水分梯度下黑河流域荒漠植物群落多樣性特征
2017-08-28 15:14:37張曉龍周繼華蔡文濤管天玉高楠楠姜聯合來利明楊大文叢振濤鄭元潤
生態學報 2017年14期
關鍵詞:物種
張曉龍,周繼華 ,蔡文濤,管天玉 ,高楠楠,杜 會,姜聯合,來利明,楊大文,叢振濤,鄭元潤,*
1 中國科學院植物研究所, 北京 1000932 中國科學院大學, 北京 1000493 清華大學水利水電工程系, 北京 100084
水分梯度下黑河流域荒漠植物群落多樣性特征
張曉龍1,2,周繼華1,2,蔡文濤1,2,管天玉1,2,高楠楠1,2,杜 會1,姜聯合1,來利明1,楊大文3,叢振濤3,鄭元潤1,*
1 中國科學院植物研究所, 北京 1000932 中國科學院大學, 北京 1000493 清華大學水利水電工程系, 北京 100084
物種多樣性對大尺度環境條件的響應是植物生態學的熱點問題,干旱區內陸河流域的特殊環境條件如何影響物種多樣性仍存在較大的不確定性。以黑河流域荒漠植物群落為對象,探討了荒漠植物群落α多樣性和β多樣性對不同水分梯度的響應。結果表明:(1)荒漠植物群落隨水分條件不同而變化,在區域降水梯度上,依次分布有梭梭[Haloxylonammodendron(C. A. Mey.) Bunge]、西伯利亞白刺(NitrariasibiricaPall.)、毛瓣白刺(NitrariapraevisaBobr.)、沙蒿(ArtemisiadesertorumSpreng.)、細枝鹽爪爪(KalidiumgracileFenzl)、珍珠豬毛菜(SalsolapasserinaBunge)和尖葉鹽爪爪[Kalidiumcuspidatum(Ung.Sternb.) Grub.]群落;隨距黑河河道距離增加,主要分布有胡楊(PopuluseuphraticaOliv.)、多枝檉柳(TamarixramosissimaLebed.)、花花柴[Kareliniacaspia(Pall.)Less.]和紅砂[Reaumuriasongarica(Pall.) Maxim.]群落。(2)荒漠植物群落物種多樣性指數偏低,Margalef指數、Shannon-Weiner指數和Simpson指數最大值僅為0.985、1.641、0.596,群落結構簡單,物種組成單一,單寡種優勢群落現象極為明顯,群落相對穩定。(3)在區域降水梯度上,α多樣性指數大致呈先增加后降低的變化趨勢,在珍珠豬毛菜群落(190 mm)達到峰值后迅速下降,S?rensen相似指數在細枝鹽爪爪群落(166 mm)與珍珠豬毛菜群落(190 mm)之間達到最大值,Cody指數相對較小,表明珍珠豬毛菜群落(190 mm)物種豐富度最高,細枝鹽爪爪群落(166 mm)與珍珠豬毛菜群落(190 mm)之間β多樣性最小,物種更替速率最慢,群落處于較穩定階段。(4) 隨距黑河河道距離增加,α多樣性指數大體呈“M”型變化,在多枝檉柳群落(1 km)和多枝檉柳群落(4.1 km)存在兩個峰值,此處物種多樣性相對較高,S?rensen指數在1—1.5 km和4.1—4.8 km之間出現兩個較小值,Cody指數出現兩個峰值,兩者之間β多樣性最大,為物種更替速率相對較快的過渡地帶。對荒漠植被,以重要值為指標計測多樣性指數較為可行,荒漠植物群落的物種多樣性并不是隨水分的增加而直線增加,這對維持和管理荒漠植物群落的多樣性具有重要意義。
黑河流域;荒漠植物群落;α多樣性;β多樣性;水分梯度
植物群落是植物在長期環境變化中自我調節和適應而形成的集合體,作為生態系統物質和能量的主要提供者,是維持生態系統穩定和持續生產的物質基礎[1]。其自身發展總是趨于最大限度地利用當地環境資源,構成多樣的群落結構特征,以維持群落結構和功能的穩定[2- 4]。在植物群落多樣性研究中,群落α多樣性是反映群落內物種組成狀況的一個重要指標,即物種豐富度和均勻度,β多樣性表示物種沿環境梯度發生替代的程度或物種變化的速率,α、β多樣性構成總體多樣性[5- 7]。植物群落α、β多樣性是當前群落生態學乃至整個生態學研究中十分重要的內容[8- 11],研究植物群落多樣性特征對揭示群落、生態系統結構和功能演變具有極為重要的意義[12]。
