多巴胺通過mTOR-EAAT2通路影響星形膠質細胞谷氨酸攝取能力*
2017-10-20 05:22:58溫芳芳劉樂平楊建靜丁賽丹
中國病理生理雜志 2017年10期
溫芳芳, 徐 竹, 劉樂平, 楊建靜, 丁賽丹
(溫州醫科大學附屬第一醫院,浙江省神經老化與疾病研究重點實驗室, 浙江 溫州 325000)
多巴胺通過mTOR-EAAT2通路影響星形膠質細胞谷氨酸攝取能力*
溫芳芳, 徐 竹, 劉樂平, 楊建靜, 丁賽丹△
(溫州醫科大學附屬第一醫院,浙江省神經老化與疾病研究重點實驗室, 浙江 溫州 325000)
目的研究多巴胺(dopamine,DA)對星形膠質細胞谷氨酸(glutamate,Glu)攝取能力的影響,以及DA通過哺乳動物雷帕霉素靶蛋白(mTOR)-興奮性氨基酸轉運體2(EAAT2)信號通路對星形膠質細胞Glu攝取能力的影響。方法采用Amplex Red谷氨酸測定試劑盒檢測經過干預的原代皮層星形膠質細胞對Glu攝取含量的變化,RT-qPCR、Western blot和免疫熒光染色等檢測EAAT2和mTOR mRNA 和蛋白質相對表達量,mTOR拮抗劑雷帕霉素或mTOR興奮劑MHY1485干預在DA中共培養的星形膠質細胞,檢測mTOR和EAAT2的表達情況,以及培養上清液Glu的含量。結果DA干預的原代星形膠質細胞中mTOR表達下調,EAAT2表達下調,培養上清液Glu水平上升;雷帕霉素干預后,EAAT2表達下調,培養上清液中Glu的含量增加;MHY1485干預后,EAAT2表達上調,培養上清液中Glu的含量下降。結論DA通過與星形膠質細胞mTOR-EAAT2通路相互作用,減弱星形膠質細胞攝取Glu的能力,引起細胞外Glu蓄積,最終損傷星形膠質細胞的功能。
多巴胺; 谷氨酸; 星形膠質細胞
多巴胺(dopamine,DA)是α、β腎上腺素受體和多巴胺受體的激動劑。谷氨酸(glutamate,Glu)是大腦內主要的興奮性神經遞質,在學習和記憶上有著重要作用。Glu是哺乳動物的中樞神經系統中主要的興奮性神經遞質,但是過度刺激Glu受體反而會造成興奮性中毒[1]。Glu中毒已知參與很多神經系統疾病包括阿爾茨海默癥,缺血性腦卒,帕金森綜合征,癲癇癥和抑郁癥[2-4]。
活化的星形膠質細胞既具有保護神經元的作用,又能分泌細胞毒因子、炎癥因子和補體蛋白而損害神經元[5]。我們前期實驗證實DA蓄積會誘導兒茶酚氧位甲基轉移酶(catechol-O-methyltransferase,COMT)抑制劑表達上調[6]。COMT是多巴胺(dopamine,DA)的降解酶,本課題組前期實驗中采用DA干預,發現星形膠質細胞分泌TNF-α并導致神經元凋亡[7],后又發現DA抑制神經元中學習記憶通路Glu-NO-cGMP的活性[8],因此推測腦組織DA含量增加可能導致細胞外Glu累積。突觸間隙的興奮性氨基酸轉運體2(excitatory amino acid transporter 2, EAAT2)能清除谷氨酸,哺乳動物類雷帕霉素靶蛋白(mammalian target of rapamycin, mTOR)被報道可以調控Glu轉運體表達[3]。Glu誘導的興奮性中毒可能在DA干預的星形膠質細胞功能損傷中扮演著重要角色。本研究探索DA是否會減弱星形膠質細胞對Glu的攝取能力,闡明DA和信號通路協同損傷星形膠質細胞的分子機制。
材 料 和 方 法
1藥物、儀器與試劑
微量滲析探針和微量泵購自BAS;mTOR拮抗劑雷帕霉素、mTOR興奮劑MHY1485、辣根過氧化物酶、 L-谷氨酸氧化酶、 谷氨酸丙酮酸氧基轉移酶、丙氨酸和6-羧基熒光素購自Sigma-Aldrich; Amplex Red谷氨酸測定試劑盒、RNA-Easy試劑盒、低聚糖(dT)、dNTP、 DTT、莫羅尼鼠白血病病毒逆轉錄酶、RNaseOUT和5×FS緩沖液購自Invitrogen;EAAT2和mTOR引物由Invitrogen合成。ABI-棱柱7700序列檢測系統購自Applied Biosystems。iTaqTM超高速與ROX購自Bio-Rad。
