無脊椎動物先天免疫分子研究概述
2017-08-20 01:59:18劉文杰
生物學教學 2017年7期
劉文杰
(福建省廈門市大同中學 361009)
章躍陵
(廣東汕頭大學理學院生物學系/廣東省海洋生物技術重點實驗室 515063)
無脊椎動物的先天防御體系是在長期自然選擇過程中進化形成的,非常有效且功能強大,其生物學意義已被廣泛認識。無脊椎動物先天免疫系統與哺乳動物的先天免疫系統有很高的同源性,其相關研究成果可以為理解更復雜的高等動物免疫反應提供線索。此外,隨著耐藥性細菌的出現和傳統抗生素治療作用的下降,亟需尋找新型抗致病菌的活性物質。因此,關于無脊椎動物的先天免疫分子的研究是一個熱點。
1 無脊椎動物先天免疫分子
一般認為,無脊椎動物缺乏真正的抗體和特異性的免疫細胞,機體防御反應依靠非特異性的先天免疫,與之相關的先天免疫分子是免疫反應系統的一個重要組成部分。
1.1 酚氧化酶 酚氧化酶(PO)是生物體內一類含銅氧化酶,能通過氧化酚或多酚形成對應的醌,大體可分為酪氨酸酶、兒茶酚氧化酶和漆酶三大類(后二者統稱為多酚氧化酶)。無脊椎動物的多酚氧化酶常以酶原的形式存在,其活性位點由一個保守的苯丙氨酸占據并阻擋底物進入,不具有酶活性,需要時經過特異絲氨酸蛋白酶的切割形成有活性的PO才能起作用。
昆蟲多酚氧化酶是一種非常重要的免疫蛋白,可同時介導體液和細胞有關成分參與免疫,是誘發黑化反應的主要酶,對于防止病原微生物入侵以及傷口愈合至關重要。依靠多酚氧化酶的免疫作用,昆蟲可以在發病前清除大部分病原物,若多酚氧化酶的活性消失或被有效抑制后,昆蟲對病原物的抵抗力顯著下降[1]。漆酶型PO是海洋無脊椎動物免疫應答系統的重要組成部分,其氧化產物通常具有窄譜抗菌活性,且對弧菌有強烈的殺傷抑制作用,但與昆蟲漆酶型PO在氧化產物抗菌譜上的差異顯著。此外,漆酶型PO自身可通過多種方式有效應答病毒、細菌和真菌的入侵,對病原的刺激非常敏感、應答迅速,在無脊椎動物免疫力調控和病害預警中顯示出潛在的應用價值[2]。
1.2 凝集素 凝集素(lectin)是無脊椎動物先天免疫系統的重要組分,存在于細胞外基質以及血淋巴細胞表面、細胞基質等處。凝集素也廣泛分布于動物、植物和微生物中。凝集素是一類對特定細胞多糖具有結合親和力的、多價構型的熱敏蛋白或糖蛋白復合物,能選擇性凝集脊椎動物血細胞、微生物細胞和單細胞藻類以及沉淀某些復雜碳水化合物。因此,可凝集外來病原菌、進行非己識別、調理和介導血細胞吞噬,在免疫應答早期階段發揮著積極的作用。
