蒙古沙冬青根系DSE定殖與土壤因子典范對應分析
2017-10-19 09:16:32王少杰侯力峰解琳琳胡從從賀學禮
河北大學學報(自然科學版) 2017年5期
王少杰,侯力峰,解琳琳,胡從從,賀學禮
(河北大學 生命科學學院,河北 保定 071002)
蒙古沙冬青根系DSE定殖與土壤因子典范對應分析
王少杰,侯力峰,解琳琳,胡從從,賀學禮
(河北大學 生命科學學院,河北 保定 071002)
為進一步明確不同荒漠地區蒙古沙冬青(Ammopiptanthusmongolicus)根系深色有隔內生真菌(DSE)生態分布與土壤環境之間的關系,分別于2015年7月從烏海、磴口、阿拉善左旗、民勤和沙坡頭5個樣地采集蒙古沙冬青根圍0~10、10~20、20~30、30~40和40~50 cm共5個土層土壤樣品,運用典范對應分析(CCA)方法系統研究了DSE生態分布及其與土壤因子的相關性.結果表明,蒙古沙冬青根系DSE分布與環境密切相關,其分布在不同樣地間差異顯著,定殖率表現為阿拉善左旗>烏海>沙坡頭>民勤>磴口.5個樣地DSE平均定殖率為52.02%,說明蒙古沙冬青與DSE能形成良好根-菌共生關系.不同樣地土壤性質差異顯著,對DSE分布具有重要影響.主要表現為同一樣地不同土層,DSE分布與土壤理化指標均表現出淺土層大于深土層的趨勢.CCA結果表明,DSE菌絲、微菌核、定殖率和定殖強度在CCA排序軸上得到較好分化.CCA二維排序圖表明,烏海樣地影響DSE菌絲定殖的主要土壤因子為酸性磷酸酶(ACP)、堿性磷酸酶(ALP)和速效P;磴口樣地主要為堿解N和有機C;阿拉善左旗樣地主要為ALP和pH;民勤樣地主要為pH、ALP和土壤脲酶(Urease);沙坡頭樣地主要為ACP、Urease和堿解N.研究表明,不同地區DSE分布受不同土壤因子影響,這可為荒漠地區土壤狀況評估以及改善提供依據.
深色有隔內生真菌(DSE);定殖率;土壤因子;典范對應分析
蒙古沙冬青(Ammopiptanthusmongolicus)屬第三紀孑遺物種,是國家保護漸危種,主要分布在干旱地區,是西北荒漠獨有的旱生常綠豆科闊葉灌木[1].沙冬青的抗逆能力極強,在高鹽度、強紫外輻射和極端溫度等環境條件下具有較強的適應性[2].馬彥軍等[3]通過對沙冬青種子萌發期抗逆性研究,證明沙冬青種子在萌發期及幼苗期具有較強的耐旱、耐鹽性.Wu等[4]利用Illumina技術和DEGs分析比較了蒙古沙冬青種子在干旱和低溫環境下的基因圖譜,得到完整的沙冬青耐旱耐寒基因轉錄圖譜,進一步表明沙冬青的耐旱、耐寒性.
深色有隔內生真菌(dark septate endophytes,DSE)泛指一類定居在植物根表皮、皮層和維管組織細胞內或細胞間隙的內生真菌,具有深色有隔菌絲和微菌核等結構[5-6].DSE廣泛分布在沿海灘涂、內陸高原山地和熱帶、溫帶、凍原及南北極地區[7].據調查,DSE宿主涵蓋114科320屬600余種植物[5],除菌根植物外,在莎草科、十字花科、藜科等傳統非菌根植物根系中也有定殖[7].DSE細胞壁中產生的黑色素能夠增強宿主植物在重金屬污染區[8]、干旱地區[9]以及低pH土壤[10]等脅迫環境中的抗逆性;分泌的多種胞外水解酶,能夠保證其對各種形態營養元素的利用[11].研究發現,蒙古沙冬青根部皮層組織有DSE定殖,并形成菌絲網絡、胞間菌絲和微菌核等結構,其定殖率與土壤理化性質具有顯著相關性[12].目前,關于蒙古沙冬青DSE空間分布與土壤因子的相關性時有報道[13-14],但尚未對不同地區影響蒙古沙冬青DSE分布的主要土壤因子進行系統研究.
本實驗通過對西北不同荒漠樣帶蒙古沙冬青根圍土壤樣品采集和分析,基于典范對應分析(CCA)來探討不同樣地蒙古沙冬青DSE定殖規律與土壤因子的交互關聯,旨在闡明不同荒漠環境影響DSE定殖和生態分布的主要土壤因子,為充分利用微生物資源和荒漠生態改良提供依據.
1 材料和方法
1.1 樣品采集
實驗樣地為內蒙古烏海、磴口、阿拉善左旗,甘肅民勤和寧夏沙坡頭(表1).2015年7月在各樣地隨機選取4株長勢良好的蒙古沙冬青植株,在距植株主干0~30 cm范圍內挖土壤剖面,分別在0~10、10~20、20~30、30~40和40~50 cm 5個土層采集土樣和根樣,編號裝入自封袋中,記錄采樣時間、地點、土壤類型等,帶回實驗室.自然風干后,過2 mm篩分離根樣和土樣.土樣用于土壤理化性質測定,將根樣剪成1 cm長根段,用于DSE形態結構和定殖率觀測.
