Cav1.2鈣通道C末端遠端片段dDCT重組質粒的構建及蛋白制備
2017-10-12 06:48:03胡慧媛雷帥孫德日孫旋旋晏珊劉世浩王健王瑩李越郝麗英
中國醫科大學學報 2017年10期
胡慧媛,雷帥,孫德日,孫旋旋,晏珊,劉世浩,王健,王瑩,李越,郝麗英
(中國醫科大學 1. 藥學院藥物毒理學教研室,沈陽 110122;2. 附屬第四醫院骨科,沈陽 110032)
· 論著 ·
Cav1.2鈣通道C末端遠端片段dDCT重組質粒的構建及蛋白制備
胡慧媛1,雷帥1,孫德日2,孫旋旋1,晏珊1,劉世浩1,王健1,王瑩1,李越1,郝麗英1
(中國醫科大學 1. 藥學院藥物毒理學教研室,沈陽 110122;2. 附屬第四醫院骨科,沈陽 110032)
目的構建Cav1.2鈣通道C末端遠端片段(dDCT)(2 080~2 169)質粒,表達、提取、純化蛋白并進行生物學活性鑒定。方法將dDCT的cDNA片段插入pGEX-6p-1質粒載體并轉化大腸桿菌,異丙基硫代-β-D-半乳糖苷誘導蛋白表達,超聲破碎法提取蛋白。GS-4B beads純化蛋白后,pull-down方法分析其生物學活性。結果構建的dDCT質粒經限制性內切酶和測序雙重鑒定成功,經超聲破碎法提取的dDCT蛋白純度和濃度均較高,并具有能夠與GST-CT1融合蛋白濃度依賴性結合的生物學活性。結論本研究成功構建了dDCT重組質粒,為深入探討Cav1.2鈣通道的自身調節機制奠定了重要的物質基礎。
Cav1.2鈣通道; C末端遠端片段; pull-down方法
Abstract ObjectiveTo construct a recombinant plasmid vector containing the distal fragment of the distal C-terminus( dDCT) of the Cav1.2 channel,and express,extract,and purify dDCT protein and characterize its biological activity.MethodsdDCT cDNA was ligated into the pGEX-6p-1 vector to create a recombinant plasmid that was subsequently transformed into Escherichia coli BL21 competent cells.Expression of GST-dDCT fusion protein from this plasmid was induced with isopropy-β-D-thiogalactoside,and the resulting protein was purified using glutathione-sepharose 4B beads. The biological activity of dDCT was analyzed by GST pull-down assay.ResultsThe recombinant plasmid was verified by restriction enzyme digestion and sequencing. The concentration and purity of the dDCT protein,which was extracted by ultrasonication,were high enough to detect dDCT activity. The binding of dDCT to CT1 was determined to be concentration-dependent.ConclusionThe recombinant dDCT plasmid was successfully constructed,providing the fundamental basis for future studies on mechanisms of Cav1.2 channel autoregulation.
