吉林蛟河闊葉紅松林樣地種-面積關(guān)系
2017-08-28 15:14:37陳桂蓮譚凌照范春雨張新娜張春雨趙秀海
生態(tài)學(xué)報(bào) 2017年14期
陳桂蓮,譚凌照,范春雨,張新娜,張春雨,趙秀海,*
1 北京林業(yè)大學(xué)林學(xué)院,北京 1000832 森林資源生態(tài)系統(tǒng)過(guò)程北京市重點(diǎn)實(shí)驗(yàn)室,北京 100083
吉林蛟河闊葉紅松林樣地種-面積關(guān)系
陳桂蓮1,2,譚凌照1,2,范春雨1,2,張新娜1,2,張春雨1,2,趙秀海1,2,*
1 北京林業(yè)大學(xué)林學(xué)院,北京 1000832 森林資源生態(tài)系統(tǒng)過(guò)程北京市重點(diǎn)實(shí)驗(yàn)室,北京 100083
種-面積關(guān)系是群落生態(tài)學(xué)的核心問(wèn)題之一,是生物多樣性尺度轉(zhuǎn)換的重要依據(jù)。利用吉林蛟河闊葉紅松林30 hm2的樣地?cái)?shù)據(jù),采用隨機(jī)取樣與巢式取樣方法,分別在10、20、30 hm2尺度上建立對(duì)數(shù)模型(Logarithmic function)、冪函數(shù)模型(Power function)和邏輯斯蒂模型(Logistic function)擬合局域種-面積關(guān)系,并利用赤池信息準(zhǔn)則(AIC)進(jìn)行擬合結(jié)果優(yōu)度檢驗(yàn)。結(jié)果表明,取樣方法對(duì)種-面積關(guān)系的構(gòu)建有顯著影響,隨機(jī)取樣優(yōu)于巢式取樣。種-面積關(guān)系的構(gòu)建與尺度(取樣上限)密切相關(guān):在小尺度上(10 hm2),對(duì)數(shù)模型與邏輯斯蒂模型擬合效果優(yōu)于冪函數(shù)模型;在中尺度和大尺度上(20、30 hm2),相對(duì)于對(duì)數(shù)模型和冪函數(shù)模型,邏輯斯蒂模型能更好地?cái)M合闊葉紅松林的種-面積關(guān)系。據(jù)AIC值可知,隨機(jī)取樣下的邏輯斯蒂模型擬合效果最好,是擬合30 hm2闊葉紅松林樣地種-面積關(guān)系的最適模型。因此研究時(shí)需要根據(jù)區(qū)域森林群落的實(shí)際情況選擇種-面積模型。
種-面積關(guān)系;擬合優(yōu)度;取樣方法;取樣尺度
種-面積關(guān)系描述了物種隨取樣面積的增加而變化的規(guī)律,是群落生態(tài)學(xué)研究的核心問(wèn)題之一[1]。種-面積關(guān)系可以解決許多生態(tài)問(wèn)題,諸如確定群落的最小面積,評(píng)估區(qū)域物種多樣性,確定自然保護(hù)區(qū)范圍,預(yù)測(cè)生境破壞對(duì)物種豐富度影響等[2- 6]。種-面積關(guān)系相較于用單一的物種數(shù)目,可以更好地描述植物群落的物種多樣性[7],對(duì)區(qū)域生物保護(hù)以及物種多樣性維持具有重要意義[8]。
目前為止,生態(tài)學(xué)家們提出了上百種種-面積關(guān)系的擬合方程,其中應(yīng)用最廣泛的分別是冪函數(shù)模型、對(duì)數(shù)模型和邏輯斯蒂模型,這3種模型因其參數(shù)具有明確的生態(tài)學(xué)含義并且簡(jiǎn)單實(shí)用,一直以來(lái)被科學(xué)家們廣泛接受[9-10]。但以上3種模型對(duì)種-面積關(guān)系擬合的準(zhǔn)確性受取樣尺度和取樣方法的影響較大。種-面積關(guān)系應(yīng)用哪個(gè)模型擬合最優(yōu),目前還飽受爭(zhēng)議[10]。
大量研究結(jié)果表明,種-面積關(guān)系會(huì)隨著取樣尺度不同而發(fā)生變化[11],主要原因在于生境過(guò)濾效應(yīng)以及擴(kuò)散限制等生態(tài)過(guò)程影響不同尺度上的物種空間分布格局[12- 14]。一些研究表明,隨著尺度變化,在小尺度和中尺度上適合采用冪函數(shù)模型,而在大尺度上適合采用邏輯斯蒂模型[15- 16]。另一些研究,如:Fisher則認(rèn)為在小尺度上對(duì)數(shù)模型能夠更好地?cái)M合種-面積曲線[17]。進(jìn)行多尺度上的種-面積關(guān)系研究對(duì)于準(zhǔn)確反映物種豐富度和空間分布格局背后的生態(tài)學(xué)機(jī)制具有重要意義。