干旱區內陸河流域是具有獨特生態-水文過程的生態系統,各要素之間以水為紐帶進行著頻繁的物質和能量交換,彼此強烈影響[13-14]。黑河流域作為我國西北干旱區第二大內陸河流域,干流從祁連山發源地到尾閭居延海,橫跨山地、綠洲和荒漠3種不同的地理單元,形成獨特的以水為紐帶的“冰川/凍土—高山植被—山前綠洲—尾閭荒漠”多源景觀以及相伴而生的復合生態系統[15- 17],特殊的自然環境使其成為研究不同水分梯度下植物群落結構、功能和多樣性特征的理想場所。在黑河流域,水分梯度變化主要表現在3個方面,一是上游山區隨海拔變化引起的水分梯度差異,二是山下緯度地帶上隨區域降水變化引起的水分梯度差異,三是沿河道遠近隨地下水位變化引起的水分梯度差異。這3種水分梯度變化均會導致植物群落結構和功能的改變,也會導致植物群落多樣性的變化。黑河中下游地區荒漠分布廣泛,荒漠植物群落是植被的重要組成部分[18- 21],荒漠植物分布和演替受水分條件限制并表現出一定的響應規律[22- 25],水分成為維系荒漠地區植物群落生存和發展的最重要因子。目前,有關荒漠植物群落特征[9-10,26-27]、物種多樣性及其對單一因素如土壤水、地下水等水分變化的響應研究較多[22,28- 30],對于不同類型水分梯度變化對黑河流域荒漠植物群落多樣性的影響則關注較少。在黑河流域,沿自然降水梯度荒漠植物群落α、β多樣性如何變化?垂直河岸帶不同距離區域荒漠植物群落的α、β多樣性又如何變化?這些問題還未得到很好闡釋。
本文以黑河流域荒漠植物群落為研究對象,研究植物群落α、β多樣性沿區域降水和黑河垂直距離遠近的變化規律,探討群落多樣性與水分之間的關系,為更好理解黑河流域生態-水文關系,維持黑河流域荒漠生態系統的穩定性提供科學依據。
1 研究區概況
黑河流域是我國西北典型的內陸河流域,干流發源于祁連山北麓,途徑青海、甘肅和內蒙古三省(區),全長821 km,海拔高程為900—5500 m,流域總面積約14.29x104km2[17,31]。流域地處內陸,氣候惡劣,地形復雜,地勢南高北低,呈連續下降趨勢,地貌景觀大體可分為:冰雪凍土帶、高山植被帶、山前綠洲帶和荒漠帶[16- 17,32]。上游植被茂密、冰川發育,年均降水量200—800 mm,為黑河流域徑流主要形成區,中下游農田荒漠廣布,降水稀少,年均降水量28—200 mm,最小年僅17 mm,為黑河流域徑流的主要消耗區[31]。其中1961年和1992年尾閭西居延海和東居延海相繼干涸,部分河段發生斷流現象,植被生長受干旱脅迫的影響程度較重,生態環境十分脆弱,由于黑河流域生態應急輸水工程的實施,中下游荒漠地區地下水每年都有固定地表河水的補給,使得黑河中下游荒漠地區地下水位不斷升高。有研究表明,2000年以來,下游額濟納綠洲的地下水受到地表河水的補給,地下水埋深基本維持在3 m左右,且年際波動較小[33],基于烏蘭圖格嘎查垂直于河道的7口生態監測井數據,黑河下游(烏蘭圖格)年內最大地下水埋深為3.96 m,最小地下水埋深為1.05 m,地下水埋深與距河道遠近呈正相關關系[34]。
黑河流域植被以荒漠為主,約占流域總面積的76.94%[35],荒漠植物群落分布廣泛,由于受到水分條件的限制,植被分布格局明顯呈斑塊化分布,植被生產力低下,群落結構簡單,種類組成單一。植被類型主要以荒漠戈壁成分為主,喬木有胡楊(PopuluseuphraticaOliv.)、梭梭[Haloxylonammodendron(C. A. Mey.) Bunge];灌木、半灌木有檉柳(Tamarixspp.)、西伯利亞白刺(NitrariasibiricaPall.)、沙蒿(ArtemisiadesertorumSpreng.)和紅砂[Reaumuriasongarica(Pall.) Maxim.]等;草本有苦豆子(SophoraalopecuroidesLinn.)、花花柴[Kareliniacaspia(Pall.) Less.]和駱駝蓬(PeganumharmalaL.)等。
2 研究方法
2.1 野外調查
依據流域區域自然降水及距黑河遠近的水分梯度布設野外觀察樣地,共設置樣地17個,在區域降水梯度上,在流域荒漠區按自然降水梯度隨機設置7個調查樣地,樣地選擇在遠離河流和公路的荒漠地區,自然降水為唯一水分來源,降水數據來自1964—2014年日降水均值[31],由小到大依次記為P1(29 mm)、P2(30 mm)、P3(65 mm)、P4(77 mm)、P5(166 mm)、P6(190 mm)、P7(291 mm)。在沿河梯度上,參照2010年在黑河下游額濟納旗烏蘭圖格嘎查垂直于河道布設的7 口水位監測井,在大體垂直于主河道5.5km范圍內設置10個典型樣地,記為T0.2、T0.5、T1、T1.5、T3.5、T3.7、T3.9、T4.1、T4.8、T5.5,其中,字母T后面的數字代表距離黑河的垂直距離(km),植被類型依據樣地調查資料計算物種重要值確定。