軟體動物中的凝集素在識別異體、凝集和吞噬等方面發揮著重要作用,如櫛孔扇貝(Chlamysfarreri) 的血淋巴細胞內有多種含量豐富的凝集素,可凝集大腸桿菌等,并在細胞介導的吞噬作用中起協同作用[3]。文蛤(Meretrixmeretrix)在受到細菌刺激后,C-型凝集素基因表達量顯著上升, 并參與對微生物入侵的免疫應答, 有助于提高文蛤等的免疫防御能力[4]。對幾種棘皮動物凝集素的研究發現,它們在進行調理作用和創傷修復等防御機制中起重要作用。例如,擬球海膽(Paracentrotuslividus) 紅細胞凝集素可能參與凝血作用、創面修復作用、調理作用和包囊作用,刺瓜參(Cucumariaechinata) 中C型凝集素可溶解兔和人的紅細胞,并可能對外來微生物產生毒性等[5]。
1.3 抗菌肽 抗菌肽(AMPs)是無脊椎動物天然免疫系統的主要成分之一,是高效、具有廣譜抗微生物(細菌和真菌)的活性小分子多肽,一般由12~100個氨基酸殘基組成,具有親脂、親水性(雙親性)。抗菌肽的抗菌機制與傳統抗生素截然不同,特點是分子量小、水溶性好、耐熱性強、無免疫原性、殺菌速度快和不易被蛋白酶水解等,在無脊椎動物防御病原微生物中起著重要作用,被譽為“天然抗生素”。自Boman等發現第一個抗菌肽——天蠶素以來, 人們相繼又從細菌、真菌、高等植物、昆蟲、兩棲類、哺乳動物甚至人體中發現具有類似性質的活性多肽。據統計,截止到2012年9月已確定的抗菌肽數量達到2032種,并且數據還在不斷增加中。
現有研究結果顯示,抗菌肽除了具有廣譜抗菌活性外,還具有抗病毒和寄生蟲等活性,以及趨化和免疫調節、中和毒素、抗腫瘤、促進血管生成和創傷修復等生物學功能[6]。
1.4 血藍蛋白 血藍蛋白(hemocyanin)是一種與呼吸作用有關的含銅蛋白,其特點是脫氧狀態為無色,結合氧時呈藍色,存在于節肢動物和軟體動物的血淋巴中。通常認為血藍蛋白的主要生物學功能與機體內的氧運輸有關,故與血紅蛋白、蚯蚓血紅蛋白合稱為動物界的三大呼吸蛋白。但近年來的研究表明,血藍蛋白還是無脊椎動物中的一種具有多種免疫學活性的非特異性免疫分子,可以識別和抵抗病毒、細菌、真菌、異種動物紅細胞及腫瘤細胞等多種病原體。血藍蛋白及其亞基不僅在胰蛋白酶、甲殼動物血細胞溶解產物等激活下表現出酚氧化酶免疫功能, 而且其自身或裂解產生的不同分子質量大小的片段也可以抵御病毒的入侵,具備抗菌活性[7]。例如,對蝦血藍蛋白對魚、雞、鼠和人的紅細胞以及對副溶血性弧菌、大腸桿菌K12和金黃色葡萄球菌呈現明顯的凝集活性[8]。此外,血藍蛋白還與能量的貯存、滲透壓的維持以及蛻皮過程的調節有關[9]。
1.5 補體系統 補體系統是由一系列40多種蛋白質分子所組成、有著精密調控機制的蛋白質反應系統。補體作為眾多免疫效應機制中的一員,在免疫過程的末端和吞噬作用以及炎癥反應等一起發揮著作用,是連接先天免疫和適應性免疫的樞紐[10]。