目前的研究著重從生態(tài)學(xué)理論角度討論種-面積關(guān)系的尺度效應(yīng)而忽略了生態(tài)學(xué)方法對(duì)種-面積關(guān)系的影響,有研究證明,取樣方法是影響種-面積關(guān)系的重要因素[18]。目前最常用的取樣方式包括巢式取樣和隨機(jī)取樣[19-20]。巢式取樣按一定規(guī)則不斷擴(kuò)大取樣面積,其中大樣方包括小樣方,其研究尺度可從群落擴(kuò)展至區(qū)域乃至大洲[21],成為構(gòu)建種-面積關(guān)系的主要方式。但Gleason等認(rèn)為,巢式取樣在構(gòu)建種-面積關(guān)系上易受稀有種、物種聚集以及空間異質(zhì)性等因素的影響[22],會(huì)降低預(yù)測(cè)物種數(shù)的準(zhǔn)確性。隨機(jī)取樣即在研究區(qū)域隨機(jī)選取規(guī)定面積的樣方來(lái)構(gòu)建種-面積關(guān)系曲線,可用來(lái)估計(jì)區(qū)域種庫(kù)的大小[23],在當(dāng)前的研究中應(yīng)用越來(lái)越廣泛。
闊葉紅松林是我國(guó)溫帶森林典型的群落類型,與其他同緯度的森林群落相比,物種豐富度高,建群種比較獨(dú)特,建立闊葉紅松林的種-面積關(guān)系對(duì)反映東北主要植被類型多樣性格局和多樣性保護(hù)具有重要意義[24],是當(dāng)前種-面積關(guān)系研究不足之處的補(bǔ)充案例。本文基于吉林蛟河30 hm2闊葉紅松林固定監(jiān)測(cè)樣地,利用不同尺度下巢式取樣和隨機(jī)取樣數(shù)據(jù),建立了基于冪函數(shù)模型、對(duì)數(shù)模型和邏輯斯蒂模型的區(qū)域種-面積關(guān)系曲線,目的在于探討樣地內(nèi)取樣方法和取樣尺度對(duì)種-面積關(guān)系的影響。
1 研究區(qū)概況

圖1 蛟河 30 hm2闊葉紅松林樣地地形模擬圖 Fig.1 Terrain simulation diagram of the 30 hm2 broad-leaved korean pine forest sample area in Jiaohe
研究區(qū)域位于吉林省蛟河林業(yè)實(shí)驗(yàn)區(qū)管理局,處于吉林省蛟河市前進(jìn)鄉(xiāng)境內(nèi),屬于長(zhǎng)白山系張廣才嶺山脈(地理位置:43°57.928′—43°58.214′N,127°45.287′—127°45.790′E),海拔為576.03—748.18 m,最高點(diǎn)與最低點(diǎn)高差達(dá)172.15 m(圖1)。研究區(qū)域?yàn)榧撅L(fēng)性溫帶大陸性氣候,年均氣溫為3.8℃。每年7月為最熱月,平均氣溫21.7℃,1月為最冷月,平均氣溫-15.4℃。年均降雨量695.9 mm。樣地類型為闊葉紅松林,屬于長(zhǎng)白山植被區(qū)系,該樣地的土壤類型為暗棕色森林土,厚度為20—100 cm。林分的平均胸徑為10 cm,最大胸徑達(dá)114.5 cm,DBH ≤5 cm的個(gè)體占全部個(gè)體的51.57%。喬木樹種主要包括紅松(Pinuskoraiensis)、裂葉榆(Ulmuslaciniata)、紫椴(Tiliaamurensis)、色木槭(Acermono)、白牛槭(Acermandshuricum)、胡桃楸(Juglansmandshurica)、水曲柳(Fraxinusmandshurica)、白樺(Betulaplatyphylla)等,灌木主要包括暴馬丁香(Syringareticulata)、毛榛(Corylusmandshurica) 、瘤枝衛(wèi)矛(Euonymuspauciflorus) 和東北鼠李(Rhamnusdavurica)等,草本主要包括白花碎米薺(Cardamineleucantha)、北重樓(Parisverticillata)、荷青花(HylomeconJaponicum)、東北延胡索(Corydalisambigua)、多被銀蓮花(Anemoneraddeana)等。