KeywordsCav1.2 Channel; distal fragment of the distal C-terminus; pull-down assay
電壓依賴性L型鈣通道(voltage-dependent L-type calcium channel,VDCC)是可興奮細胞中將電信號轉換成鈣離子內流,進而引起細胞內生物學反應的主要途徑[1]。VDCC是由多個亞單位構成的蛋白復合體,主要包括孔道形成亞單位α1,輔助亞單位β、α2δ和γ。其中α1亞單位決定鈣通道的基礎電生理學特性,且根據其編碼基因的不同可分為7個亞型(Cav1.1~Cav1.4,Cav2.1~Cav2.3)。心臟主要表達Cav1.2亞型[2-3]。心臟的Cav1.2鈣通道主要定位于T管系統,其輔助亞單位Cavβ2b和Cavα2δ-1具有調節鈣通道電壓門控特性和通道轉運的功能[4-5]。
先前的研究表明,Cav1.2鈣通道α1C亞單位遠端C末端部分(distal C-terminus,DCT)易被蛋白水解酶水解,產生截短的Cav1.2鈣通道和水解的C末端片段DCT兩部分。正常的心肌細胞中,大約80%的Cav1.2鈣通道α1C亞單位DCT被切斷,且切斷的DCT仍與鈣通道近端C末端部分(proximal C-terminus,PCT)具有相互作用。DCT可作為Cav1.2鈣通道的自動抑制劑,發揮抑制鈣通道電流的作用[6-7]。研究[8-9]發現,僅表達截短的Cav1.2鈣通道α1C亞單位的轉基因小鼠因心衰在出生前后即死亡,表明Cav1.2鈣通道的DCT片段對心臟的發育和正常功能十分必要。然而,DCT的功能特征以及其對Cav1.2鈣通道的調控機制目前為止尚未闡明。
本研究首先構建Cav1.2鈣通道C末端遠端部分的活性片段(distal fragment of DCT,dDCT)的重組質粒,進而制備純化的dDCT蛋白,同時采用pull-down方法檢測dDCT蛋白與GST-CT1融合蛋白的相互作用,最后鑒定dDCT的生物學活性和功能。dDCT的成功構建,為進一步探討DCT與Cav1.2鈣通道相互作用的功能特征及其分子機制提供了重要的物質基礎,進而為研究與Cav1.2鈣通道相關的心血管疾病的發病機制和防治措施提供了重要的線索。
1 材料與方法
1.1 材料
pGEX-6p-1/dDCT重組質粒由生工生物工程(上海)股份有限公司合成;SanPrep柱式質粒小量抽提試劑盒購自生工生物工程(上海)股份有限公司;BL21(DE3)化學感受態細菌購自北京全式金生物公司;限制性內切酶BamHⅠ和NotⅠ購自美國Thermo Scientific公司;Bradford蛋白定量試劑盒購自碧云天公司;異丙基硫代-β-D-半乳糖苷、氨芐西林、溶菌酶、十二烷基肌氨酸鈉、DTT均購自美國Sigma公司;Precission蛋白酶、谷胱甘肽瓊脂糖凝膠4B(glutathione-sepharose 4B,GS-4B) beads購自美國GE Healthcare公司,其他試劑均購自美國Biosharp公司。
1.2 重組質粒的提取和鑒定
將生工公司提供的表達有pGEX-6p-1/dDCT重組 質 粒 的TOP10細 菌(100 μ L)于5 mL含50 μ g/mL氨芐西林的LB液體培養基中,37 ℃、120 r/min水浴搖床中振搖培養12~16 h,5 000 r/min離心5 min,收集菌液。按照質粒小提試劑盒說明書抽提質粒,測定質粒DNA濃度,分別取1 μ g質粒進行BamHⅠ和NotⅠ的酶切鑒定并送公司進行測序分析。測序結果經PubMed網站的BLAST程序比對同源性。