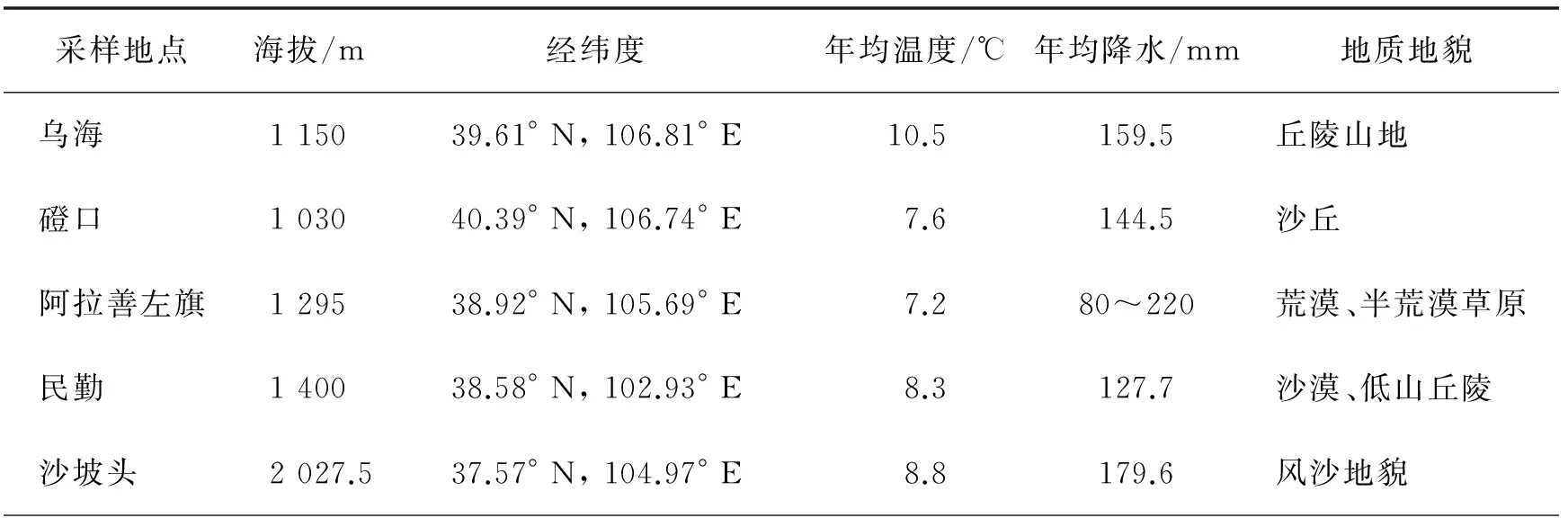
表1 樣地概況Tab.1 Sampling sites and overview
1.2 研究方法
根據Phillips和Hayman方法[15]對DSE進行染色,然后按Biermann和Linderman的方法[16]測定DSE定殖率.隨機選取30條1 cm長根段用100 g/L KOH處理,酸性品紅染色后,脫色液脫色,在光學顯微鏡下觀察DSE定殖結構,并統計DSE定殖率和定殖強度.


根據Li等[12]研究,本實驗選取pH、速效P、有機C、堿解N、酸性磷酸酶(ACP)、堿性磷酸酶(ALP)、土壤脲酶(Urease)等土壤因子作為測量指標.其中土壤pH值用電位法測定,用碳酸氫鈉-鉬銻抗比色法測定,堿解N用堿解擴散法測定,用灼燒質量法測定土壤有機C.ACP和ALP用改進的Tabatabai和Brimner方法[17]測定,活性分別以每g土樣培養1 h ACP和ALP轉化對硝基苯磷酸二鈉(pNPP)的量(μg·g-1)表示; Urease用改進的Hoffmann和Tabatabai 比色法[18]測定,活性以每g土樣培養1 h催化尿素分解產生NH3-N的μg表示.
1.3 數據處理
數據經 Excel 整理后,采用 SPSS20 統計軟件進行單因子方差( One-Way ANOVA)分析和Pearson 法兩兩相關分析,用Canoco 4.5軟件進行典范性分析(canonical correspondence analysis,CCA)[19].
2 結果與分析
2.1 DSE定殖結構
觀察結果表明,DSE能夠侵入到蒙古沙冬青根組織,形成菌絲和微菌核結構.菌絲多為深色,具橫隔,能侵入到宿主根系表皮和維管組織(圖1a-e).菌絲可在皮層細胞間隙延伸并形成菌絲網(圖1c),有時可延伸到皮層細胞內形成胞內菌絲(圖1d),偶見橫隔距離縮短的特化菌絲(圖1e).微菌核是由皮層細胞或維管組織(圖1f-l)內膨大、加厚、變圓的菌絲堆積而成,根據其形態可分為聚合型和離散型,其中聚合型多為腦狀(圖1g-h)、啞鈴狀(圖1i);離散型多為帶狀(圖1j)和鏈狀(圖1k),有時呈星點狀(圖1l).
2.2 DSE空間分布
由表2可知,沙冬青根圍DSE分布與生態環境密切相關.同一樣地,DSE各項指標在不同土層間無顯著差異,除烏海外,其他樣地菌絲定殖率和微菌核定殖率最大值均在0~20 cm土層.不同樣地,烏海和阿拉善左旗菌絲定殖率顯著大于其他樣地,民勤最低;微菌核定殖率在不同樣地差異顯著,表現為沙坡頭>民勤>烏海>阿拉善左旗>磴口;總定殖率表現為阿拉善左旗>烏海>沙坡頭>民勤>磴口;阿拉善左旗、民勤和沙坡頭定殖強度顯著大于烏海和磴口.
同列數據后不同小寫字母表示同一樣地不同土層間在P<0.05 水平差異顯著,不同大寫字母表示不同樣地同一土層在P<0.05水平差異顯著;同列均值后不同大寫字母表示不同樣地在P<0.05水平上差異顯著.
2.3 土壤因子空間分布
由表3知,土壤pH在不同樣地差異顯著,沙坡頭顯著大于其他樣地,但同一樣地,不同土層間無顯著差異.不同樣地,民勤ACP和ALP活性最大,磴口最小;各樣地0~10 cm土層ACP活性最大;烏海和阿拉善左旗樣地0~10 cm土層ALP活性最大,并隨土層加深而降低,磴口和民勤樣地0~20 cm土層ALP活性顯著大于20~50 cm土層,沙坡頭樣地ALP活性最大值在30~40 cm土層.烏海、磴口和阿拉善左旗樣地Urease活性顯著大于民勤和沙坡頭;烏海和民勤樣地Urease活性最大值在0~10 cm土層,并隨土層加深而降低;磴口和阿拉善左旗樣地Urease活性最大值在30~40 cm土層,沙坡頭樣地在10~20 cm土層,3樣地最小值均在40~50 cm土層.
速效P、有機C和堿解N含量變化與樣地和土層深度有密切關系.速效P、有機C和堿解N最大值均在民勤樣地.烏海不同土層間速效P無顯著差異,其他樣地最大值均在0~10 cm土層.烏海、民勤、阿拉善左旗和沙坡頭樣地有機C最大值均在0~10 cm土層,磴口最大值在20~30 cm土層.烏海、阿拉善左旗和民勤樣地0~10 cm土層堿解N顯著大于其他土層,并隨土層加深而降低,沙坡頭最大值在10~20 cm土層,磴口樣地土層間堿解N無顯著差異.