采用隨機取樣法進行樣方調查,每個植被調查樣地設置3個重復樣方進行測定。樣方大小分別為:喬木20 m×20 m、灌木5 m×5 m、草本植物1 m×1 m。記錄樣地基本狀況和群落特征,包括樣地經緯度、海拔、生境、地貌和土壤等屬性,以及植物種類、株數、高度、冠幅、胸(地)徑、密度、蓋度、頻度和優勢種等群落特征。野外調查時間為2015年7—8月。
2.2 數據統計分析
在統計各個樣方每種植物多度、密度、蓋度和頻度的基礎上,分別按喬木、灌木和草本植物計算樣方內各植物物種的重要值,公式為:重要值IV=(相對密度+相對蓋度+相對頻度) /3[36]。
物種多樣性指數較多,在物種重要值基礎上,依據物種多樣性指數應用的廣泛程度,本文選取6種多樣性指數,公式如下:
α多樣性[37-38]
Margalef 豐富度指數
Rm=(S-1) / lnN
Simpson指數(優勢度指數)
Shannon-Wiener多樣性指數
Pielou 指數(均勻度指數)
E=H′/lnS
式中,Pi為種i的相對重要值,N為樣方中物種個體總數,S為樣方內物種數,多樣性指數以重要值計算。
β多樣性[37,39]:
S?rensen指數:
Cody指數:
式中,a和b分別為兩樣方的物種數,c為兩樣方的共有物種數;S?rensen指數反映群落或樣方間物種組成的相似性; Cody指數反映物種沿梯度的替代速率。
3 結果
3.1 植物群落類型與分布
在自然降水梯度上,生境以戈壁平原風蝕殘貌為主,群落結構簡單,物種組成1—7種,植被以半灌木、灌木為主,屬于典型戈壁荒漠植被類型。梭梭群落(P1)和西伯利亞白刺群落(P2)降水量少于50 mm,群落蓋度低,植被稀疏,以單優勢種為主,建群種重要值均為1;毛瓣白刺(NitrariapraevisaBobr.)群落(P3)和沙蒿群落(P4)降水量為50—100 mm,群落結構簡單,群落高度、蓋度較低,建群種重要值為0.76、0.72;細枝鹽爪爪(KalidiumgracileFenzl)群落(P5)和珍珠豬毛菜(SalsolapasserinaBunge)群落(P6)降水量為100—200 mm,群落蓋度較大,物種組成最多,優勢物種明顯,建群種重要值為0.51、0.61;尖葉鹽爪爪[Kalidiumcuspidatum(Ung.Sternb.) Grub.]群落(P7)降水量大于200 mm,群落蓋度最大,建群種重要值為0.69(表1)。
在沿河道距離梯度上,植物群落生境由河岸沙地向荒漠戈壁地貌過渡,植被以喬木、灌木為主,多年生草本也占很重要的地位,群落結構簡單,物種組成1—6種,單優群落較多。有分布于河岸沙地上的胡楊、多枝檉柳、苦豆子和駱駝蓬等,分布于荒漠戈壁上的花花柴和紅砂等。胡楊群落(T0.2)、花花柴群落(T4.8)和紅砂群落(T5.5)以單優種存在,建群種重要值均為1;多枝檉柳群落(T0.5—T4.1),群落蓋度較高,物種組成豐富,建群種重要值為0.46—1;多枝檉柳群落(T1)蓋度最高,物種組成最為豐富,建群種重要值為0.7(表1)。
3.2 區域降水梯度上的植物群落多樣性特征
在區域降水梯度上,植物群落多樣性水平不高,物種組成1—7種,指數值均小于1.7,物種α多樣性指數隨降水增加(P1—P7)大致呈現先增加后減少的變化趨勢,α多樣性指數最大值出現在珍珠豬毛菜群落(P6)樣地(圖1)。Margalef豐富度指數(Rm)在0—0.985間變化,各群落大小依次為珍珠豬毛菜群落(P6)>沙蒿群落(P4)>細枝鹽爪爪群落(P5)>毛瓣白刺群落(P3)>尖葉鹽爪爪群落(P7)>梭梭群落(P2)和西伯利亞白刺群落(P1)。Shannon-Weiner指數在0—1.641之間變化,以珍珠豬毛菜群落(P6)最高(1.641),沙蒿群落(P4)、細枝鹽爪爪群落(P5)、毛瓣白刺群落(P3)和尖葉鹽爪爪群落(P7)等較高(0.598—1.174),Simpson優勢度指數和Pielou均勻度指數分別在0—0.597、0—0.753之間變化,大致與多樣性指數變化趨勢一致。單優勢種梭梭群落(P2)和西伯利亞白刺群落(P1),群落結構最為簡單,多樣性指數、均勻度指數均為0。