1.3 重組質粒的轉化
鑒于表達細菌蛋白的需要,按照BL21說明書中的操作步驟,將30 ng重組質粒pGEX-6p-1/dDCT轉化至100 μ L大腸桿菌 BL21中,取轉化后的菌液(75 μ L)均勻涂布于含有 0.1 mg/mL氨芐西林的LB固體瓊脂培養基上,37 ℃恒溫細菌培養箱中培養過夜(約16 h)。挑取經氨芐西林篩選后的單克隆菌株,接種于3 mL含50 μ g/mL氨芐西林的LB液體培養基中,37 ℃、120 r/min水浴搖床中振搖培養約16 h。
1.4 蛋白的提取、純化及鑒定
根據本課題組對Cav1.2鈣通道不同片段蛋白提取方法的總結,本研究選用超聲破碎法提取dDCT蛋白[10-11]。取100 μ L轉化有dDCT質粒并擴增后的BL21菌液,加入到400 mL含50 μ g/mL氨芐西林的LB液體培養基中,振搖過夜。測定菌液的吸光度,待OD600值為0.8~1.0時,加入 1 mmol/L的異丙基硫代-β-D-半乳糖苷誘導融合蛋白表達(37 ℃振搖4 h)。離心收集細菌,然后用含有0.2 mg/mL溶菌酶、10 mmol/L DTT和1.5%十二烷基肌氨酸鈉的Tris緩沖液重懸細菌,冰上放置30 min后,冰上超聲破碎(200 W,超聲3 s停7 s)20 min,結束后加入1% TritonX-100冰上放置30 min。上述裂解液于15 000 g、4 ℃離心10 min后,收集上清。將上清液與GS-4B beads 孵育過夜,結合到beads上的蛋白清洗后,經Precission蛋白酶切除GST。純化后的dDCT蛋白用Bradford蛋白濃度測定試劑盒測定濃度。用上述同樣的方法提取純化GSTCT1融合蛋白,但不進行Precission蛋白酶切步驟,用于后續dDCT蛋白活性檢測。
1.5 dDCT蛋白生物學活性的測定
采用pull-down方法檢測dDCT蛋白與GST-CT1融合蛋白的結合情況。將GST-CT1蛋白與GS-4B beads孵育,使其借助GST附著于beads上。然后將上述beads與純化后不同濃度的dDCT蛋白在1 mmol/L[Ca2+]條件下,4 ℃旋轉儀上共同孵育4 h。將孵育好的樣品重懸于SDS上樣緩沖液中,于15% SDS-PAGE凝膠電泳分離后經考馬斯亮藍染色,拍照、掃描圖片,應用Image J軟件分析蛋白灰度值。
2 結果
2.1 dDCT重組質粒的鑒定結果
dDCT基因片段(287 bp)于pGEX-6p-1質粒載體(4 984 bp)的BamHⅠ/NotⅠ 酶切位點處插入。其中,BamHⅠ酶切位點位于質粒載體的945 bp處,而NotⅠ 酶切位點位于974 bp處。故重組pGEX-6p-1/dDCT質粒全長為5 242 bp,經BamHⅠ/NotⅠ雙酶切后得到2個堿基片段,分別為dDCT片段287 bp和載體片段4 955 bp。如圖1所示,1、2、3泳道分別為完整的dDCT重組質粒,經BamHⅠ單酶切和經BamHⅠ/NotⅠ雙酶切后的質粒片段。不同片段的分子量大小與計算得的理論值相符合。

圖1 重組dDCT質粒的瓊脂糖凝膠電泳圖Fig.1 Agarose gel electrophoresis of recombinant dDCT plasmid
進一步對提取的重組質粒進行測序鑒定分析。在構建重組質粒時,本課題組提供給公司的dDCT堿基序列因GC含量過高,經密碼子優化后,GC含量控制在53.7%,其對應氨基酸序列為AQDPKFIEVTTQE LADACDMTIGEMENAADNILSGGAPQSPNGTLLPFV NC RDPGQDRAGGDEDEGCACALGRGWSEEELADS RVHVRSL,經BLAST比對后與豚鼠Cav1.2鈣通道蛋白(NP_001166394.1)的C末端遠端序列(a.a.2080-2169) 100%同源。