Tagawa 等研究發現中國鱟中TtCRP-1、TL-5A、TL-1 和TPL-1分子可以召集C3和B因子的復合物到革蘭氏陽性菌和酵母表面形成C3轉化酶,進而催化更多的C3分子結合到這些微生物表面,同時還發現一個未知的C因子參與其中,提示其可能存在不同于脊椎動物的補體激活途徑[11]。彭茂瀟等[12]對無脊椎動物補體系統中已報道的C1q、MBL、ficolin等的成分、功能、激活途徑和系統進化情況進行了較詳盡的比較,現有的結果表明無脊椎動物可能存在與脊椎動物差異較大的補體系統,這預示著海洋無脊椎動物具有更多特有的補體激活途徑。
1.6 纖維蛋白原相關蛋白 纖維蛋白原相關蛋白(FREPs)是含有纖維蛋白原相關結構域的凝集素家族,其典型分子結構是在N-末端含有1個或2個免疫球蛋白超家族(IgSF)結構域,接著是1個調節區,在C-末端含有纖維蛋白原(FBG)結構域,具有“非己”識別能力,并能激活免疫應答[13]。研究表明:按蚊中的FREPs-FBN9既能與革蘭氏陽性菌和革蘭氏陰性菌相互作用,還能強烈凝聚位于蚊子腸上皮細胞中的嚙齒動物或人的瘧原蟲,該結果提示FREPs具有防御活性并能與病原體直接作用。FBN9也可以形成二聚體,并能以不同的親和力結合到細菌的表面。研究還發現,通過體外表達扇貝、文昌魚等的FREPs能凝集人的A、B、O型血的紅細胞和雞紅細胞,這種有鈣依賴性的凝集可被乙酰基含水化合物特異性阻斷,并且這些重組蛋白在有鈣離子的條件下還能讓革蘭氏陽性金黃色葡萄球菌以及革蘭氏陰性大腸埃希菌等細菌凝集[14]。因此,FREPs被認為是參與免疫防御最重要的分子之一,在無脊椎動物先天免疫中起著重要作用[15]。
2 無脊椎動物先天免疫系統的模式識別受體
無脊椎動物先天免疫系統作為外源物入侵后宿主防御的最重要防線,首先必須能夠識別“異己”分子,進而快速激活體內相應的信號通路,產生各種效應分子來殺滅、清除外源物。無脊椎動物能夠識別在微生物表面保守的、而在宿主中又不存在的病原相關分子模式(PAMPs),如細菌脂多糖(LPS)、肽聚糖(PG)、細菌DNA及這些生物大分子的衍生物等,識別這些分子模式的受體統稱為模式識別受體(PRRs)。這些PRRs的共同特征是配體的不專一性:一種模式識別受體可以引起不同的免疫防御反應,它們的性質和功能不盡相同(表1)[16]。
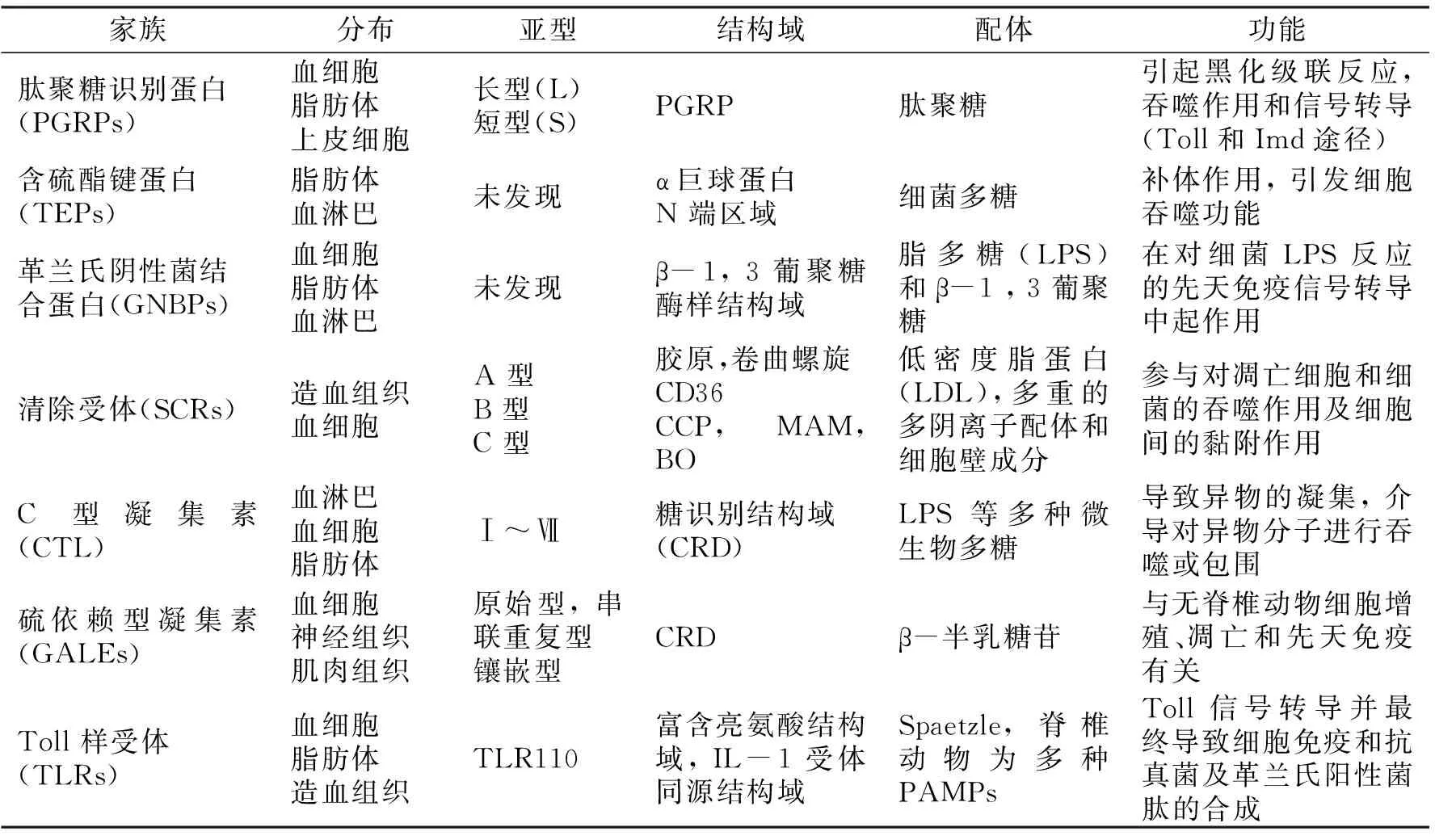
表1 模式識別受體的性質與功能
研究發現,當PAMPs被識別后,這些受體引發激活絲氨酸蛋白酶和解除絲氨酸蛋白酶抑制劑的細胞外級聯反應,將受到感染的信號放大或解除錯誤信號(即信號的調整和放大過程),然后引發信號轉導途徑,激活目的基因的轉錄,最終激活效應物反應系統,產生抗菌肽、酚氧化酶等效應物[17]。PRRs還可以作為促進吞噬的調理素,也可以是凝集、黑化或其他蛋白質修飾級聯反應等免疫過程的起始因子。