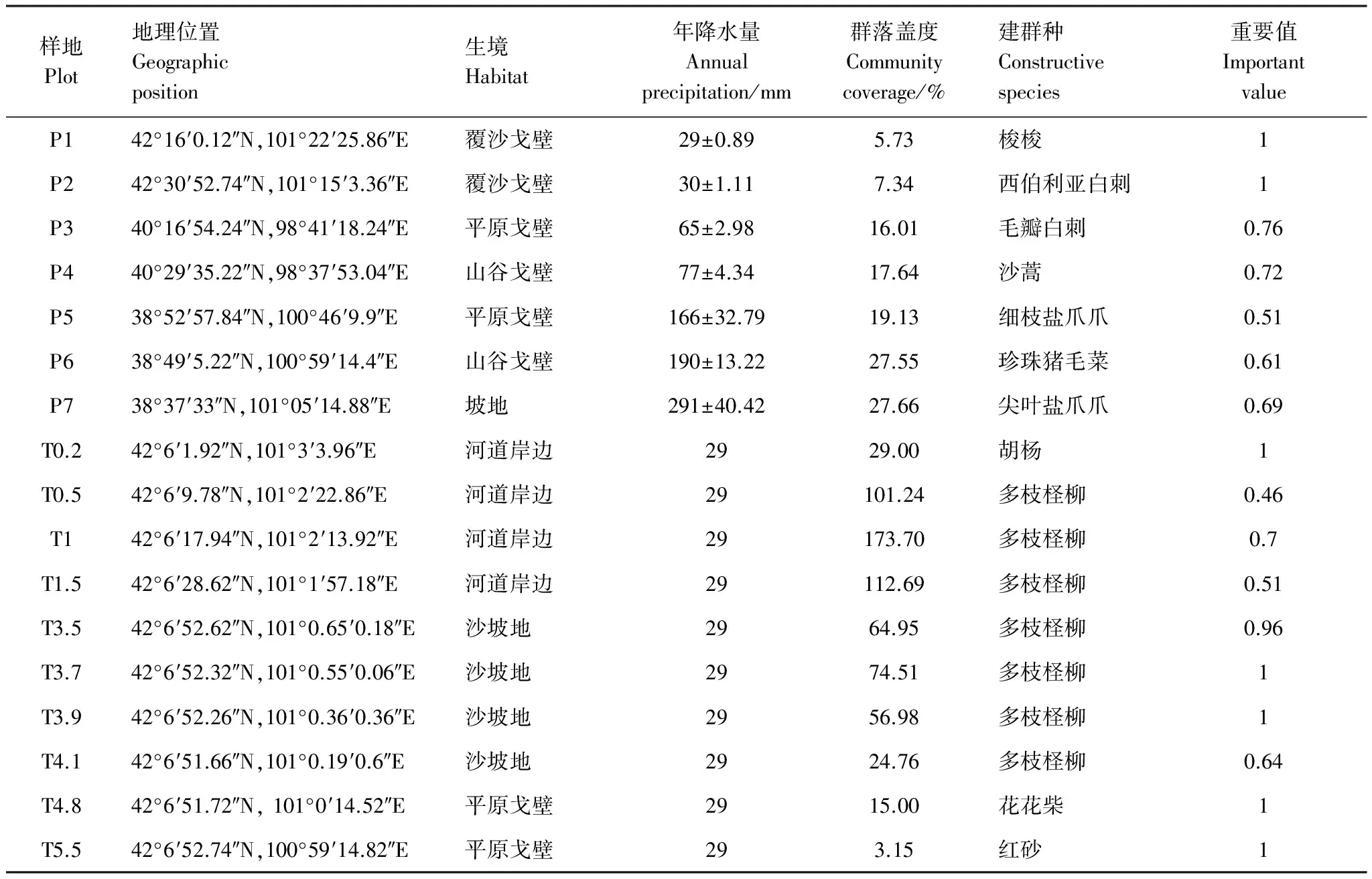
表1 樣地基本信息
P1: 梭梭群落Haloxylonammodendroncommunity; P2: 西伯利亞白刺群落Nitrariasibiricacommunity; P3: 毛瓣白刺群落Nitrariapraevisacommunity; P4: 沙蒿群落Artemisiadesertorumcommunity; P5: 細枝鹽爪爪群落Kalidiumgracilecommunity; P6: 珍珠豬毛菜群落Salsolapasserinacommunity; P7: 尖葉鹽爪爪群落Kalidiumcuspidatumcommunity; T0.2: 胡楊群落Populuseuphraticacommunity;T0.5—T4.1, 多枝檉柳群落Tamarixramosissimacommunity;T4.8, 花花柴群落Kareliniacaspiacommunity; T5.5, 紅砂群落Reaumuriasongaricacommunity
區域降水梯度之間,植物群落S?rensen指數隨降水增加總體上呈先上升后下降的變化趨勢(圖2),細枝鹽爪爪群落(P5)和珍珠豬毛菜群落(P6)之間的S?rensen相似系數最大,為0.66,而梭梭群落(P1)、西伯利亞白刺群落(P2)、毛瓣白刺群落(P3)和沙蒿群落(P4)兩者之間S?rensen相似系數為0。Cody指數與S?rensen指數隨降水的變化趨勢大致相反,細枝鹽爪爪群落(P5)和珍珠豬毛菜群落(P6)之間的Cody指數接近于最小值1。珍珠豬毛菜群落(P6)和尖葉鹽爪爪群落(P7)之間的Cody指數明顯高于其他群落之間的Cody指數,可見,隨著降水增加,植物群落間的物種更替速率先加快、再降低,最終達到最高點。
3.2 沿河梯度下的植物群落多樣性特征
在沿河梯度上,植物群落Shannon-Weiner指數呈“M”型變化趨勢,在距河道近處多枝檉柳群落(T1)呈現出一個最大峰值,迅速下降后,指數值呈現出較大程度的上升趨勢,多枝檉柳群落(T4.1)到達一個較大的峰值,其后隨著離河道距離越來越遠,指數值到達最小,接近于0(圖3)。Simpson多樣性指數和Pielou均勻度指數分別在0—0.564、0—0.726之間變化,大體趨勢與Shannon-Weiner多樣性指數相同。胡楊群落(T0.2)、多枝檉柳群落(T3.5—T3.9)、花花柴群落(T4.8)和紅砂群落(T5.5)幾乎呈單優勢群落,群落結構最為簡單,群落多樣性、優勢度和均勻度水平最低,接近于0。
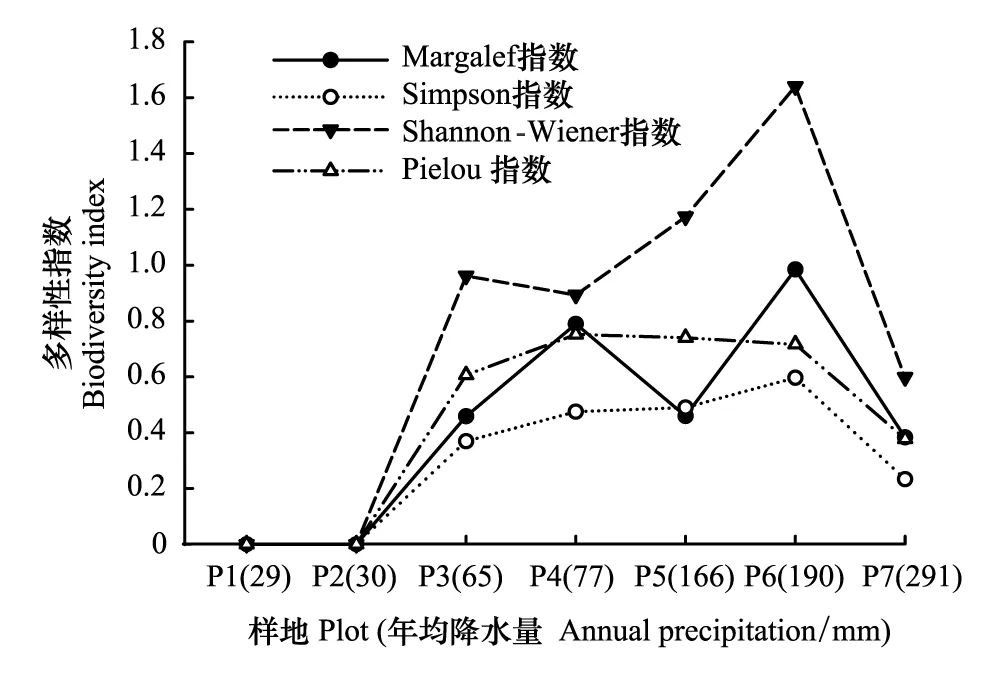
圖1 α多樣性指數隨降水的變化Fig.1 α diversity indices along precipitation gradients P1—P7, 自然降水量呈由小到大趨勢;樣地代碼同表1
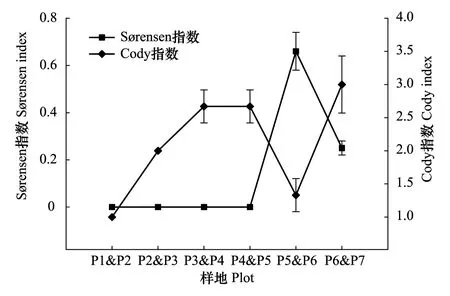
圖2 不同降水梯度植物群落間S?rensen指數和Cody指數Fig.2 S?rensen and Cody indices of plant communities along precipitation gradients樣地代碼同表1
隨距河道距離增加,不同植物群落之間的S?rensen指數總體上呈現先下降再上升后迅速下降的趨勢(圖4)。多枝檉柳群落(T3.7)和多枝檉柳群落(T3.9)為單優群落,群落組分相同,兩者S?rensen相似指數達到最大值,多枝檉柳群落(T1)和多枝檉柳群落(T1.5)之間與多枝檉柳群落(T4.1)和花花柴群落(T4.8)之間S?rensen指數相對較低。植物群落間Cody指數在沿河梯度上的變化趨勢明顯與S?rensen指數相反,多枝檉柳群落(T3.7)和多枝檉柳群落(T3.9)之間Cody指數達到最小值0,多枝檉柳群落(T1)和多枝檉柳群落(T1.5)之間與多枝檉柳群落(T4.1)和花花柴群落(T4.8)之間Cody指數值明顯高于其它群落。

圖3 α多樣性指數隨沿河距離的變化Fig.3 α diversity indices along vertical distance to river
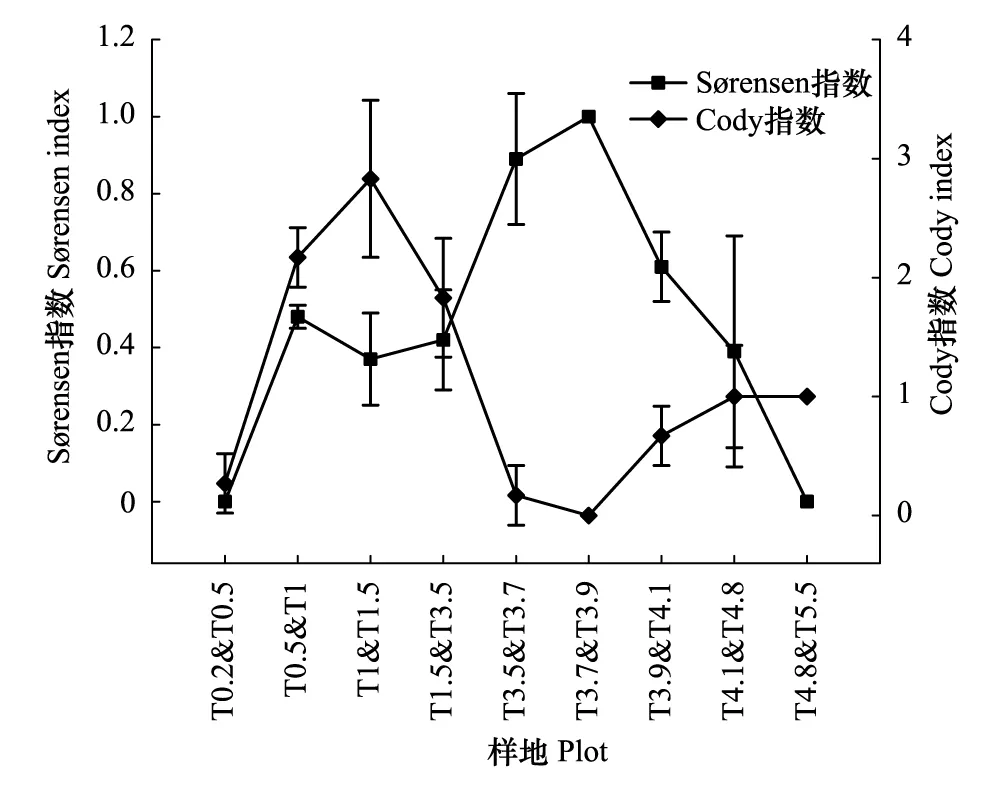