2 研究方法
2.1 樣地設(shè)置與數(shù)據(jù)調(diào)查
2010年建立30 hm2(500 m×600 m)闊葉紅松林固定監(jiān)測(cè)樣地。將樣地劃分為750個(gè)20 m×20 m的連續(xù)樣方。在每個(gè)樣方中調(diào)查樣地內(nèi)所有DBH ≥1 cm的木本植物,記錄其種名、胸徑、樹高、冠幅(東西冠幅及南北冠幅)和其在整個(gè)樣地內(nèi)相對(duì)應(yīng)的位置坐標(biāo),并進(jìn)行掛牌標(biāo)記。
2.2 取樣尺度
基于研究樣地面積,在30 hm2范圍內(nèi)隨機(jī)選取10、20 hm2的研究區(qū)域,以10、20、30 hm2分別作為小尺度、中尺度、大尺度上構(gòu)建種-面積關(guān)系的上限。
2.3 取樣方法
以30 hm2取樣尺度為例:
巢式取樣:在研究區(qū)域(500 m×600 m)內(nèi)隨機(jī)選取一點(diǎn),按照5 m×5 m、10 m×10 m、15 m×15 m、…、495 m ×495 m、500 m×500 m、500 m×540 m、500 m×580 m 、500 m×600 m的嵌套方法進(jìn)行取樣,在抽取的樣方中隨機(jī)選取30個(gè),分別統(tǒng)計(jì)各樣方中的物種數(shù),構(gòu)建巢式取樣的種-面積關(guān)系。在取樣時(shí)若取樣單元超出研究區(qū)域,則對(duì)無(wú)效取樣進(jìn)行剔除,重新取樣。
隨機(jī)取樣:在研究區(qū)域(500 m×600 m)內(nèi)以5 m×5 m、10 m×10 m、15 m×15 m…495 m×495 m、500 m×500 m、500 m×540 m、500 m×580 m 、500 m×600 m為規(guī)格進(jìn)行隨機(jī)取樣,在抽取的樣方中隨機(jī)選取30個(gè),記錄每個(gè)樣方內(nèi)的物種數(shù),構(gòu)建隨機(jī)取樣的種-面積關(guān)系。
由于取樣面積變化梯度小,樣方較多而樹種變化小,為降低因隨機(jī)取樣帶來(lái)的誤差,每次取樣重復(fù)1000次,其結(jié)果平均值為該面積下的物種數(shù)。
2.4 擬合模型
對(duì)數(shù)模型
S=Zln(A) +C
冪函數(shù)模型
S=CAZ
邏輯斯蒂模型
S=B/(C+A-Z)
式中,S為落在樣方內(nèi)的物種數(shù),A為取樣面積,C、B和Z均為模型參數(shù)。
2.5 種-面積關(guān)系擬合優(yōu)度評(píng)價(jià)
采用最小二乘法對(duì)對(duì)數(shù)模型進(jìn)行檢驗(yàn),用高斯牛頓算法(Gauss-Newton)對(duì)邏輯斯蒂模型以及冪函數(shù)模型進(jìn)行檢驗(yàn)。采用赤池信息準(zhǔn)則(AIC)對(duì)3種模型的擬合優(yōu)度進(jìn)行評(píng)價(jià),AIC數(shù)值越小,模型擬合越好,預(yù)測(cè)值越準(zhǔn)確。
本研究中所有數(shù)據(jù)分析、作圖采用R- 3.2.2、Excel 2010完成。
3 結(jié)果分析
3.1 不同尺度植物群落組成
在10、20、30 hm23個(gè)研究尺度下,監(jiān)測(cè)到DBH ≥1的木本植物 15610株、32342 株、49678 株,分別隸屬于19科,29屬,45種;19科,31 屬,47 種;19科,31屬,50種。薔薇科(Rosaceae)在3個(gè)尺度下物種數(shù)最多,其次是槭樹科(Aceraceae)和樺木科(Betulaceae)。而在不同尺度下物種最多的屬為槭屬(Acer),其次為樺木屬(Betula)、忍冬屬(Lonicera)和楊屬(Populus)。單種屬在3個(gè)尺度下分別為17屬、19屬和20屬,占總屬數(shù)的58.62%、55.93%、64.5%。