2.2 dDCT蛋白純度和濃度的鑒定結果
經SMS在線DNA和蛋白序列處理工具預測得的dDCT蛋白的表觀分子量是9 520。分別將純化后的GST-dDCT融合蛋白和經Precission蛋白酶切后的純dDCT蛋白進行15% SDS-PAGE電泳,并以純GST蛋白為對照,結果(圖2)顯示:純化后的GST蛋白在其預期分子量26 000處可見特異性條帶,dDCT蛋白和GST-dDCT融合蛋白條帶位置與預期結果一致。dDCT蛋白的雜帶較少,即純度較高;且條帶灰度較高,提示蛋白含量較大。進一步采用Bradford法對dDCT純蛋白進行濃度測定,3次提取蛋白的平均濃度為(0.58±0.10)g/L。經上述實驗鑒定,提取純化的dDCT蛋白純度和濃度均較高,可以用于生物學活性鑒定。
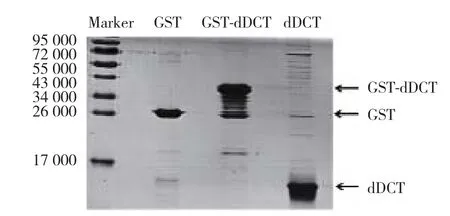
圖2 純化后dDCT蛋白的SDS-PAGE圖Fig.2 SDS-PAGE of purified dDCT protein
2.3 dDCT蛋白與GST-CT1蛋白呈濃度依賴性結合
為了考察經超聲破碎法提取的dDCT蛋白的生物學活性,將純化后不同濃度的dDCT蛋白(0.1,0.3,1,3,10,15 μ mol/L)與GST-CT1融合蛋白在1 mmol/L[Ca2+]條件下共同孵育,經pull-down方法檢測,結果(圖3)顯示:dDCT蛋白能夠與GST-CT1蛋白結合,并且此結合作用具有濃度依賴性。提示經本研究方法制備、純化的dDCT蛋白具有較好的生物學活性,能夠用于后續鈣通道調節機制的相關研究中。
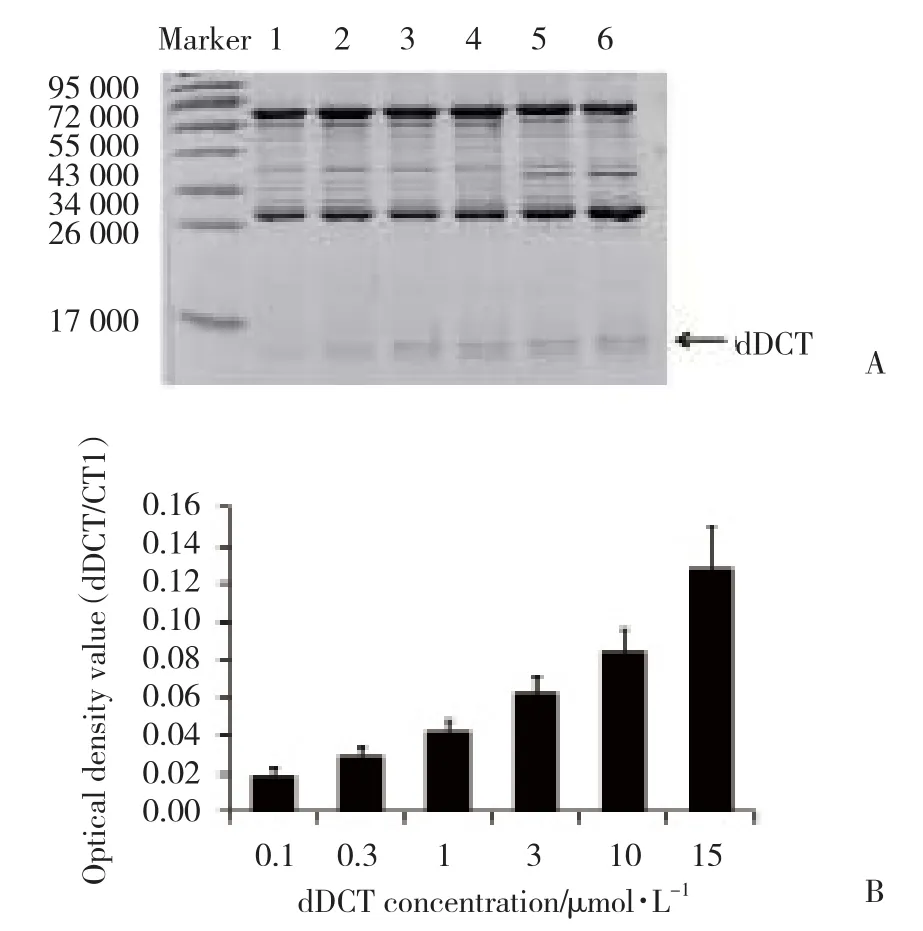
圖3 dDCT蛋白與GST-CT1蛋白呈濃度依賴性結合Fig.3 dDCT binds to GST-CT1 in a concentration-dependent manner
3 討論
隨著人們對Cav1.2鈣通道研究的不斷深入,其自身調控作用越來越受到重視[12]。Cav1.2鈣通道α1C亞單位DCT既可作為Cav1.2鈣通道的自動抑制劑,通過非共價結合的方式抑制Cav1.2鈣通道的電流;亦可作為轉錄因子與NKX2.5和環磷酸腺苷反應元件啟動子直接結合調控基因的表達,參與心肌細胞鈣穩態的維持及干細胞的分化和成熟[13-14]。
Cav1.2鈣通道α1C亞單位的C末端是其在細胞內最大的結構域,約占α1C亞單位總質量的30%。它作為細胞內一系列信號分子打靶和定位的結構支架發揮重要作用,其中包括鈣調蛋白、鈣調蛋白依賴性蛋白激酶Ⅱ、抗藥蛋白、蛋白磷酸酶ⅡA和PKA以及其錨定蛋白AKAP15。先前的研究[6-7]表明,PCT中包含由氨基酸殘基1 694~1 700片段形成短α螺旋結構,它是近端C末端調節域(proximal C-terminal regulatory domain,PCRD)的重要元件,其中R1696和R1697在介導DCT對Cav1.2鈣通道電流的調控中十分必要,但對PCT和DCT之間的結合無影響,而E2103、E2106 和 D2110 是遠端C末端調節域(distal C-terminal regulatory domain,DCRD)的重要調節氨基酸。SHAW等[2]研究發現,DCT能夠與PCT的CB/IQ區域結合,其對Cav1.2鈣電流的抑制作用能夠被Ca2+-CaM拮抗。最近的一項研究[15]表明,在神經元Cav1.3鈣通道C末端介導的抑制中,結構域preIQ-IQ、PCRD和DCRD三者空間上彼此相互靠近是其重要的結構基礎。因而,通過基因重組的方式構建不同的DCT功能片段,對于Cav1.2鈣通道自身調節機制的研究意義重大。