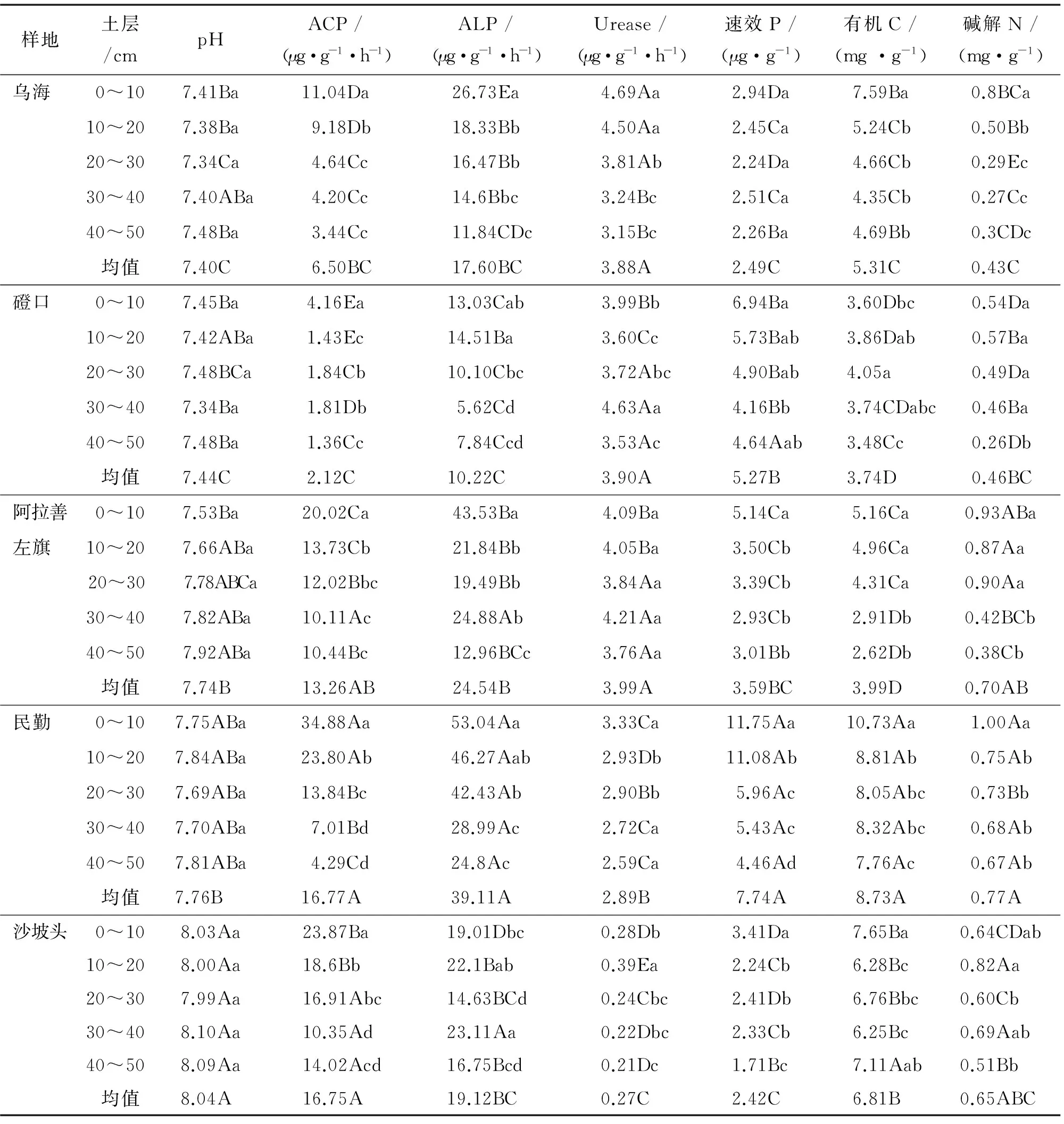
表3 蒙古沙冬青根圍土壤因子空間分布Tab.3 Spatial distribution of soil factors in the rhizosphere of Ammopiptanthus mongolicus
同列數據后不同小寫字母表示同一樣地不同土層間在P<0.05 水平差異顯著,不同大寫字母表示不同樣地同一土層在P<0.05水平差異顯著;同列均值后不同大寫字母表示不同樣地在P<0.05水平上差異顯著.
2.4 DSE不同定殖指標的相關性
由表4可知,DSE菌絲定殖率與微菌核定殖率呈負相關,與總定殖率呈極顯著正相關;DSE微菌核定殖率與定殖強度呈極顯著正相關;總定殖率與定殖強度顯著正相關.
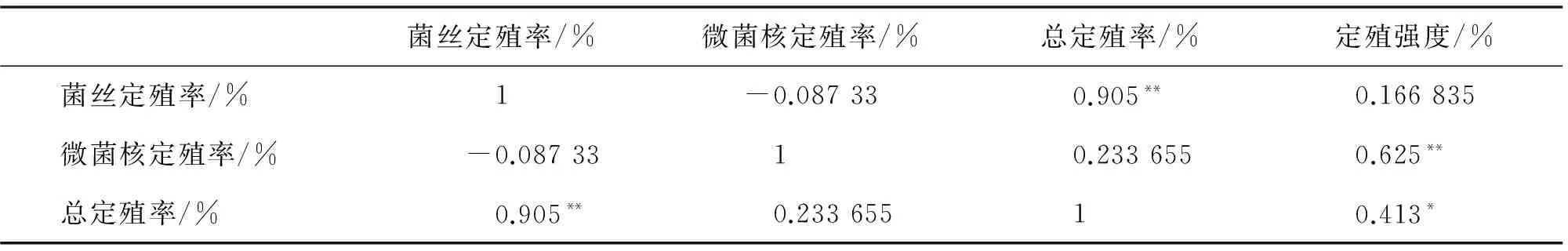
表4 DSE不同定殖指標的相關性分析Tab.4 Correlation analysis of different colonization indexes of DSE
* 表示兩者之間在P<0.05 水平(雙側)上顯著相關;**表示兩者之間在P<0.01水平上極顯著相關.
2.5 DSE分布與土壤因子的CCA
由表5可知,CCA 排序中前2個軸累積解釋了物種-環境關系的90%以上,說明CCA前兩軸已能較好地反映DSE分布與土壤因子的關系.圖2為各樣地DSE分布與土壤因子的CCA排序圖,通過排序圖可直觀看出不同土壤因子(箭頭表示)之間以及DSE分布(三角符號表示)與土壤因子之間的關系[20].
由表5可知,烏海樣地土壤因子與第1排序軸相關性最高的是pH,其次是ACP、ALP和有機C;磴口樣地最高為速效P,其次為有機C、ACP和ALP;阿拉善左旗樣地最高為pH,其次為Urease和ALP;民勤樣地最高為pH,其次是堿解N、ALP和Urease;沙坡頭樣地最高為速效P,其次是ACP、Urease和堿解N.烏海樣地土壤因子與第2排序軸相關性最高的土壤因子為ACP,其次是速效P、ACP、Urease和堿解N;磴口樣地最高為pH,其次為Urease和堿解N;阿拉善左旗樣地最高為ACP,其次為pH;民勤樣地最高為堿解N,其次是ALP、Urease和有機C;沙坡頭樣地最高為pH,其次是Urease、堿解N和速效P.