10、20、30 hm2研究尺度下的胸高斷面積分別為9.97(m2/hm2)、20.45(m2/hm2)、30.99(m2/hm2),10 hm2尺度下胸高斷面積最大的5個(gè)樹種依次為:色木槭(Acermono)、裂葉榆(Ulmuslaciniata),紫椴(Tiliaamurensis)、胡桃楸(Juglansmandshurica)、紅松(Pinuskoraiensis),占總胸高斷面積的57.76%。20 hm2尺度下胸高斷面積最大的5個(gè)樹種依次為:色木槭、裂葉榆、紫椴、紅松、胡桃楸,占總胸高斷面積的55.93%。30 hm2尺度下胸高斷面積最大的5個(gè)樹種依次為裂葉榆、色木槭、紫椴、紅松、楓樺(Betulacostata),占總胸高斷面積的58.08%,是樣地內(nèi)的先鋒樹種。
3.2 不同取樣方式下的種-面積關(guān)系(以30 hm2為例)
3.2.1 巢式取樣的種-面積關(guān)系
在巢式取樣下,根據(jù)模型的AIC值,邏輯斯蒂模型擬合最優(yōu),其次是對(duì)數(shù)模型。冪函數(shù)模型的預(yù)測(cè)值與觀察值之間有一定的差異,在0—10 hm2之間,預(yù)測(cè)物種數(shù)低于觀察值;在15—30 hm2,預(yù)測(cè)值高于觀察值,估計(jì)的預(yù)測(cè)物種數(shù)較高,其擬合效果在3個(gè)模型中最差(AIC分別為85.58,115.20,167.74)(圖2)。依據(jù)種-面積曲線來(lái)看,當(dāng)取樣面積在0—10 hm2之間,隨著取樣面積的逐漸增大,物種數(shù)迅速增加,當(dāng)取樣面積大于10 hm2后,曲線斜率趨于平緩,物種數(shù)隨著取樣面積的增加而緩慢增加。當(dāng)取樣面積為8 hm2時(shí),涵蓋了42個(gè)種,占總物種數(shù)的84%,確定為該群落的最小面積。
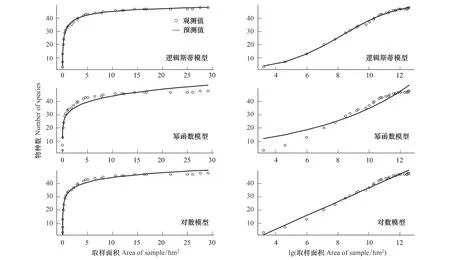
圖2 30 hm2樣地巢式取樣法獲得的3種模型的種-面積曲線Fig.2 Three species-area curves for nest sample design in a 30-hm2 plot
3.2.2 隨機(jī)取樣的種-面積關(guān)系
隨機(jī)取樣下的3個(gè)模型相比較,邏輯斯蒂模型的擬合效果最好,其次是對(duì)數(shù)模型,冪函數(shù)模型的擬合效果最差(AIC分別為76.91、87.40、134.35)。種-面積關(guān)系曲線變化規(guī)律與巢式取樣結(jié)果一致(圖3)。
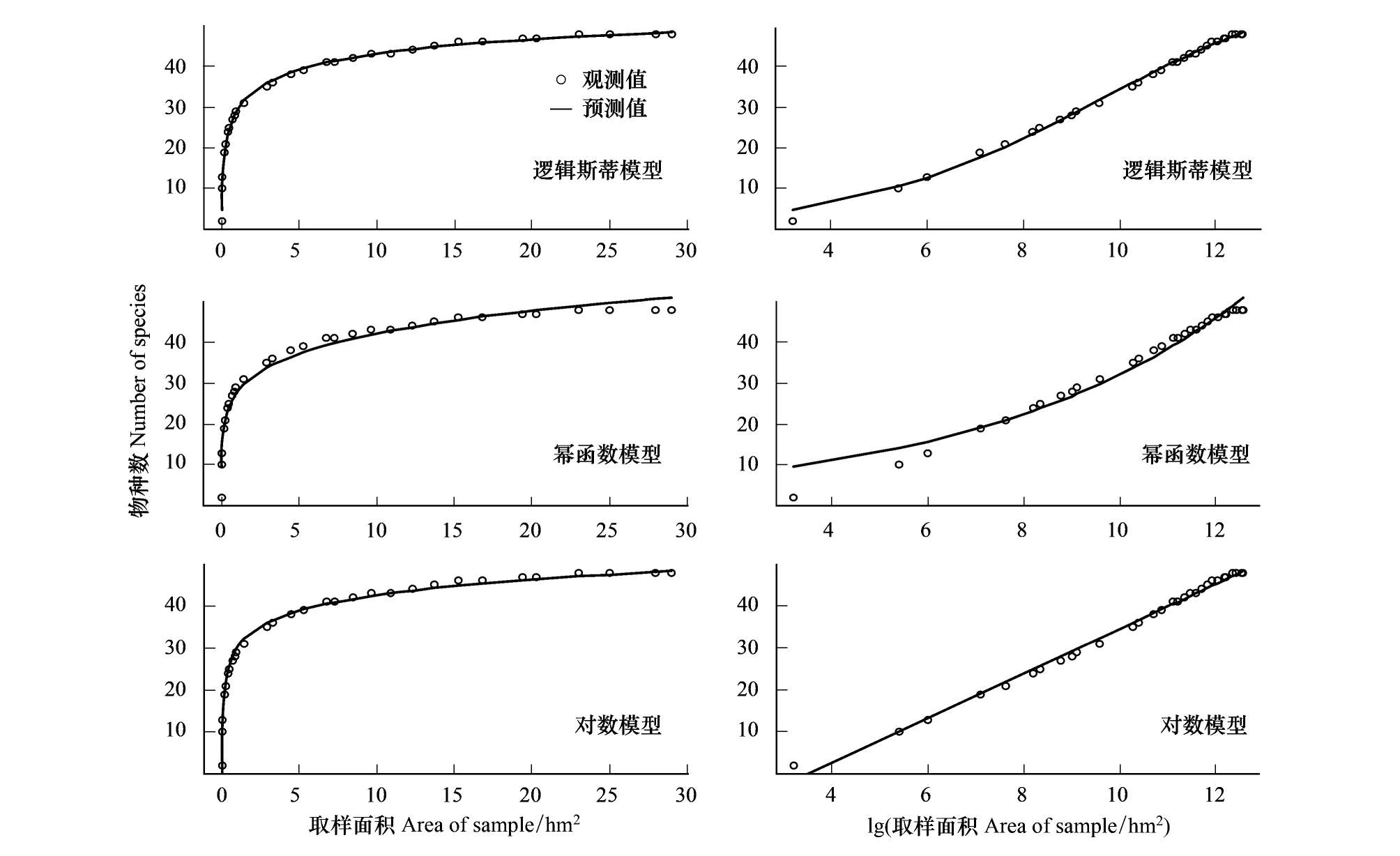
圖3 30 hm2樣地隨機(jī)取樣法獲得的3種模型的種-面積曲線Fig.3 Three species-area curves for random sample design in a 30-hm2 plot
不同取樣方式下的擬合結(jié)果表明,巢式取樣和隨機(jī)取樣下邏輯斯蒂模型的擬合效果都是最優(yōu)的,而冪函數(shù)模型的擬合效果始終相對(duì)較差(表1),可能與冪函數(shù)增長(zhǎng)的趨勢(shì)易高估預(yù)測(cè)物種數(shù)有關(guān)[25]。兩種取樣方法相比,隨機(jī)取樣下的邏輯斯蒂模型可以很好地描述該闊葉紅松林內(nèi)的種-面積關(guān)系。
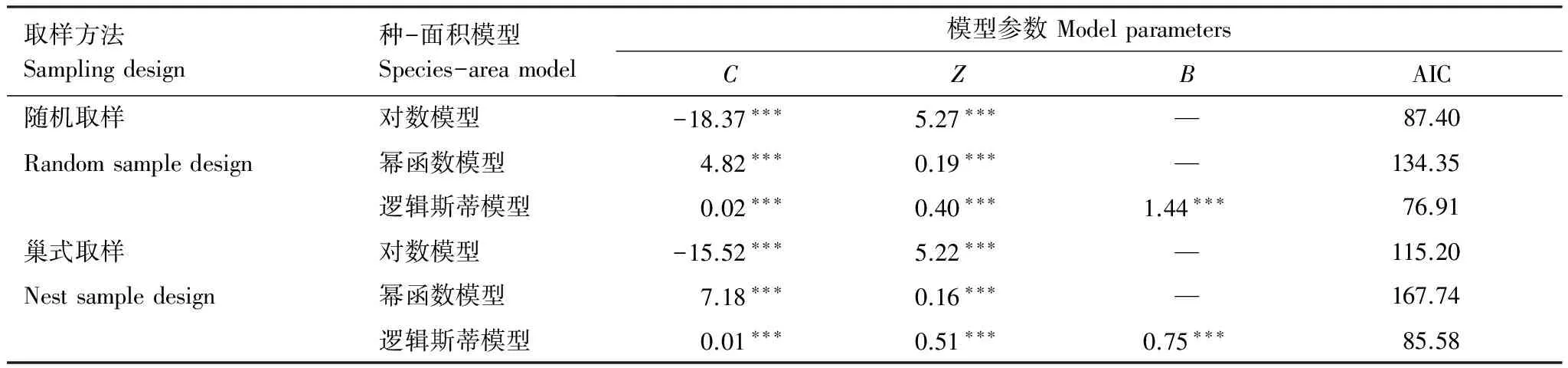
表1 巢式取樣和隨機(jī)取樣下3個(gè)種-面積模型的優(yōu)度檢驗(yàn)
C、Z、B是常數(shù)項(xiàng);***P<0. 001; AIC為赤池信息準(zhǔn)則,值越小代表擬合效果越好
3.3 不同尺度下的種-面積關(guān)系