圖4 沿河梯度植物群落間S?rensen指數和Cody指數Fig.4 S?rensen and Cody indices of plant communities along vertical distance to river (樣地代碼同表1)
4 討論
本研究聚焦于黑河流域在區域降水和沿河道距離遠近兩個梯度上荒漠植物群落的多樣性特征。總體上,荒漠植物群落結構簡單,物種組成稀少,17個荒漠植物群落中只有5個群落物種組成大于3種,單寡種優勢群落現象極為明顯,梭梭群落、西伯利亞白刺群落、胡楊群落等均為單優勢種群落,這與荒漠地區干旱惡劣的生境條件有關[40]。植物群落多樣性指數可以客觀反映群落內物種組成特征,在所有水分梯度上,植物群落α多樣性指數偏低,Margalef物種豐富度指數、Shannon-Weiner多樣性指數和Simpson優勢度指數最大值僅為0.985、1.641、0.596,植物群落物種多樣性與其它荒漠區植被物種多樣性水平相當[9,22,41-42],說明荒漠植物群落物種多樣性水平普遍較低,進一步反映了該地區植物群落結構簡單、物種組成稀少且種類分布極不均勻的荒漠植物群落特點。荒漠植物群落蓋度隨降水量增加明顯呈現上升趨勢,植被蓋度的大小與降水量呈正相關關系(表1),表明荒漠植物群落蓋度變化對降水量的響應非常敏感。在沿河梯度上,有研究表明,距河道1.2 km處植被蓋度最大[43],本研究發現植物群落蓋度最大值出現在多枝檉柳群落(T1),而不是在離河道最近的胡楊群落(T0.2),表明距河道最近植物群落蓋度并非最高。
在荒漠地區,以水分梯度為特征的物種多樣性格局和分布構建了荒漠生態系統的結構和功能,水分是荒漠地區生態-水文過程的關鍵影響因子和主要驅動力,而物種多樣性對水分條件變化的響應直接影響著荒漠生態系統的功能和穩定性[44]。有研究表明,結構簡單、單寡種優勢荒漠植物群落往往具有較高的穩定性,物種多樣性低并不代表系統穩定性差[29,45],在不同水分梯度上,如苦豆子和多枝檉柳等群落物種組成較豐富,僅分布在水分條件較好的河岸周圍,處于不穩定狀態,近年來,沿河地帶都有上中游固定地表河水的補給,植物群落處于較穩定狀態,水分是影響其生存和發展的主要因素。紅砂、梭梭和西伯利亞白刺等群落物種組成單一,地處極度干旱地區,形成穩定的群落結構,這可能是荒漠植物群落長期適應干旱風沙環境而形成的適應對策。
在區域降水梯度上,荒漠植物群落α多樣性指數隨降水量增加總體呈先增加后降低的變化趨勢,說明水分條件是限制荒漠植物群落生存和發展的最主要因子。α多樣性指數在珍珠豬毛菜群落(190 mm)達到最大值,在降水量持續增大(大于200 mm)的情況下,α多樣性指數在尖葉鹽爪爪群落(291 mm)迅速下降。植物群落的物種多樣性并不是隨降水量的增加而直線增加,出現上述情況可能與尖葉鹽爪爪群落自身及所處生境有關,尖葉鹽爪爪屬于典型的泌鹽植物[46],所處環境鹽分多集中在地表,形成鹽結皮,含鹽量可達35%—37%,在鹽生環境條件下其它植物生長受限,尖葉鹽爪爪形成單寡種優群落,從而導致尖葉鹽爪爪群落(P7)多樣指數低于珍珠豬毛菜群落(P6)樣地。
在沿河梯度上,α多樣性指數隨距河道距離增加大體呈“M”型變化趨勢,在多枝檉柳群落(T1)和多枝檉柳群落(T4.1)存在兩個峰值,距河較近的胡楊群落(T0.2)物種多樣性并非最高。參照2010年在黑河下游額濟納旗烏蘭圖格嘎查垂直于河道布設的7 口水位監測井數據,隨距河道距離增加地下水埋深逐漸加深,植物群落物種多樣性并不是隨著地下水變化而呈直線變化趨勢。理論上在一定沿河地帶范圍內,離河較近地區,土壤水分條件最好,植物多樣性應該越高[11],這與本文結果有所不同,這主要與高地下水位變化引起土壤性質變化有關。胡楊群落(T0.2)地下水埋深為1.05—2.6 m[34],所處生境土壤水分條件好,地下水位高,土壤蒸發旺盛導致近河岸帶淺層土壤鹽分集聚,使植物群落面臨鹽堿脅迫[47],這對深根系植物胡楊和檉柳的生長有利,對于淺根和不耐鹽堿的植物不利[48],因此,河岸邊形成以胡楊占絕對優勢的胡楊群落,植物群落的物種多樣性水平較低。隨距河道距離增加,地下水位下降,苦豆子、花花柴和駱駝蓬等草本植物出現,使其物種豐富度達到最大,植物群落物種多樣性最大值出現在沿河梯度1—1.5 km,在此區域內適宜的土壤水分及較低的土壤鹽分維持了較高的物種多樣性。