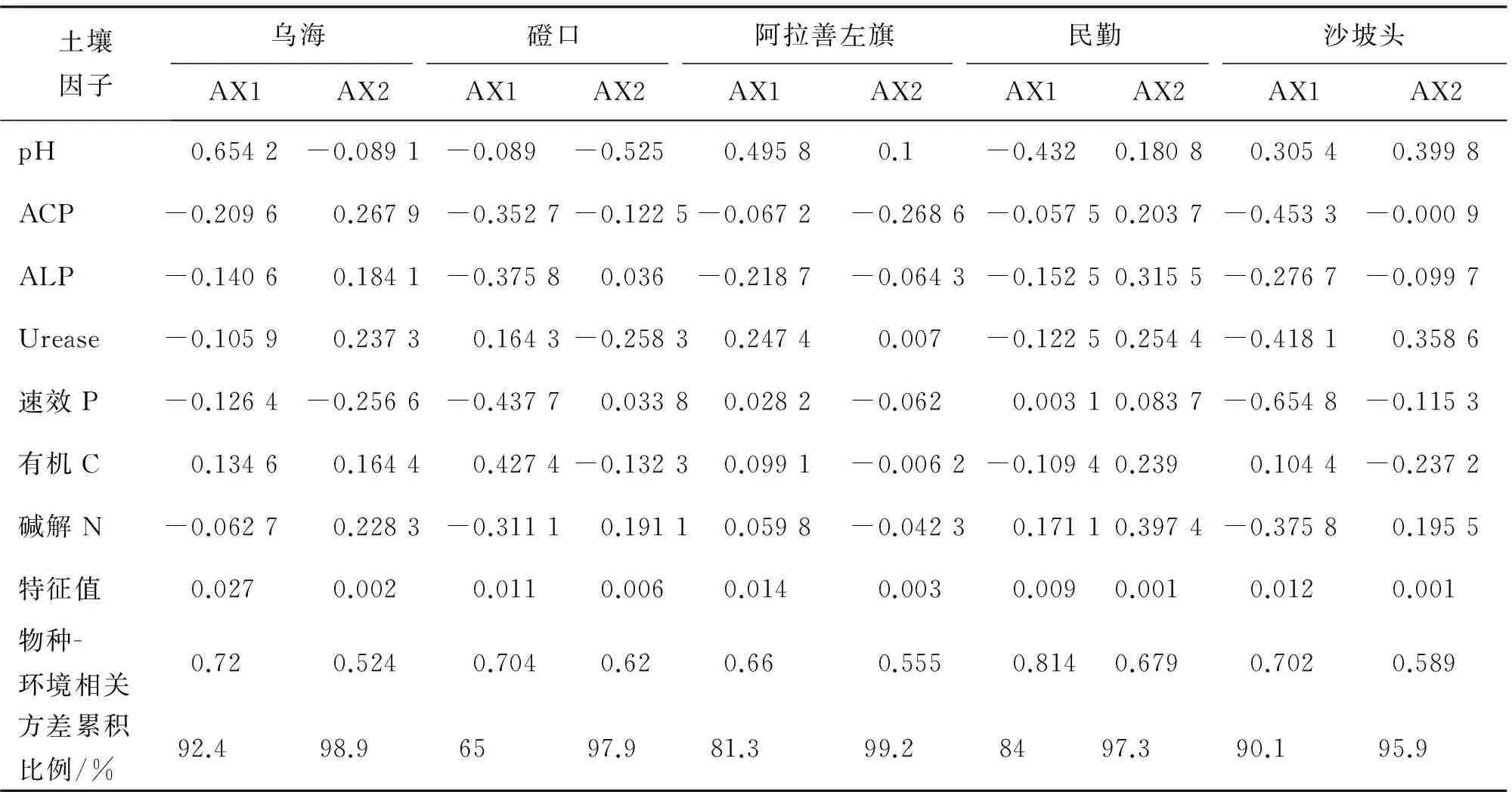
表5 土壤因子與CCA排序軸的相關關系及排序概要Tab.5 Correlation coefficients between soil factors and CCA ordination axes and and summary
AX1為第1排序軸,AX2為第2排序軸.
CCA排序圖中,研究對象(三角符號)與箭頭越接近,表明受到土壤因子的影響越大,兩者的相關性越高[20].由圖2可知,烏海DSE菌絲定殖率與總定殖率與ACP、ALP和速效P呈正相關,微菌核定殖率主要受pH的影響.磴口DSE菌絲定殖率與堿解N和有機C具有正相關,微菌核受ACP、ALP和速效P影響.阿拉善左旗DSE菌絲定殖率與ALP和pH的相關性較大,微菌核主要受Urease和有機C影響.民勤菌絲主要受pH、ALP和Urease的影響,微菌核與堿解N具有相關性.沙坡頭菌絲定殖率主要受Urease、堿解N和ACP影響,微菌核主要受速效P、ALP和有機C影響.
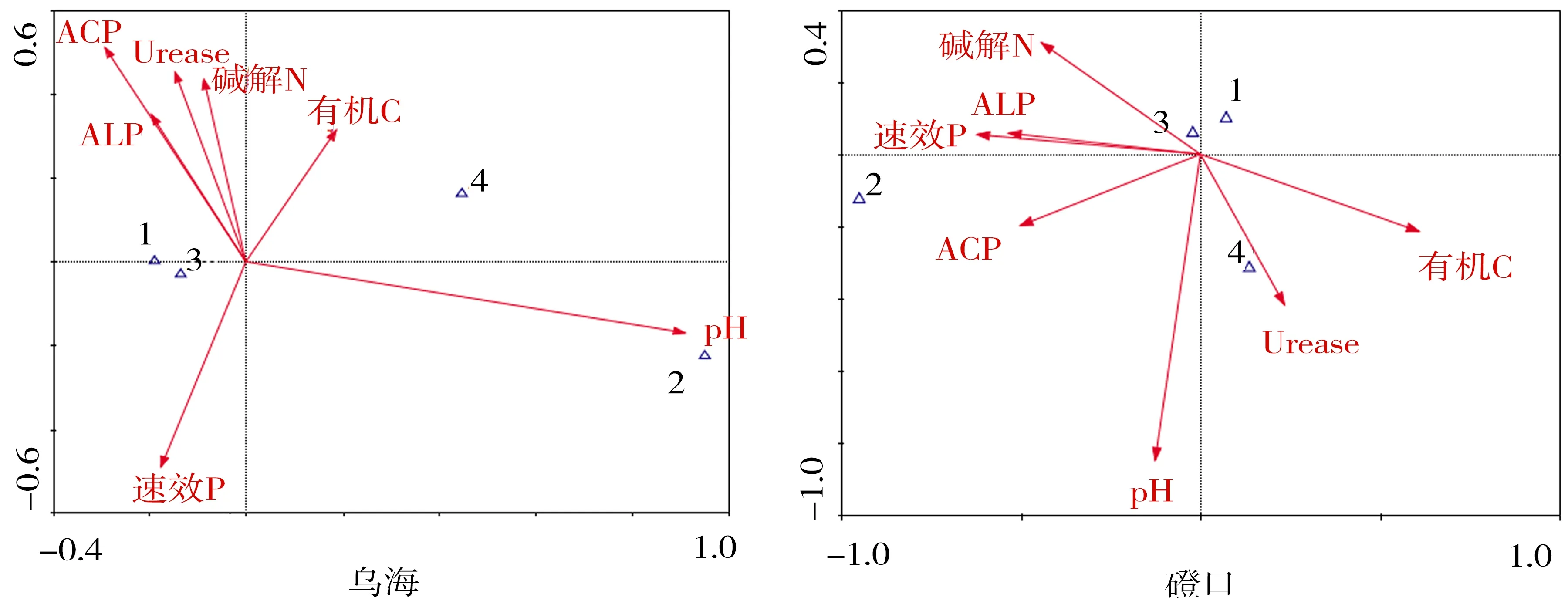
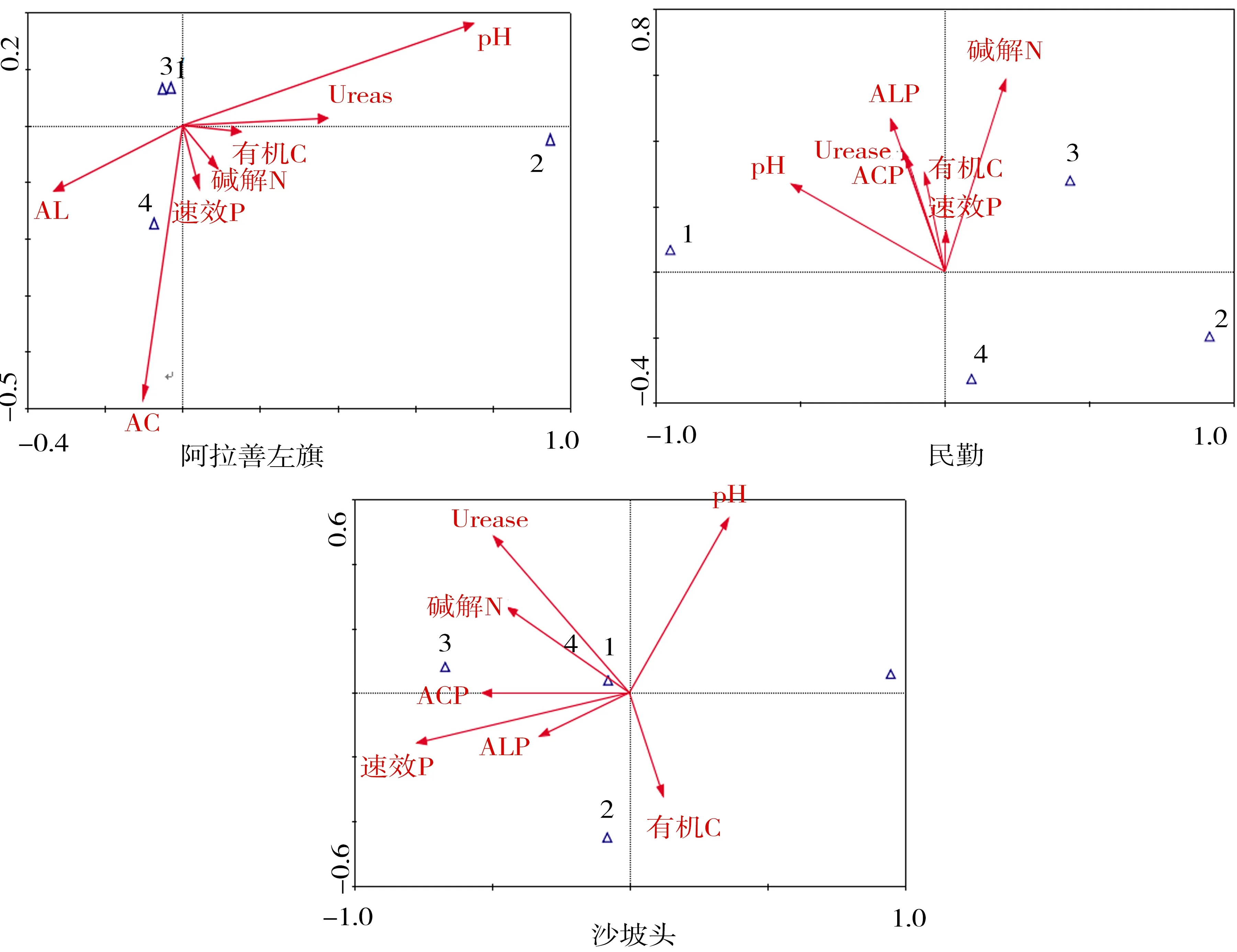