基于樣地范圍劃分大、中、小3個(gè)尺度范圍,取樣上限分別為30、20、10 hm2。不同尺度下種-面積關(guān)系的結(jié)果表明(表2),在小尺度上,隨機(jī)取樣與巢式取樣下的對(duì)數(shù)模型和邏輯斯蒂模型擬合效果相差不大,均優(yōu)于兩種取樣方法下的冪函數(shù)模型。在中尺度上,以邏輯斯蒂模型最優(yōu),AIC值分別為74.59和79.90。大尺度上,兩種取樣方法下的邏輯斯蒂模型擬合效果優(yōu)于對(duì)數(shù)模型和冪函數(shù)模型,其中隨機(jī)取樣優(yōu)于巢式取樣。
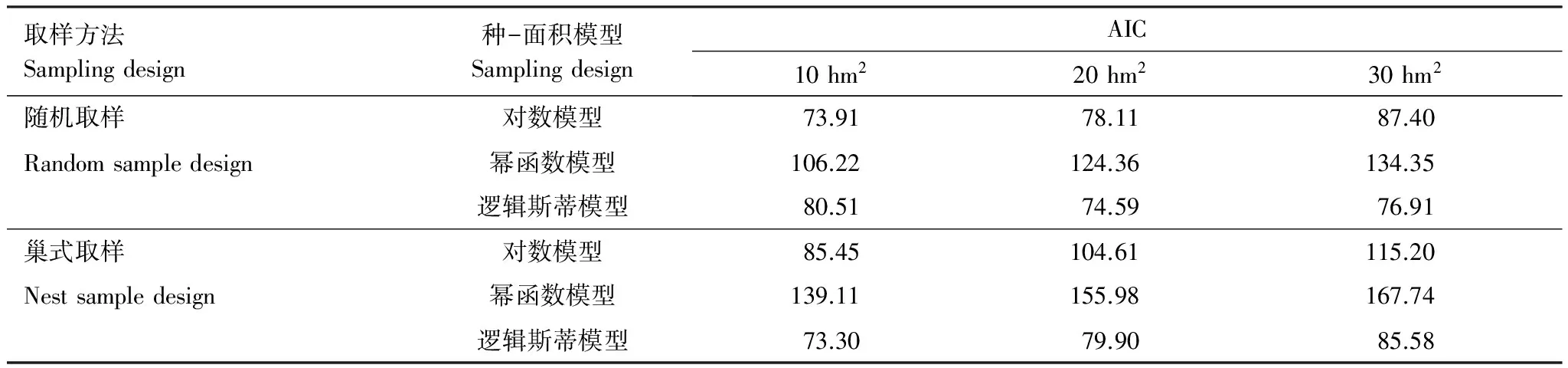
表2 不同尺度下3個(gè)種-面積模型的優(yōu)度檢驗(yàn)
4 討論
近年來(lái),國(guó)內(nèi)外的學(xué)者對(duì)種-面積關(guān)系進(jìn)行了多方面的研究,例如:相對(duì)物種豐富度(RSA)、β多樣性與種-面積關(guān)系的聯(lián)系[26]、種-面積和種-個(gè)體間的關(guān)系[27]等,結(jié)果表明種-面積關(guān)系對(duì)反映地區(qū)生物多樣性規(guī)律有基礎(chǔ)性的意義。該研究基于吉林蛟河30 hm2闊葉紅松林大樣地,分析了作為東北地區(qū)典型植被的種-面積關(guān)系變化規(guī)律,討論了取樣方式和取樣尺度對(duì)蛟河闊葉紅松林種-面積關(guān)系變化的影響。
種-面積曲線(圖2,圖3)確定了吉林蛟河闊葉紅松林的最小面積為8 hm2。物種增加速率隨取樣面積的增加而降低,當(dāng)取樣面積為8 hm2時(shí),出現(xiàn)42個(gè)物種,占總物種數(shù)的84%。而長(zhǎng)白山次生楊樺林的研究表明,在取樣面積為5 hm2時(shí),曲線趨于穩(wěn)定,出現(xiàn)近35個(gè)物種,占總物種數(shù)的80%[28]。比較蛟河樣地和長(zhǎng)白山樣地發(fā)現(xiàn),蛟河森林群落的最小取樣面積大于長(zhǎng)白山森林群落的最小取樣面積,可能是因?yàn)轵院由秩郝湮锓N分布相對(duì)較為分散。
種-面積模型具有尺度效應(yīng),不同尺度下模型的擬合優(yōu)度不同。研究表明,研究區(qū)域的種-面積關(guān)系在大尺度(30 hm2)和中尺度(20 hm2)上適合采用邏輯斯蒂模型,而小尺度(10 hm2)上適合用對(duì)數(shù)模型。在中尺度和大尺度上,邏輯斯蒂模型優(yōu)于對(duì)數(shù)模型和冪函數(shù)模型,為最佳擬合模型,說(shuō)明在取樣范圍足夠大且包括樣地所有物種條件下,邏輯斯蒂模型能夠更好地描述物種和面積的關(guān)系[29]。冪函數(shù)模型擬合優(yōu)度雖然在中尺度和大尺度上低于邏輯斯蒂模型,但模型參數(shù)具有明確的生物學(xué)意義,反映了研究樣地中的α多樣性(C值)(隨機(jī)取樣時(shí)α=4.82;巢式取樣時(shí)α=7.18)和β多樣性(Z值)(隨機(jī)取樣時(shí)β=0.19;巢式取樣時(shí)β=0.16)。本文結(jié)果中,冪函數(shù)模型在不同尺度的擬合均未表現(xiàn)出理想的擬合效果。而在許多其他研究中,小尺度、中尺度上冪函數(shù)模型擬合效果優(yōu)于其他模型[17,30]。這可能是因?yàn)楸狙芯繕拥靥幱跍貛?物種數(shù)相對(duì)較少且聚集程度相對(duì)更低,而冪函數(shù)曲線趨于無(wú)限增長(zhǎng)形式,容易造成預(yù)測(cè)誤差[25]。在小尺度上對(duì)數(shù)模型的擬合效果優(yōu)于其他模型,可能是因?yàn)樾〕叨壬先郝涞沫h(huán)境條件及物種組成相似,新物種數(shù)增加緩慢,更適合用對(duì)數(shù)模型擬合[29- 31]。