隨距河道距離的進一步增加,地下水位持續下降,土壤水分明顯下降,不能維持較高的物種多樣性,物種多樣性指數迅速下降。在降水梯度上,隨著降水量的增加,物種多樣性隨降水增加,先增加后降低,在沿河梯度上,隨水分條件變化,總體上呈先增加后降低的趨勢,其中在沿河梯度上多枝檉柳群落(T4.1)樣地α多樣性指數有所升高,可能與“檉柳-黑果枸杞-花花柴”灌木草本植物組合存在有一定關系。有研究表明,檉柳主要吸收200—300 cm的深層土壤水,多年生草本植物花花柴主要吸收50—100 cm的中層土壤水,檉柳與其它物種之間水分利用存在互助關系[49-50],在土壤含水量較低的情況下,群落中各物種可以長期共存,提高對水資源的利用率,增加群落的穩定性,從而保持T4.1處較高的物種豐富度。
β多樣性用于反映環境梯度下植物群落間的差異性,當一個群落向另一個群落過渡時,環境梯度變化較大,不同群落或樣方間共有種越少,β多樣性越大[51]。在降水梯度上,梭梭群落(P1)和西伯利亞白刺群落(P2)、西伯利亞白刺群落(P2)和毛瓣白刺群落(P3)、毛瓣白刺群落(P3)和沙蒿群落(P4)之間 S?rensen相似系數為0,原因在于在降水量小于100mm的荒漠地區,群落間生境差異較大,植物群落多為單寡種優勢群落,群落間物種相似性低。隨著降水量的增加,群落間的S?rensen相似系數在細枝鹽爪爪群落(P5)和珍珠豬毛菜群落(P6)之間達到最大值0.66,然而在珍珠豬毛菜群落(P6)和尖葉鹽爪爪群落(P7)之間迅速降低到0.25,同時Cody指數從1.33增加到3,說明細枝鹽爪爪群落(P5)和珍珠豬毛菜群落(P6)之間β多樣性最小,即細枝鹽爪爪群落(P5)和珍珠豬毛菜群落(P6)所處區域荒漠植物群落結構最為穩定,物種更替速率最小。在沿河梯度上,S?rensen相似系數大體上呈現先下降再上升后迅速下降的趨勢,Cody指數變化趨勢與之相反。S?rensen相似系數在多枝檉柳群落(T1)和多枝檉柳群落(T1.5)之間和多枝檉柳群落(T4.1)和花花柴群落(T4.8)之間出現兩個較小值,Cody指數則在兩者出現兩個峰值,說明多枝檉柳群落(T1)和多枝檉柳群落(T1.5)之間和多枝檉柳群落(T4.1)和花花柴群落(T4.8)之間β多樣性最大,物種更替速率相對較快,T1&T1.5之間和T4.1&T4.8之間過渡性質明顯,有研究表明,距黑河河道1.2 km處為綠洲向荒漠過渡區過渡的地帶,距河道4.3 km處為荒漠過渡區和荒漠區的過渡地帶[43],這與本研究結果相符。由于荒漠生境惡劣,受調查季節和空間所限,各植物群落與環境因子之間定量關系的分析有待深入研究。
[1] Lieth H. Modeling the primary productivity of the world//Lieth H, Whittaker R, eds. Primary Productivity of the Biosphere. New York: Springer-Verlag, 1975: 237- 263.
[2] Levine J M. Complex interactions in a streamside plant community. Ecology, 2000, 81(12): 3431- 3444.
[3] Lomolino M V. Elevation gradients of species-density: historical and prospective views. Global Ecology and Biogeography, 2001, 10(1): 3- 13.
[4] Lortie C J, Brooker R W, Chole P, Kikvidze Z, Michalet R, Pugnaire F I, Callaway R M. Rethinking plant community theory. Oikos, 2004, 107(2): 433- 438.
[5] 馬克平, 黃建輝, 于順利, 陳靈芝. 北京東靈山地區植物群落多樣性的研究Ⅱ豐富度、均勻度和物種多樣性指數. 生態學報, 1995, 15(3): 268- 277.
[6] 馬克平, 劉燦然, 劉玉明. 生物群落多樣性的測度方法Ⅱβ多樣性的測度方法. 生物多樣性, 1995, 3(1): 38- 43.
[7] 張金屯. 數量生態學. 北京: 科學出版社, 2011: 77- 98.