1.菌絲定殖率;2.微菌核定殖率;3.總定殖率;4.定殖強度.圖2 各樣地DSE定殖與土壤因子的CCA排序Fig.2 CCA ordination biplot between DSE and soil factors in all samples
3 討論
實驗結果表明,5個樣地總定殖率為43.33%~58.22%,平均總定殖率為52.02%,這與前人研究結果一致[21].說明DSE是蒙古沙冬青根部重要組成.從實驗結果來看,荒漠區植物根系DSE定殖強度(31.29%)和重金屬礦區植物根系(32.8%)基本一致,但荒漠區植物根系DSE微菌核具有多種形態,且定殖率(10.62%)明顯高于重金屬礦區(2.4%)[22].說明DSE在不同生態系統均有廣泛分布,相對于重金屬礦區,荒漠環境更利于微菌核分布.
本研究中,DSE菌絲定殖率與微菌核定殖率表現出明顯的空間異質性,且菌絲定殖率與微菌核定殖率呈負相關關系.菌絲是DSE與植物進行營養物質交換的通道[23], DSE黏質菌絲可幫助宿主植物在干旱環境中維持水分和營養的運輸[24].微菌核是DSE菌絲膨大加厚堆積而成,有時與菌絲相連[8],可作為DSE的繁殖結構[14],因此菌絲和微菌核可能存在一種相互轉化關系.當宿主植物與DSE互動聯系降低時,為應對不利環境,菌絲轉變成微菌核,此外微菌核內還含有多種酶,能夠幫助荒漠植物分解土壤中的營養物質[21];反之微菌核則重新萌發出菌絲.