種-面積關(guān)系表現(xiàn)出的尺度效應(yīng)與群落物種組成格局密切相關(guān)。Reed認(rèn)為,生境異質(zhì)性影響了物種在不同空間尺度上的分布格局,是影響種-面積關(guān)系的主要推動(dòng)力[32-34]。小尺度上,選取的樣方之間生境差異表現(xiàn)明顯,故物種組成上也會(huì)出現(xiàn)較大的差異,呈現(xiàn)出種-面積曲線的較大變異;而大尺度上,生境異質(zhì)性降低,樣本間差異小,種-面積曲線更為穩(wěn)定[16]。代力民等對(duì)二道白河河岸植物群落及森林群落對(duì)比研究也發(fā)現(xiàn),河岸帶生境異質(zhì)性強(qiáng)、物種豐富度高,二者的種-面積曲線差異較大,表現(xiàn)為河岸帶群落的最小面積小于森林群落[35]。環(huán)境梯度是形成生境異質(zhì)性的主要成分之一[36],闊葉紅松林樣地海拔梯度變化大、坡度變化明顯,群落生境異質(zhì)性高,生境過(guò)濾作用強(qiáng),生態(tài)位相近的物種表現(xiàn)出相似的分布格局[37],物種聚集程度高。隨著研究尺度的增大,出現(xiàn)的新物種數(shù)少,大部分為稀有種或偶見種,這種分布格局與生境異質(zhì)性顯著相關(guān)[38]。
另外,由于研究林分演替階段不同,群落空間格局顯著不同,影響不同尺度上的取樣結(jié)果。姜俊等在蛟河42 hm2針闊混交林的研究也證明了種-面積關(guān)系具有尺度效應(yīng),但與該結(jié)果相異的是小尺度和中尺度以冪函數(shù)模型最優(yōu),在大尺度上以邏輯斯蒂模型最優(yōu),造成兩個(gè)研究結(jié)果差異的原因可能是,處于演替中后期的闊葉紅松林在物種組成以及物種豐富度增加速率上與演替中期的針闊混交林顯著不同[7,39],張春雨等人的研究也表明不同演替階段物種隨取樣面積增加的速率不同,其中中后期的闊葉紅松林的物種增加速率小于中期的針闊混交林[40]。
在蛟河30 hm2闊葉紅松林樣地中,取樣方法對(duì)種-面積關(guān)系影響顯著,其中隨機(jī)取樣優(yōu)于巢式取樣。兩種取樣方法構(gòu)建的種-面積關(guān)系不同,會(huì)影響對(duì)局域物種數(shù)預(yù)測(cè)的準(zhǔn)確性[41]。隨機(jī)取樣時(shí)按設(shè)置樣方的規(guī)格進(jìn)行等概率的抽樣,樣地中各區(qū)域被抽中的概率相等,減少了因生境異質(zhì)性、種子傳播、物種萌生能力強(qiáng)等帶來(lái)的物種聚集分布的情況。與隨機(jī)取樣相比,巢式取樣方法簡(jiǎn)單,但同時(shí)小樣方中的稀有種也出現(xiàn)在大樣方,容易高估物種數(shù),降低預(yù)測(cè)物種數(shù)的準(zhǔn)確性。倪瑞強(qiáng)等對(duì)長(zhǎng)白山森林群落的研究也表明,在取樣方法上隨機(jī)取樣優(yōu)于巢式取樣[42]。Collins等的研究也表明,巢式樣方的種-面積關(guān)系不適合外推估算大尺度的物種數(shù)量[43]。此外,一些研究還表明種-面積關(guān)系也受到取樣面積、樣方數(shù)以及樣方形狀等影響[2,35]。
本研究基于演替階段中后期的闊葉紅松林樣地,認(rèn)為區(qū)域種-面積關(guān)系同時(shí)受研究尺度、取樣方法的影響。在實(shí)際應(yīng)用時(shí),要充分考慮研究區(qū)域的實(shí)際群落結(jié)構(gòu)和樣地內(nèi)環(huán)境差異,在相應(yīng)的尺度上選取合適的取樣方法,從而利用種-面積關(guān)系對(duì)森林經(jīng)營(yíng)給予合理建議。
[1] Lomolino M V. Ecology′s most general, yet protean1pattern: the species-area relationship. Journal of Biogeography, 2000, 27(1): 17- 26.
[2] Palmer M W, White P S. Scale dependence and the species-area relationship. The American Naturalist, 1994, 144(5): 717- 740.
[3] Chave J, Muller-Landau H C, Levin S A. Comparing classical community models: theoretical consequences for patterns of diversity. The American Naturalist, 2002, 159(1): 1- 23.
