顯示在茯苓菌發(fā)酵4 d添加150 μmol/L MeJA可使得茯苓三萜的積累水平達到20.95 mg/L,為空白組的 1.55倍、吐溫-80組的 1.32倍,取得了較好的調控效果。為揭示該調控機制,采用RT-PCR技術分析了MeJA調控條件下茯苓MVA途徑關鍵酶fps和sqs的表達水平,顯示MeJA可顯著提高sqs表達水平,為吐溫-80組的788.70倍,意味著鯊烯合酶基因的表達水平在茯苓三萜生物合成途徑發(fā)揮著關鍵調控作用,因此可作為后續(xù)理性調控的關鍵節(jié)點。這些結果將為實現茯苓三萜合成的理性強化提供必要基礎。
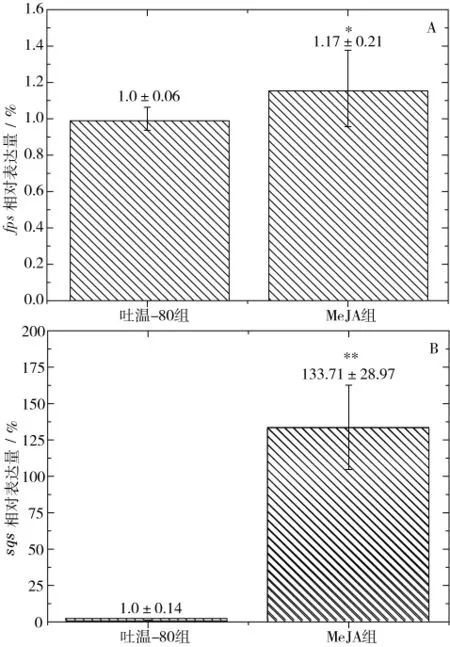
圖 4 MeJA 對茯苓菌 fps(A)、sqs(B)表達水平的影響
[1]國家藥典委員會.中華人民共和國藥典一部[S].北京:中國醫(yī)藥科技出版社,2015:241-242.
[2]張建逵,竇德強,王 冰,等.茯苓類藥材的本草考證[J].時珍國醫(yī)國藥,2014,25(5):1181-1183.
[3]劉鴻高,王元忠.云南不同產地茯苓總三萜含量測定[J].中國食用菌,2012,31(1):33-34.
[4]Wu LF,Wang KF,Mao X,et al.Screening and analysis of the potential bioactive components of Poria cocos (Schw.)Wolf by HPLC and HPLC-MS(n)with the aid of chemometrics[J].Molecules,2016,21(2):21020227.
[5]董遠文.茯苓中三萜的分離純化及含量測定研究[D].武漢:湖北中醫(yī)藥大學,2014:1.
[6]趙英博,徐 斌,昝俊峰,等.不同產地茯苓中茯苓酸含量的比較研究[J].中國中醫(yī)藥信息雜志,2009,16(7):41-42.
[7]Rios JL.Chemical constituents and pharmacological properties of Poria cocos[J].Planta Medica,2011,77(7):681-691.
[8]Wang W,Dong H,Yan R,et al.Comparative study of lanostane-type triterpene acidsin differentpartsofPoria cocos(Schw.)Wolf by UHPLC-Fourier transform MS and UHPLC-triple quadruple MS[J].Journal of Pharmaceutical and Biomedical Analysis,2015,102:203-214.
[9]Dong H,Wu P,Yan R,et al.Enrichment and separation of antitumor triterpene acids from the epidermis of Poria cocos by pH-zone-refining counter-current chromatography and conventional high-speed counter-current chromatography[J].Journal of Separation Science,2015,38(11):1977-1982.
[10]Shu SH,Chen B,Zhou MC,et al.De novo sequencing and transcriptome analysis of Wolfiporia cocos to reveal genes related to biosynthesis of triterpenoids[J].PLoS One,2013,8(8):e71350.
[11]Wang J,Li Y,Liu D.Cloning and characterization of farnesyl diphosphate synthase gene involved in triterpenoids biosynthesis from Poria cocos[J].International Journal of Molecular Sciences,2014,15(12):22188-22202.