研究表明,土壤理化性質對植被分布、微生物群落的組成和功能具有重要影響[25-26],其中土壤有機C是土壤碳庫的重要組成,對土壤特性、根系深度和有效水分保持能力等土壤學特性具有重要影響,其含量的高低可直接影響土壤肥力[27].賀學禮等[28]研究表明,土壤有機C、堿解N和ALP對土壤細菌、真菌和放線菌數量具有顯著影響.實驗結果表明磴口有機C含量顯著低于其他樣地,土壤肥力相應偏低,植物不能為DSE生長提供足夠的養分,直接影響DSE菌絲和微菌核分布.沙坡頭和民勤樣地微菌核定殖率和磷酸酶活性均顯著大于其他樣地,土壤磷酸酶主要來自土壤微生物和植物根系分泌,能夠分解土壤有機磷[29],增加土壤P含量.DSE微菌核與其他土壤微生物能夠協同作用,可通過分泌多種酶來改善植物根部微環境,增加土壤養分,促進植物營養吸收.此外,沙坡頭和民勤樣地有機C和堿解N含量也很豐富,植物可直接從土壤中吸收養分,DSE菌絲對宿主植物的促生作用降低,DSE菌絲分布減少.阿拉善左旗樣地菌絲定殖率高,可增加根系營養和水分吸收面積,促進植物生長,反過來植物又為DSE生長提供養分,因此,DSE菌-根共生結構可能是旱生植物應對干旱環境的有效對策之一.
本實驗中,除了烏海樣地,其他樣地DSE菌絲定殖率和微菌核定殖率最大值都出現在0~20 cm土層,并隨土層加深而有減少趨勢.土壤酶活性和有機C、速效P和堿解N等土壤因子也表現出淺土層大于深土層的趨勢.這可能因為土壤表層落葉和腐殖質積累較多,土壤通氣狀況好,微生物種類豐富,有利于DSE生長.Stone等[30]研究表明隨著土壤深度增加,土壤理化性質和土壤酶活性也隨之發生變化.Eilers等[31]指出隨著土壤深度增加,碳氮含量以及土壤微生物量和相對豐度也隨之降低.CCA結果表明,DSE定殖率與土壤因子呈正相關,這一對應關系也說明DSE的生長與土壤養分密切相關.在一定養分范圍內,隨著土壤養分的增加,可供蒙古沙冬青吸收的養分也隨之增加,為DSE菌絲萌發和生長提供養料,從而提高蒙古沙東青DSE定殖率.
CCA能夠直接反映環境因子對排序結果的影響[32],相對于常規統計學方法而言,其所包含的信息量更大,結果更加直觀[20].近年來,CCA排序法已廣泛應用于物種分布與環境因子關系的研究[33],在土壤-環境領域和土壤性質與環境因子間定量關系方面也取得了重要進展[34].本實驗CCA排序圖結果表明,5個樣地影響DSE分布的主要土壤因子各不相同.研究表明,土壤生物碳儲量[35]和侵蝕程度[36]對土壤理化性質和微生物分布具有重要影響.本實驗5個樣地位于西北不同荒漠地帶,其氣候特征、地質狀況以及土壤侵蝕情況(表1)、主要植物種群分布均具有較大差異[13,37],使得不同樣地影響DSE分布的主要土壤因子存在差異.Li[12]、孫茜[13]和張淑容[14]等人通過主成分分析法已經證實西北荒漠區蒙古沙冬青DSE生態分布與堿解N、土壤酶和球囊霉素等土壤因子具有顯著相關性,與本實驗結果相比具有一定差異.這可能因為前者是對西北整個荒漠地區DSE生態分布與土壤因子的相關性進行分析,而忽略了不同樣地之間的差異.因此,本實驗在前人研究的基礎上,利用典范對應分析法對每個地區DSE分布與土壤因子的相關性進行分析,并篩選出每個樣地影響DSE分布的主要土壤因子,用于評估5個樣地土壤營養狀況,并為土壤肥力改善提供支持.
綜上,植物共生真菌、宿主植物和環境因子是一個相互聯系、相互制約的動態平衡體系.通過檢測荒漠地區植被DSE定殖情況及其與土壤因子之間的關系,可評估荒漠土壤環境狀況,進一步認識荒漠地區DSE 的生態作用和意義,為荒漠土壤環境改善和充分利用土壤真菌資源,促進荒漠環境植被恢復提供理論基礎.
[1] 尹林克,潘伯榮,趙振東,等.沙冬青屬植物引種試驗研究[J].干旱區研究,1988(4): 36-43.DOI:10.13866/j.azr.1988.04.007.
[2] 王燁,尹林克.兩種沙冬青耐鹽性測定[J].干旱區研究,1991,8(2): 20-22.DOI:10.13866/j.azr.1991.02.005.
[3] 馬彥軍,段慧榮,曹致中,等.沙冬青種子萌發期抗逆性研究[J].中國沙漠,2011,31(4): 963-967.
MA Y J,DUAN H R,CAO Z Z,et al.Stress resistance ofAmmopiptanthusmongolicusseeds during germination period[J].Journal of Desert Research,2011,31(4): 963-967.
[4] WU Y,WEI W,PANG X,et al.Comparative transcriptome profiling of a desert evergreen shrub,Ammopiptanthusmongolicus,in response to drought and cold stresses[J].BMC Genomics,2014,15(1): 671.DOI:10.1186/1471-2164-15-671.
[5] JUMPPONEN A,TRAPPE J M.Dark septate endophytes: a review of facultative biotrophic root-colonizing fungi[J].New Phytologist,1998,140(2): 295-310.DOI:10.1046/j.1469-8137.1998.00265.x.
[6] BARROW J,AALTONEN R.Evaluation of the internal colonization ofAtriplexcanescens(Pursh)Nutt.roots by dark septate fungi and the influence of host physiological activity[J].Mycorrhiza,2001,11(4):199-205.DOI 10.1007/s005720100111
[7] 劉茂軍,張興濤,趙之偉.深色有隔內生真菌(DSE)研究進展[J].菌物學報,2009,28(6): 888-894.DOI:10.13346/j.mycosystema.2009.06.009.
LIU M J,ZHANG X T,ZHAO Z W.Advances in the research of dark septate endophytes[J].Mycosystema,2009,28(6): 888-894.DOI:10.13346/j.mycosystema.2009.06.009.
[8] ZHAO D,LI T,SHEN M,et al.Diverse strategies conferring extreme cadmium (Cd)tolerance in the dark septate endophyte (DSE),Exophialapisciphila: evidence from RNA-seq data[J].Microbiol Res,2015,170: 27-35.DOI:10.1016/j.micres.2014.09.005.
[9] KNAPP D G,PINTYE A,KOVáCS G M.The dark side is not fastidious-dark septate endophytic fungi of native and invasive plants of semiarid sandy areas[J].PLoS ONE,2012,7(2): e32570.DOI:10.1371/journal.pone.0032570.
[10] POSTMA J W M,OLSSON P A,FALKENGREN-GRERUP U.Root colonisation by arbuscular mycorrhizal,fine endophytic and dark septate fungi across a pH gradient in acid beech forests[J].Soil Biology and Biochemistry,2007,39(2): 400-408.DOI:10.1016/j.soilbio.2006.08.007.