[4] Zurlini G, Grossi L, Rossi O. Spatial-accumulation pattern and extinction rates of Mediterranean flora as related to species confinement to habitats in preserves and larger areas. Conservation Biology, 2002, 16(4): 948- 963.
[5] Pimm S L, Russell G J, Gittleman J L, Brooks T M. The future of biodiversity. Science, 1995, 269(5222): 347- 350.
[6] Thomas C D, Cameron A, Green R E, Bakkenes M, Beaumont L J, Collingham Y C, Erasmus B F N, de Siqueira M F, Grainger A, Hannah L, Hughes L, Huntley B, van Jaarsveld A S, Midgley G F, Miles L, Ortega-Huerta M A, Peterson A T, Phillips O L, Williams S E. Extinction risk from climate change. Nature, 2004, 427(6970): 145- 148.
[8] 唐志堯, 喬秀娟, 方精云. 生物群落的種-面積關(guān)系. 生物多樣性, 2009, 17(6): 549- 559.
[9] Connor E F, McCoy E D. The statistics and biology of the species-area relationship. The American Naturalist, 1979, 113(6): 791- 833.
[10] Tj?rve E. Shapes and functions of species-area curves: a review of possible models. Journal of Biogeography, 2003, 30(6): 827- 835.
[11] Turner W R, Tj?rve E. Scale-dependence in species-area relationships. Ecography, 2005, 28(6): 721- 730.
[12] Seidler T G, Plotkin J B. Seed dispersal and spatial pattern in tropical trees. PLoS Biology, 2006, 4(11): e344.
[13] 饒米德, 馮剛, 張金龍, 米湘成, 陳建華. 生境過(guò)濾和擴(kuò)散限制作用對(duì)古田山森林物種和系統(tǒng)發(fā)育β多樣性的影響. 科學(xué)通報(bào), 2013, 58(13): 1204- 1212.
[14] 王世雄. 黃土高原子午嶺植物群落物種多樣性的時(shí)空格局與過(guò)程[D]. 西安: 陜西師范大學(xué), 2013.
[15] Williamson M, Gaston K J, Lonsdale W M. The species-area relationship does not have an asymptote. Journal of Biogeography, 2001, 28(7): 827- 830.
[16] 姜俊, 張春雨, 趙秀海. 吉林蛟河42hm2針闊混交林樣地植物種-面積關(guān)系. 植物生態(tài)學(xué)報(bào), 2012, 36(1): 30- 38.
[17] Fisher R A, Corbet A S, Williams C B. The relation between the number of species and the number of individuals in a random sample of an animal population. Journal of Animal Ecology, 1943, 12(1): 42- 58.
[18] 臧婧. 河南寶天曼落葉闊葉林種-面積關(guān)系研究[D]. 哈爾濱: 東北林業(yè)大學(xué), 2015.
[19] Drakare S, Lennon J J, Hillebrand H. The imprint of the geographical, evolutionary and ecological context on species-area relationships. Ecology Letters, 2006, 9(2): 215- 227.
[20] Scheiner S M. Six types of species-area curves. Global Ecology and Biogeography, 2003, 12(6): 441- 447.
[21] Arrhenius O. Species and area. Journal of Ecology, 1921, 9(1): 95- 99.
[22] Gleason H A. Species and area. Ecology, 1925, 6(1): 66- 74.
[23] Ugland K I, Gray J S, Ellingsen K E. The species-accumulation curve and estimation of species richness. Journal of Animal Ecology, 2003, 72(5): 888- 897.
[24] 王雨茜. 吉林蛟河老齡林群落特征及其與地形關(guān)系[D]. 北京: 北京林業(yè)大學(xué), 2013.
[25] Ulrich W, Buszko J. Sampling design and the shape of species-area curves on the regional scale. Acta Oecologica, 2007, 31(1): 54- 59.
[26] Grilli J, Azaele S, Banavar J R, Maritan A. Spatial aggregation and the species-area relationship across scales. Journal of Theoretical Biology, 2012, 313: 87- 97.
[27] Condit R, Hubbell S P, Lafrankie J V, Sukumar R, Manokaran N, Foster R B, Ashton P S. Species-area and species-individual relationships for tropical trees: a comparison of three 50-ha plots. Journal of Ecology, 1996, 84(4): 549- 562.
[28] 郝占慶, 李步杭, 張健, 王緒高, 葉吉, 姚曉琳. 長(zhǎng)白山闊葉紅松林樣地(CBS): 群落組成與結(jié)構(gòu). 植物生態(tài)學(xué)報(bào), 2008, 32(2): 238- 250.
[29] He F L, Legendre P. On species-area relations. The American Naturalist, 1996, 148(4): 719- 737.
[30] Archibald E E A. The specific character of plant communities: II. A quantitative approach. Journal of Ecology, 1949, 37(2): 274- 288.
[31] Williams C B. Area and number of species. Nature, 1943, 152(3853): 264- 267.
[32] Reed T. The number of breeding land bird species on British Islands. Journal of Animal Ecology, 1981, 50(2): 613- 624.
[33] 沈國(guó)春. 生境異質(zhì)性和擴(kuò)散限制對(duì)亞熱帶和熱帶森林物種多樣性維持的作用[D]. 杭州: 浙江大學(xué), 2010.
[34] Guo Y L, Lu J M, Franklin S B, Wang Q G, Xu Y Z, Zhang K H, Bao D C, Qiao X J, Huang H D, Lu Z J, Jiang M X. Spatial distribution of tree species in a species-rich subtropical mountain forest in central China. Canadian Journal of Forest Research, 2013, 43(9): 826- 835.