[8] Franklin J F. Preserving biodiversity: species in landscapes: response. Ecological Applications, 1994, 4(2): 208- 209.
[9] 馬斌, 周志宇, 張莉麗, 高文星, 陳善科, 張寶林. 阿拉善左旗植物物種多樣性空間分布特征. 生態學報, 2008, 28(12): 6099- 6106.
[10] 陳杰, 郭屹立, 盧訓令, 丁圣彥, 蘇思, 郭靜靜, 李乾璽. 伊洛河流域草本植物群落物種多樣性. 生態學報, 2012, 32(10): 3021- 3030.
[11] 張雪妮, 呂光輝, 王庭權, 馬玉, 阿布里孜·阿不都熱合曼, 趙曉英, 郭振潔, 朱修逸. 荒漠區垂直河岸帶植物多樣性格局及其成因. 生態學報, 2015, 35(18): 5966- 5974.
[12] Pandey S K, Shukla R P. Plant diversity in managed sal (ShorearobustaGaertn.) forests of Gorakhpur, India: species composition, regeneration and conservation. Biodiversity and Conservation, 2003, 12(11): 2295- 2319.
[13] 趙文智, 程國棟. 干旱區生態水文過程研究若干問題評述. 科學通報, 2001, 46(22): 1851- 1857.
[14] Montaldo N, Rondena R, Albertson J D, Mancini M. Parsimonious modeling of vegetation dynamics for ecohydrologic studies of water-limited ecosystems. Water Resources Research, 2005, 41(10): W10416.
[15] 盧玲, 李新, 程國棟, 肖洪浪. 黑河流域景觀結構分析. 生態學報, 2001, 21(8): 1217- 1224.
[16] 盧玲, 程國棟, 李新. 黑河流域中游地區景觀變化研究. 應用生態學報, 2001, 12(1): 68- 74.
[17] 黑河計劃數據管理中心. 黑河流域邊界數據集. 蘭州: 中科院寒區旱區環境與工程研究所, 2011.
[18] 斯琴巴特爾, 秀敏. 荒漠植物蒙古扁桃水分生理特征. 植物生態學報, 2007, 31(3): 484- 489.
[19] 趙良菊, 肖洪浪, 程國棟, 宋耀選, 趙亮, 李彩芝, 楊秋. 黑河下游河岸林植物水分來源初步研究. 地球學報, 2008, 29(6): 709- 718.
[20] Odum E P. The strategy of ecosystem development. Science, 1969, 164(3877): 262- 270.
[21] Ricklefs R E. Community diversity: relative roles of local and regional processes. Science, 1987, 235(4785): 167- 171.
[22] 張佩, 袁國富, 莊偉, 薛沙沙. 黑河中游荒漠綠洲過渡帶多枝檉柳對地下水位變化的生理生態響應與適應. 生態學報, 2011, 31(22): 6677- 6687.
[23] 劉加珍, 李衛紅, 吳純淵, 陳永金. 荒漠河岸生態系統退化與物種多樣性恢復研究. 干旱區研究, 2009, 26(2): 212- 220.
[24] 張海娜, 蘇培璽, 李善家, 周紫鵑, 解婷婷, 趙慶芳. 荒漠區植物光合器官解剖結構對水分利用效率的指示作用. 生態學報, 2013, 33(16): 4909- 4918.
[25] 周曉兵, 張元明, 王莎莎, 張丙昌. 模擬氮沉降和干旱對準噶爾盆地兩種一年生荒漠植物生長和光合生理的影響. 植物生態學報, 2010, 34(12): 1394- 1403.
[26] 魚騰飛, 馮起, 司建華, 席海洋. 黑河下游額濟納綠洲植物群落特征與物種多樣性研究. 西北植物學報, 2011, 31(5): 1032- 1038.
[27] 王國宏, 任繼周, 張自和. 河西山地綠洲荒漠植物群落種群多樣性研究I生態地理及植物群落的基本特征. 草業學報, 2001, 10(1): 1- 12.
[28] 朱軍濤, 于靜潔, 王平, 王志勇. 額濟納荒漠綠洲植物群落的數量分類及其與地下水環境的關系分析. 植物生態學報, 2011, 35(5): 480- 489.
[29] 李新榮, 何明珠, 賈榮亮. 黑河中下游荒漠區植物多樣性分布對土壤水分變化的響應. 地球科學進展, 2008, 23(7): 685- 691.
[30] 陳亞寧, 張宏鋒, 李衛紅, 陳亞鵬. 新疆塔里木河下游物種多樣性變化與地下水位的關系. 地球科學進展, 2005, 20(2): 158- 165.
[31] Gao B, Qin Y, Wang Y H, Yang D W, Zheng Y R. Modeling ecohydrological processes and spatial patterns in the upper Heihe Basin in China. Forests, 2016, 7(1), doi: 10.3390/f7010010.
[32] 康爾泗, 程國棟, 藍永超, 金會軍. 西北干旱區內陸河流域出山徑流變化趨勢對氣候變化響應模型. 中國科學(D輯), 1999, 29(S1): 47- 54.
[33] 蔣曉輝, 劉昌明. 黑河下游植被對調水的響應. 地理學報, 2009, 64(7): 791- 797.
[34] 周洪華, 李衛紅, 木巴熱克·阿尤普, 徐茜. 荒漠河岸林植物木質部導水與栓塞特征及其對干旱脅迫的響應. 植物生態學報, 2012, 36(1): 19- 29.
[35] 王建華, 趙軍, 胡曉利, 蓋春梅. 2012年新編黑河流域2000年土地利用/土地覆蓋數據集. 黑河: 黑河計劃數據管理中心, 2014.
[36] 方精云, 王襄平, 沈澤昊, 唐志堯, 賀金生, 于丹, 江源, 王志恒, 鄭成洋, 朱江玲, 郭兆迪. 植物群落清查的主要內容、 方法和技術規范. 生物多樣性, 2009, 17(6): 533- 548
[37] Magurran A E. Ecological Diversity and Its Measurement. New Jersey: Princeton University Press, 1988.
[38] 錢迎倩, 馬克平. 生物多樣性研究的原理與方法. 北京: 中國科學技術出版社, 1994: 141- 165.