[11] MANDYAM K,JUMPPONEN A.Seeking the elusive function of the root-colonising dark septate endophytic fungi[J].Studies in Mycology,2005,53: 173-189.DOI:10.3114/sim.53.1.173.
[12] LI B,HE X,HE C,et al.Spatial dynamics of dark septate endophytes and soil factors in the rhizosphere ofAmmopiptanthusmongolicusin Inner Mongolia,China[J].Symbiosis,2015,65(2): 75-84.DOI:10.1007/s13199-015-0322-6.
[13] 張淑容,賀學禮,徐浩博,等.蒙古沙冬青根圍 AM 和 DSE 真菌與土壤因子的相關性研究[J].西北植物學報,2013,33(9): 1891-1897.
ZHANG S R,HE X L,XU H B,et al.Correlation study of AM and DSE fungi and soil factors in the rhizosphere ofAmmopiptanthusmongolicus[J].Acta Botanica Boreali-Occidentalia Sinica,2013,33(9): 1891-1897.
[14] 孫茜,賀超,賀學禮,等.沙冬青與伴生植物深色有隔內生真菌定殖規律及其與土壤因子的相關性[J].植物生態學報,2015,39(9): 878-889.DOI:10.17521/cjpe.2015.0084.
SUN Q,HE C,HE X L,et al.Colonization of dark septate endophytes in roots ofAmmopiptanthusmongolicusand its associated plants as influenced by soil properties[J].Chinese Journal of Plant Ecology,2015,39(9): 878-889.DOI:10.17521/cjpe.2015.0084.
[15] PHILLIPS J M,HAYMAN D S.Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection[J].Transactions of the British Mycological Society,1970,55(1): 158-118.DOI:10.1016/s0007-1536(70)80110-3.
[16] BIERMANN B,LINDERMAN R G.Quantifying vesicular-arbuscular mycorrhizae: a proposed method towards standardization[J].New Phytologist,1981,87(1): 63-67.DOI:10.1111/j.1469-8137.1981.tb0190.x.
[17] 宋勇春,李曉林,馮固.菌根真菌磷酸酶活性對紅三葉草生境中土壤有機磷虧缺的影響[J].生態學報,2001,21(7): 1130-1135.DOI:10.3321/j.issn:1000-0933.2001.07.014.
SONG Y C,LI X L,FENG G.Effect of phosphatase activity on soil organic phosphorus loss in the environment of clover growth[J].Acta Ecologica Sinica,2001,21(7):1130-1135.DOI:10.332/j.issn:1000-0933.2001.07.014.
[18] 周禮愷.土壤酶學[M].北京:科學出版社,1987.
[19] 魏玉蓮,姬蘭柱,王淼,等.長白山北坡靜水水體中水甲蟲分布與環境關系的典范對應分析[J].應用生態學報,2002,13(1): 91-94.DOI :10.13287/j .1001 -9332.2002.0020.
WEI Y L,JI L Z,WANG M,et al.CCA of water beetles' distribution and environmental factors in lentic samples of North Changbai Mountain.[J].Chinese Journal of Applied Ecology,2002,13(1): 91-94.DOI :10.13287/j .1001 -9332.2002.0020.
[20] 楊思存,逄煥成,王成寶,等.基于典范對應分析的甘肅引黃灌區土壤鹽漬化特征研究[J].中國農業科學,2014,47(1): 100-110.DOI:10.3864/j.issn.0578-1752.2014.01.011.
YANG S C,PANG H C,WANG C B,et al.Characterization of soil salinization based on canonical correspondence analysis method in Gansu Yellow River Irrigation District of Northwest China[J].Scientia Agricultura Sinica,2014,47(1): 100-110.DOI:10.3864/j.issn.0578-1752.2014.01.011.
[21] 姜橋,賀學禮,陳偉燕,等.新疆沙冬青 AM 和 DSE 真菌的空間分布[J].生態學報,2014,34(11): 2929-2937.DOI: 10.5846 / stxb201305121025.
JIANG Q,HE X L,CHEN W Y,et al.Spatial distribution of AM and DSE fungi in the rhizosphere ofAmmopiptanthusnanus[J].Acta Ecologica Sinica,2014,34(11): 2929-2937.DOI: 10.5846 / stxb201305121025.
[22] XU R,LI T,CUI H,et al.Diversity and characterization of Cd-tolerant dark septate endophytes (DSEs)associated with the roots of Nepal alder (Alnusnepalensis)in a metal mine tailing of southwest China[J].Applied Soil Ecology,2015,93: 11-18.DOI:10.1016/j.apsoil.2015.03.013.
[23] PETERSON R L,WAGG C,PAUTLER M.Associations between microfungal endophytes and roots: do structural features indicate function[J].Botany,2008,86(5): 445-456.DOI:10.1139/b08-016.
[24] BARROW J R.Atypical morphology of dark septate fungal root endophytes ofBoutelouain arid southwestern USA rangelands[J].Mycorrhiza,2003,13(5): 239-247.DOI:10.1007/s00572-003-0222-0.
[25] 陳少鋒,韓新輝,馮永忠,等.陜北黃土丘陵區土壤碳氮庫對人工植被恢復的響應[J].河北大學學報(自然科學版),2014,34(05):508-515.DOI:10.3969/j.issn.1000-1565.2014.05.012.
CHEN S F,HAN X H,FENG Y Z,et al.Response of soil organic carbon and total nitrogen pool to different types of artificial vegetations in the gully region of Loess Plateau[J].Journal of Hebei University(Natural Science Edition),2014,34(05):508-515.DOI:10.3969/j.issn.1000-1565.2014.05.012.
[26] 尚占環,丁玲玲,龍瑞軍,等.江河源區退化高寒草地土壤微生物與地上植被及土壤環境的關系[J].草業學報,2007,16(1): 34-40.DOI:10.3321/j.issn:1004-5759.2007.01.006.
SHANG Z H,DING L L,LONG R J,et al.Relationship between soil microorganisms,above-ground vegetation,and soil environment of degraded alpine meadows in the headwater areas of the yangtze and yellow rivers,qinghai-tibetan plateau[J].Acta Prataculturae Sinica,2007,16(1): 34-40.DOI:10.3321/j.issn:1004-5759.2007.01.006.
[27] 徐薇薇,喬木.干旱區土壤有機碳含量與土壤理化性質相關分析[J].中國沙漠,2014,34(6): 1558-1561.DOI:10.7522/j.issn.1000-694X.2013.00311.
XU W W,QIAO M.Soil carbon contents in relation to soil physicochemical properties in arid regions of china[J].Journal of Desert Research,2014,34(6): 1558-1561.DOI:10.7522/j.issn.1000-694X.2013.00311.
[28] 賀學禮,李寶庫,陳嚴嚴,等.蒙古沙冬青根圍土壤微生物分布及與土壤因子相關性研究[J].干旱區資源與環境,2016,30(4): 51-55.DOI:10.13448/j.cnki.jalre.2016.112.