[35] 代力民, 王青春, 鄧紅兵, 陳高, 王慶禮. 二道白河河岸帶植物群落最小面積與物種豐富度. 應(yīng)用生態(tài)學(xué)報(bào), 2002, 13(6): 641- 645.
[36] Redon M, Bergès L, Cordonnier T. Effects of increasing landscape heterogeneity on local plant species richness: how much is enough? Landscape Ecology, 2014, 29(5): 773- 787.
[37] Debski I, Burslem D F R P, Palmiotto P A, Lafrankie J V, Lee H S, Manokaran N. Habitat preferences of Aporosa in two Malaysian forests: implications for abundance and coexistence. Ecology, 2002, 83(7): 2005- 2018.
[38] Hubbell S P. Tree dispersion, abundance, and diversity in a tropical dry forest. Science, 1979, 203(4387): 1299- 1309.
[39] 劉萬(wàn)德, 蘇建榮, 李帥鋒, 張志鈞, 郎學(xué)東. 南亞熱帶季風(fēng)常綠闊葉林不同演替階段物種-面積關(guān)系. 應(yīng)用生態(tài)學(xué)報(bào), 2011, 22(2): 317- 322.
[40] 張春雨, 趙秀海, 趙亞洲. 長(zhǎng)白山溫帶森林不同演替階段群落結(jié)構(gòu)特征. 植物生態(tài)學(xué)報(bào), 2009, 33(6): 1090- 1100.
[41] Fattorini S. To fit or not to fit? A poorly fitting procedure produces inconsistent results when the species-area relationship is used to locate hotspots. Biodiversity and Conservation, 2007, 16(9): 2531- 2538.
[42] 倪瑞強(qiáng). 長(zhǎng)白山典型針闊混交林群落結(jié)構(gòu)與動(dòng)態(tài)研究[D]. 北京: 北京林業(yè)大學(xué), 2014.
[43] Collins M D, Vázquez D P, Sanders N J. Species-area curves, homogenization and the loss of global diversity. Evolutionary Ecology Research, 2002, 4(3): 457- 464.
Species-area relationships within sample plot in a broad-leaved Korean pine forest at Jiaohe, Jilin Province
CHEN Guilian1,2,TAN Lingzhao1,2, FAN Chunyu1,2, ZHANG Xinna1,2, ZHANG Chunyu1,2, ZHAO Xiuhai1,2,*
1ForestryCollege,BeijingForestryUniversity,Beijing100083,China2KeyLaboratoryforForestResources&EcosystemProcessesofBeijing,BeijingForestryUniversity,Beijing100083,China
The species-area relationship (SAR) is a core component of community ecology, and is an important basis for biological diversity scaling. The SAR is used to describe community types and can solve many ecological problems, such as the determination of minimum sampling areas in a community. Therefore, it is of great importance to diversity conservation. Recently, a number of studies have demonstrated substantial uncertainties in selecting the best SAR model for a data set. In the present study, a 30-hm2permanent forest plot was established in a broad-leaved Korean pine forest in Jiaohe, Jilin Province, China. All trees with diameters at breast height (DBH) ≥ 1 cm were tagged and the height, DBH, and crown diameter of these trees were measured and recorded. We established a logarithmic model, a power function model, and a logistic model using the 30-hm2sample plot to simulate the SAR of a broad-leaved Korean pine forest. We examined how SARs simulated by logarithmic, power function, and logistic models differed after random sampling or nested sampling methods had been used to collect data,and how this difference was affected by sampling scales (broad, moderate, and fine scales). The Akaike Information Criterion (AIC) value was used to compare the goodness-of-fit for each SAR model. The results showed that the sampling method had a significant influence on the SAR, and that the goodness-of-fit for random sampling was better than that for nested sampling at all sampling scales. The establishment of a species-area relationship was closely related to the sampling scales, and the logarithm and logistic models were superior to the power function model at the fine scale (10 hm2). At the moderate and large scales (20 hm2and 30 hm2, respectively), the logistic model better fitted the species-area relationship for broad-leaved Korean pine forest than did the logarithm and the power function models..A comparison of the different models showed that the logistic model with random sampling produced an optimal fit for the species-area relationship within the 30 hm2broad-leaved Korean pine sample area (AIC =76.91), and that the appropriate minimum sampling area was 10 hm2. We concluded that both sampling scale and sampling method had significant influences on the SAR. The scale effect on the SAR is closely related to the community species distribution pattern, and the impacts may result from habitat heterogeneity and successional stage. Habitat heterogeneity and community succession stage might have influenced the number of regional species and species composition, and these different species distribution patterns were reflected in the different SAR curves. Therefore, in practical applications, the variation in the actual community structure and environments within the sampling area should be fully considered. Further work needs to consider the actual situation of the local forest community to simulate the species-area relationship models
species-area relationship; goodness-of-fit; sampling methods; sampling scale
國(guó)家自然科學(xué)基金項(xiàng)目(31670643)
2016- 04- 22; 網(wǎng)絡(luò)出版日期:2017- 03- 02
10.5846/stxb201604220762
*通訊作者Corresponding author.E-mail: zhaoxh@bjfu.edu.cn
陳桂蓮,譚凌照,范春雨,張新娜,張春雨,趙秀海.吉林蛟河闊葉紅松林樣地種-面積關(guān)系.生態(tài)學(xué)報(bào),2017,37(14):4770- 4777.
Chen G L,Tan L Z, Fan C Y, Zhang X N, Zhang C Y, Zhao X H.Species-area relationships within sample plot in a broad-leaved Korean pine forest at Jiaohe, Jilin Province.Acta Ecologica Sinica,2017,37(14):4770- 4777.
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
童話王國(guó)·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(bào)(2020年2期)2020-06-02 11:29:24
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
核科學(xué)與工程(2015年4期)2015-09-26 11:59:03