本研究成功構建了Cav1.2鈣通道C末端遠端片段 dDCT (2 080~2 169)質粒,它是 CT3(1 942~2 169)的遠端部分,且包含了部分DCRD結構域。該質粒能夠在大腸桿菌BL21中表達高濃度蛋白,經GS-4B beads純化后,具有較好的純度和生物學活性,能夠用于后續鈣通道調節機制的相關研究中。本研究還發現,dDCT能夠與Cav1.2鈣通道C末端近端片段CT1蛋白濃度依賴性結合,提示dDCT可能參與調控Cav1.2鈣通道的電生理特性以及心肌細胞鈣穩態的維持。而深入探討Cav1.2鈣通道自身調節機制將有助于進一步闡明心血管相關疾病的發生機制和防治藥物的研發。
[1] BODI I,MIKALA G,KOCH SE,et al. The L-type calcium channel in the heart:the beat goes on [J]. J Clin Invest,2005,115(12):3306-3317. DOI:10.1172/JCI27167.
[2] SHAW RM,COLECRAFT HM. L-type calcium channel targeting and local signalling in cardiac myocytes [J]. Cardiovasc Res,2013,98(2):177-186. DOI:10.1093/cvr/cvt021.
[3] HOFMANN F,FLOCKERZI V,KAHL S,et al. L-type CaV1.2 calcium channels:from in vitro findings to in vivo function[J].Physiol Rev,2014,94(1):303-326. DOI:10.1152/physrev.00016.2013.
[4] FANG K,COLERAFT HM. Mechanism of auxiliary β-subunit-mediated membrane targeting of L-type (Ca(V)1.2) channels [J]. J Physiol,2011,589(Pt 18):4437-4455. DOI:10.1113/ jphysiol.2011.214247.
[5] DOLPHIN AC. Voltage-gated calcium channels and their auxiliary subunits:physiology and pathophysiology and pharmacology [J]. J Physiol,2016,594(19):5369-5390. DOI:10.1113/ JP272262.
[6] HULME JT,YAROV-YAROVOY V,LIN TW,et al. Autoinhibitory control of the CaV1.2 channel by its proteolytically processed distal C-terminal domain[J]. J Physiol,2006,576(Pt 1):87-102. DOI:10.1113/jphysiol.2006.111799.