HE X L,LI B K,CHEN Y Y,et al.Correlations between soil microbial distribution and soil factors in the rhizosphere ofAmmopiptanthusmongolicus[J].Journal of Arid Land Resources and Environment,2016,30(4): 51-55.DOI:10.13448/j.cnki.jalre.2016.112.
[29] 張愛民,李乃康,趙鋼勇,等.土壤中解磷、解鉀微生物研究進展[J].河北大學學報(自然科學版),2015,35(4):442-448.DOI:10.3969/j.issn.1000-1565.2015.04.019.
ZHANG A M,LI N K,ZHAO G Y,et al.Research progress on the phosphate-solubilizing and potassium-solubilizing microorganisms[J].Journal of Hebei University(Natural Science Edition),2015,35(4):442-448.DOI:10.3969/j.issn.1000-1565.2015.04.019.
[30] STONE M M,DEFOREST J L,PLANTE A F.Changes in extracellular enzyme activity and microbial community structure with soil depth at the Luquillo Critical Zone Observatory[J].Soil Biology and Biochemistry,2014,75: 237-247.DOI:10.1016/j.soilbio.2014.04.017.
[31] EILERS K G,DEBENPORT S,ANDERSON S,et al.Digging deeper to find unique microbial communities: The strong effect of depth on the structure of bacterial and archaeal communities in soil[J].Soil Biology and Biochemistry,2012,50: 58-65.DOI:10.1016/j.soilbio.2012.03.011.
[32] LEP J,MILAUER P.Multivariate analysis of ecological data using CANOCO[M].Cambridge: Cambridge University Press,2003.
[33] BROWN N D,CURTIS T,ADAMS E C.Effects of clear-felling versus gradual removal of conifer trees on the survival of understorey plants during the restoration of ancient woodlands[J].Forest Ecology and Management,2015,348: 15-22.DOI:10.1016/j.foreco.2015.03.030.
[34] JIN Z,LI Z,LI Q,et al.Canonical correspondence analysis of soil heavy metal pollution,microflora and enzyme activities in the Pb-Zn mine tailing dam collapse area of Sidi village,SW China[J].Environmental Earth Sciences,2014,73(1): 267-274.DOI:10.1007/s12665-014-3421-4.
[35] HERATH H M S K,CAMPS-ARBESTAIN M,HEDLEY M.Effect of biochar on soil physical properties in two contrasting soils: an alfisol and an andisol[J].Geoderma,2013,209: 188-197.DOI:10.1016/j.geoderma.2013.06.016.
[36] 胡嬋娟,劉國華,郭雷,等.土壤侵蝕對土壤理化性質及土壤微生物的影響[J].干旱區研究,2014,31(4): 702-708.DOI:10.13866/j.azr.2014.04.17.
HU C J,LIU G H,GUO L,et al.Effects of soil erosion on soil physicochemical properties and soil microorganisms[J].Arid Zone Research,2014,31(4): 702-708.DOI:10.13866/j.azr.2014.04.17.
[37] 郭清華,胡從從,賀學禮,等.蒙古沙冬青伴生植物AM真菌的空間分布[J].生態學報,2016,36(18):5809-5819.DOI: 10.5846 /stxb201503300609.
GUO Q H,HU C C,HE X L,et al.Exploration of the spatial distribution of AM fungi in the rhizospheres ofAmmopiptanthusmongolicus-associated plants[J].Acta Ecologica Sinica,2016,36(18): 5809-5819.DOI: 10.5846 /stxb201503300609.
(責任編輯:趙藏賞)
CCAanalysisofcolonizationofDSEintherhizosphereofAmmopiptanthusmongolicusinrelationtosoilfactors
WANGShaojie,HOULifeng,XIELinlin,HUCongcong,HEXueli
(College of Life Sciences,Hebei University,Baoding 071002,China)
Ammopiptanthusmongolicusis the only evergreen broadleaf shrub grown in the northwest China that has the characteristics of cold and drought resistance,salt resistance and wind resistance.It plays an important role in windbreak and soil and water conservation.To elucidate the spatial distribution of dark septate endophytes (DSE)and the relationship of DSE with soil factors associated with the desert plantA.mongolicusbased on canonical correspondence analysis (CCA),the author sampled from five different sites in northwest China in July 2015,which are Wuhai,Dengkou,Alxa,Minqin and Shapotou,respectively.Root samples were collected from four robustA.mongolicus,and soil samples were taken from the rhizosphere ofA.mongolicusin five sites and divided into five depth intervals,0-10 cm,10-20 cm,20-30 cm,30-40 cm and 40-50 cm,respectively.The results showed that the roots ofA.mongolicuscould be infected highly by DSE and formed a typical type of septate hyphae and microsclerotia.The distribution of DSE was significantly affected by ecological conditions.There were significant differences in distribution of DSE among the five sites.The colonization rates of DSE among the five sites exhibited a pattern as Alxa>Wuhai>Shapotou>Minqin>Dengkou.The average colonization rate of five sites was 52.02%,which showed thatA.mongolicusand DSE could form a symbiotic relationship.There were also significant differences in soil factors among different sites.In the same site,DSE distribution and soil indicators had the maximum values in the profile of a surface layer and showed the trend of decreasing with the depth of the profile.The results of CCA showed that DSE hyphae,microsclerotia,colonization rate and colonization intensity in the axis of CCA got better differentiation.The species-environment biplots were drawn based on the result of CCA.The relationships between the colonization of DSE and the seven soil factors were clearly revealed on these biplots.Accoding to the biplots,the main soil factors of five sites were different.The acid phosphatase (ACP),alkaline phosphatase (ALP)and available phosphate were found to be main soil factors affecting the hypha colonization of DSE in Wuhai.The main soil factors of Dengkou were available nitrogen and organic carbon.The hypha colonization of DSE in Alxa were mainly affected by ALP and pH.The main soil factors affecting the hypha colonization of DSE were pH,ALP and Urease,in Minqin.And the main soil factors affecting the hypha colonization of DSE in Shapotou were ACP,Urease and available nitrogen.The research results showed that the distribution of DSE in different regions was affected by different soil factors,which could provide the basis for the evaluation and improvement of soil conditions in the desert area.
dark septate endophytes (DSE);colonization rate;soil factors;canonical correspondence analysis
Q939.96
A
1000-1565(2017)05-0497-11
10.3969/j.issn.1000-1565.2017.05.009
2016-11-20
國家自然科學基金資助項目(31170488)
王少杰(1988—),男,河南漯河人,河北大學碩士研究生,主要從事菌根生物學研究.E-mail: wsj663@163.com
賀學禮(1963—),男,陜西蒲城人,河北大學教授,博士生導師,主要從事菌根生物學與生態學研究.E-mail: xuelh1256@aliyun.com