[12]Ren A,Qin L,Shi L,et al.Methyl jasmonate induces ganoderic acid biosynthesis in the basidiomycetous fungus Ganoderma lucidum[J].Bioresource Technology,2010,101(17):6785-6790.
[13]張倡輝,郝 娟,羅 軍,等.干擾TIP47基因對奶山羊乳腺上皮細胞脂滴形成及相關基因表達的影響[J].農業(yè)生物技學報,2016,24(12):1-8.
[14]崔培梧,胡亞強,鐘瑜萍,等.氧載體對茯苓菌液態(tài)深層發(fā)酵的影響[J].中藥材,2015,38(6):1157-1160.
[15]徐艷紅,楊 欣,梁 良,等.茉莉酸甲酯誘導的白木香cDNA文庫的構建及初步鑒定[J].中草藥,2014,45(1):102-106.
[16]Hao H,Lei CY,Dong QL,et al.Effects of exogenous methyl jasmonate on the biosynthesis of shikonin derivatives in callus tissuesofArnebiaeuchroma[J].Applied Biochemistryand Biotechnology,2014,173(8):2198-2210.
(本文編輯 蘇 維)
Study on Regulation of Triterpenoids Biosynthesis in Poria cocos Submerged Fermentation System by Methyl Jasmonate
CHEN Lin1,2,CUI Peiwu1,2,LU Yaobang1,2,LIAO Yan1,2,ZHANG Zhili1,2,ZHANG Shuhang1,2,HUANG Shaoguo1,2,YI Gangqiang1,2*
(1.College of Pharmacy,Hunan University of Chinese Medicine,Changsha,Hunan 410208,China;2.TCM Potency&Efficacy Laboratory,Hunan University of Chinese Medicine,Changsha,Hunan 410208,China)
Objective To explore the regulation effects and its molecular mechanism of methyl jasmonate (MeJA)on triterpenoid biosynthesis in submerged fermentation system of Poria cocos.Methods The effects of addition amount and feeding time of MeJA on Poria cocos triterpenoid biosynthesis were discussed,respectively.Besides,to illustrate regulation mechanism of MeJA,real time PCR(RT-PCR)was employed to analysis the expression level of fps and sqs in mevalonate(MVA)pathway of Poria cocos.Results The optimal regulation strategy was adding 150 μmol/L MeJA on 4thday.Under this condition,the maximum content can reach up to 20.95 mg/L,which was 1.55 times of blank group,1.32 times of Tween-80 group.RTPCR results showed that fps and sqs gene expression level both increased significantly,which were 1.17 times,788.70 times of that in Tween-80 group,respectively.Conclusion MeJA,as an effective exogenous regulatory factor for increasing triterpenoid biosynthesis level of Poria cocos,could efficiently improve triterpenoid biosynthesis level in Poria cocos submerged fermentation system.
Poria cocos;triterpenoids;biosynthesis;regulation of fermentation;methyl jasmonate
R284.4
A
doi:10.3969/j.issn.1674-070X.2017.06.007
2016-12-29
湖南省發(fā)改委科研計劃 (湘發(fā)改高技 〔2014〕1199號);湖南省中醫(yī)藥科研計劃重點項目 (201621);湖南省教育廳科研計劃(16C1219);湖南省中藥粉體與創(chuàng)新藥物省部共建國家重點實驗室培育基地開放基金(ZYFT201404);湖南中醫(yī)藥大學研究性學習和創(chuàng)新性實驗計劃(201616);湖湘中藥資源保護與利用協同創(chuàng)新中心開放基金(00175-12);湖南中醫(yī)藥大學生物工程重點學科共同資助。
陳 林,女,在讀碩士研究生,研究方向為藥用菌物次級代謝產物的合成及其過程調控。
* 易剛強,男,副教授,碩士研究生導師,E-mail:ygq8228@163.com。
本文引用:陳 林,崔培梧,魯耀邦,廖 彥,張志麗,張書航,黃紹國,易剛強.茉莉酸甲酯對茯苓三萜生物合成的調控研究[J].湖南中醫(yī)藥大學學報,2017,37(6):606-610.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中國衛(wèi)生(2016年3期)2016-11-12 13:23:26