[39] Whittaker R H. Evolution and measurement of species diversity. Taxon, 1972, 21(2/3): 213- 251.
[40] 李新榮, 張新時. 鄂爾多斯高原荒漠化草原與草原化荒漠灌木類群生物多樣性的研究. 應用生態學報, 1999, 10(6): 665- 669.
[41] 張錦春, 王繼和, 趙明, 劉虎俊, 廖空太, 徐先英. 庫姆塔格沙漠南緣荒漠植物群落多樣性分析. 植物生態學報, 2006, 30(3): 375- 382.
[42] 張林靜, 岳明, 張遠東, 顧峰雪, 潘曉玲, 趙桂仿. 新疆阜康綠洲荒漠過渡帶植物群落物種多樣性特征. 地理科學, 2003, 23(3): 329- 334.
[43] 路明, 劉加珍, 陳永金, 路亞坤. 黑河流域下游綠洲-荒漠梯度上的植被數量特征. 西北農業學報, 2013, 22(6): 176- 182.
[44] Whitford W G. Ecology of Desert Systems. New York: Academic Press, 2002.
[45] 李文華, 趙景柱. 生態學研究回顧與展望. 北京: 氣象出版社, 2004: 625- 649.
[46] 章英才, 張晉寧. 尖葉鹽爪爪葉的解剖結構與分析. 寧夏農學院學報, 2004, 25(2): 43- 45, 50- 50.
[47] 趙文智, 劉鵠. 荒漠區植被對地下水埋深響應研究進展. 生態學報, 2006, 26(8): 2702- 2708.
[48] 鄭丹, 李衛紅, 陳亞鵬, 劉加珍. 干旱區地下水與天然植被關系研究綜述. 資源科學, 2005, 27(4): 160- 167.
[49] 陳小麗, 陳亞寧, 陳亞鵬. 黑河下游荒漠河岸林植物水分利用關系研究. 中國生態農業學報, 2014, 22(8): 972- 979.
[50] 付愛紅, 陳亞寧, 李衛紅. 中國黑河下游荒漠河岸林植物群落水分利用策略研究. 中國科學: 地球科學, 2014, 44(4): 693- 705.
[51] 王慶鎖, 馮宗煒, 羅菊春. 河北北部、內蒙古東部森林-草原交錯帶生物多樣性研究. 植物生態學報, 2000, 24(2): 141- 146.
Diversity characteristics of plant communities in the arid desert of the Heihe basin under different moisture gradients
ZHANG Xiaolong1,2, ZHOU Jihua1,2, CAI Wentao1,2, GUAN Tianyu1,2, GAO Nannan1,2, DU Hui1, JIANG Lianhe1, LAI Liming1, YANG Dawen3, CONG Zhentao3, ZHENG Yuanrun1,*
1InstituteofBotany,ChineseAcademyofSciences,Beijing100093,China2UniversityofChineseAcademyofSciences,Beijing100049,China3DepartmentofHydraulicEngineering,TsinghuaUniversity,Beijing100084,China
The responses of species diversity to the large-scale environment represent an important issue in plant ecology. In arid regions, the influence of the unique environmental conditions of inland river basins on species diversity remains uncertain. We examined the characteristics of plant community alpha and beta diversity under different moisture gradients in plant communities of the arid desert of the Heihe basin, Northwest China. The results showed that:(1) the type of desert plant community changed with moisture gradients. The communities which were most distinguished based on important value by increasing precipitation gradients were communities dominated byHaloxylonammodendron,Nitrariasibirica,Nitrariapraevisa,Artemisiadesertorum,Kalidiumgracile,Salsolapasserina, andKalidiumcuspidatum. The communities ofPopuluseuphratica,Tamarixramosissima,Kareliniacaspia, andReaumuriasongaricawere distributed along the moisture gradient, with species listed in order of increasing distance of their communities from rivers. (2) The species diversity indexes of desert plant communities were low; the maximum Margalef, Shannon-Wiener, and Simpson values were only 0.985, 1.641, and 0.596, respectively. The structure and composition of the studied communities were simple, and the communities were relatively stable. Moreover, monotypic (oligotypic) community types were prevalent. (3) Under the gradient of regional increasing precipitation,αdiversity showed a trend towards increasing and then decreasing; there was a high-value area around the community ofSalsolapasserina(190 mm), but then values decreased rapidly. The S?rensen index reached a maximum in the communities ofKalidiumgracile(166 mm) andSalsolapasserina(190 mm), while the Cody index remained relatively small. This suggests that the species richness was highest in theSalsolapasserinacommunity (190 mm). Theβdiversity reached minimum values in theKalidiumgracile(166 mm) andSalsolapasserina(190 mm) communities. The species replacement rate was also the lowest in these communities, indicating that these communities were relatively stable. (4) Theαdiversity showed a trend towards “M” type change with increasing distance from the river, and reached two peaks at around the community ofTamarixramosissima(1km) and the community ofTamarixramosissima(4.1 km). There were two smaller S?rensen index values approximately 1—1.5 km and 4.1—4.8 km, and the Cody index value showed a trend opposite to that of the S?rensen index. Furthermore, theβdiversity values were high, which suggests that these were transitional zones. We concluded that the diversity index based on important value was more applicable in arid desert areas and that the species diversity of the desert plant community showed no linear increase with increasing moisture. These results are important for maintaining and managing the diversity of desert plant communities.
Heihe basin; desert plant community;αdiversity;βdiversity; moisture gradients
國家自然科學基金項目(91225302)
2016- 04- 08; 網絡出版日期:2017- 03- 02
10.5846/stxb201604080643
*通訊作者Corresponding author.E-mail: zhengyr@ibcas.ac.cn
張曉龍,周繼華,蔡文濤,管天玉,高楠楠,杜會,姜聯合,來利明,楊大文,叢振濤,鄭元潤.水分梯度下黑河流域荒漠植物群落多樣性特征.生態學報,2017,37(14):4627- 4635.
Zhang X L, Zhou J H, Cai W T, Guan T Y, Gao N N, Du H, Jiang L H, Lai L M, Yang D W, Cong Z T, Zheng Y R.Diversity characteristics of plant communities in the arid desert of the Heihe basin under different moisture gradients.Acta Ecologica Sinica,2017,37(14):4627- 4635.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