[7] CRUMP SM,ANDRES DA,SIEVERT G,et al. The cardiac L-type calcium channel distal carboxy terminus autoinhibition is regulated by calcium[J]. Am J Physiol Heart Circ Physiol,2013,304(3):H455-H464. DOI:10.1152/ajpheart.00396.2012.
[8] FU Y,WESTNBROEK RE,YU FH,et al. Deletion of the distal C terminus of CaV1.2 channels leads to loss of beta-adrenergic regulation and heart failure in vivo[J]. J Biol Chem,2011,286(14):12617-12626. DOI:10.1074/jbc.M110.175307.
[9] DOMES K,DING J,LEMKE T,et al. Truncation of murine CaV1.2 at Asp-1904 results in heart failure after birth [J]. J Biol Chem,2011,286(39):33863-33871. DOI:10.1074/jbc.M111.252312.
[10] SHAO D,ZHAO M,XU J. The individual N- and C-lobes of calmodulin tether to the Cav1.2 channel and rescue the channel activity from run-down in ventricular myocytes of guinea-pig heart[J]. FEBS Lett,2014,588(21):3855-3861. DOI:10.1016/j.febslet.2014.09.029.
[11] 何桂林,邵冬雪,印丹丹,等. 體外重組CaV1.2不同蛋白片段純化及其與CaM相互作用的研究[J]. 中國醫科大學學報,2013,42(9):773-776.
[12] SATIN J,SCHRODER EA. Auto-regulation of cardiac L-type calcium channels[J]. Trends Cardiovasc Med,2009,19(8):268-271.DOI:10.1016/j.tcm.2010.02.009.
[13] BANNISTER JP,LEO MD,NARAYANAN D,et al. The voltage-dependent L-type Ca2+(CaV1.2) channel C-terminus fragment is a bi-modal vasodilator[J]. J Physiol,2013,591(12):2987-2998.DOI:10.1113/jphysiol.2013.251926.
[14] GAO Q,GE J,JU Y,et al. Roles of L-type calcium channels (CaV1.2)and the distal C-terminus (DCT) in differentiation and mineralization of rat dental apical papilla stem cells (rSCAPs) [J]. Arch Oral Biol,2017,74:75-81. DOI:10.1016/j.archoralbio.2016.11.004.
[15] LIU N,YANG Y,GE L,et al. Cooperative and acute inhibition by multiple C-terminal motifs of L-type Ca2+channels [J]. Elife,2017,6.pii:e21989. DOI:10.7554/eLife.21989.
(編輯 陳 姜)
Construction of and Protein Preparation from a Recombinant Plasmid Containing the Distal Fragment of the Distal C-terminus of the Cav1.2 Channel
HU Huiyuan1,LEI Shuai1,SUN Deri2,SUN Xuanxuan1,YAN Shan1,LIU Shihao1,WANG Jian1,WANG Ying1,LI Yue1,HAO Liying1
(1. Department of Pharmaceutical Toxicology,School of Pharmacy,China Medical University,Shenyang 110122,China;2. Department of Orthopedics,The Fourth Affiliated Hospital,China Medical University,Shenyang 110032,China)
R96
A
0258-4646(2017)10-0865-04
http://kns.cnki.net/kcms/detail/21.1227.R.20170927.0942.002.html
10.12007/j.issn.0258‐4646.2017.10.001
國家自然科學基金(81100108,31471091);醫學電生理學教育部重點實驗室開放基金(201605);遼寧省大學生創新創業訓練項目(201410159018)
胡慧媛(1978 -),女,副教授,博士.
郝麗英,E-mail:lyhao@cmu.edu.cn
2017-03-28
網絡出版時間:2017-09-27 09:42
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
現代畜牧科技(2021年4期)2021-07-21 06:13:00
科技傳播(2019年22期)2020-01-14 03:06:54
福建基礎教育研究(2019年1期)2019-09-10 07:22:44
福建基礎教育研究(2019年1期)2019-05-28 08:39:49
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
數學物理學報(2017年2期)2017-06-05 09:12:30