2實驗方法
2.1原代皮層星形膠質細胞(primary cortical astrocytes,PCAs)的分離和干預[9]取新生24 h內的SD乳鼠,大腦皮層組織用機械消化得到細胞懸液。細胞鋪板在75 cm2的細胞培養瓶,將1.5×106個細胞培養在1 mL含1% 血清的DMEM/F12培養基中,培養72 h。之后,PCAs分別以每孔1.5×106培養在6孔培養板中。mTOR拮抗劑雷帕霉素(100 μmol/L)或mTOR興奮劑MHY1485(10 μmol/L)與DA(最終濃度為10 μmol/L)共培養星形膠質細胞,對照加同樣量的緩沖液,3個重復。
2.2細胞外Glu攝取含量檢測 通過Amplex Red谷氨酸測定試劑盒檢測經過干預后PCAs對Glu攝取含量的變化。在DA(10 μmol/L)干預PCAs 12 h之后,由含500 μmol/L谷氨酸的HEPES緩沖液替代,在每隔相同時間,取50 μL上清液轉入96孔板中,接著與50 μL混合物(100 mmol/L Amplex Red,250 U/L辣根過氧化物酶,80 U/L L-谷氨酸氧化酶,500 U/L谷氨酸丙酮酸氧基轉移酶和200 μL丙氨酸)混合后,37℃溫浴 30 min。酶標儀在波長為530 nm處(對比參考波長為590 nm)測定吸光度。谷氨酸含量代入谷氨酸標準曲線計算得到。
2.3實時熒光定量PCR(RT-qPCR)實驗 用RNA-Easy試劑盒提取總RNA。采用相對定量法定量靶基因,以 β-actin 為內參照,計算待測EAAT2和mTOR mRNA 相對表達量。引物序列:EAAT2的上游引物為5′-ATGCTCCTCATTCTCACAG-3′,下游引物為5′-CTACATTGACCGAAGTTCTC-3′;mTOR的上游引物為5’-CTGGGACTCAAATGTGTGCAGTTC-3’,下游引物為5’-GAACAATAGGGTGAATGATCCGGG-3’;β-actin的上游引物為5′-GCTGAGAGGGAAATCGTGCGTGAC-3′ ,下游引物為5′-GGAGGAAGAGGATGCGGCAGTGG-3′。qPCR用ABI-棱柱7700序列系統檢測,mRNA水平采用2-ΔΔCt法計算。
2.4Western blot分析 原代星形膠質細胞裂解,勻漿,離心后取上清液進行蛋白變性,以15% SDS聚丙烯酰胺凝膠電泳分離蛋白,將分離的蛋白轉移至硝酸纖維素膜,5% 脫脂奶粉封閉1 h,加EAAT2、mTOR和β-actin抗體,4 ℃孵育過夜,用TBST充分洗滌后,加入辣根過氧化物酶標記 II 抗37 ℃孵育1 h,TBST 充分洗滌,ECL 發光顯影;用CIS 凝膠圖像處理系統進行分析。
3統計學處理
應用SPSS 16.0統計分析軟件,研究結果以均數±標準差(mean±SD)表示,計量資料用完全隨機設計單因素方差分析(one-way ANOVA);計數資料用χ2檢驗;以P<0.05為差異有統計學意義。
結 果
1DA對星形膠質細胞EAAT2表達的影響
RT-qPCR的結果顯示DA干預的PCAs中EAAT2的mRNA水平顯著減少,并且呈時間依賴性;Western blot實驗結果顯示,DA干預的PCAs中EAAT2蛋白表達水平也隨時間逐漸減少,見圖1。
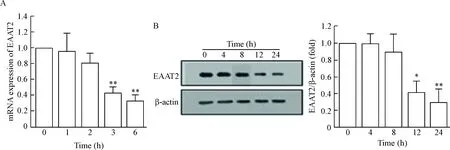
Figure 1. The change of EAAT2 mRNA (A) and protein (B) expression in the PCAs after treatment with DA. Mean±SD.n=3.*P<0.05,**P<0.01vs0 h.
圖1DA干預PCAs后EAAT2的mRNA和蛋白表達水平的變化
2DA對星形膠質細胞mTOR表達的影響
RT-qPCR的結果顯示,DA干預的PCAs中mTOR的mRNA水平顯著減少,并且呈時間依賴性;Western blot實驗結果與mRNA結果趨勢一致,mTOR蛋白表達水平顯著減少,并且呈時間依賴性,見圖2。
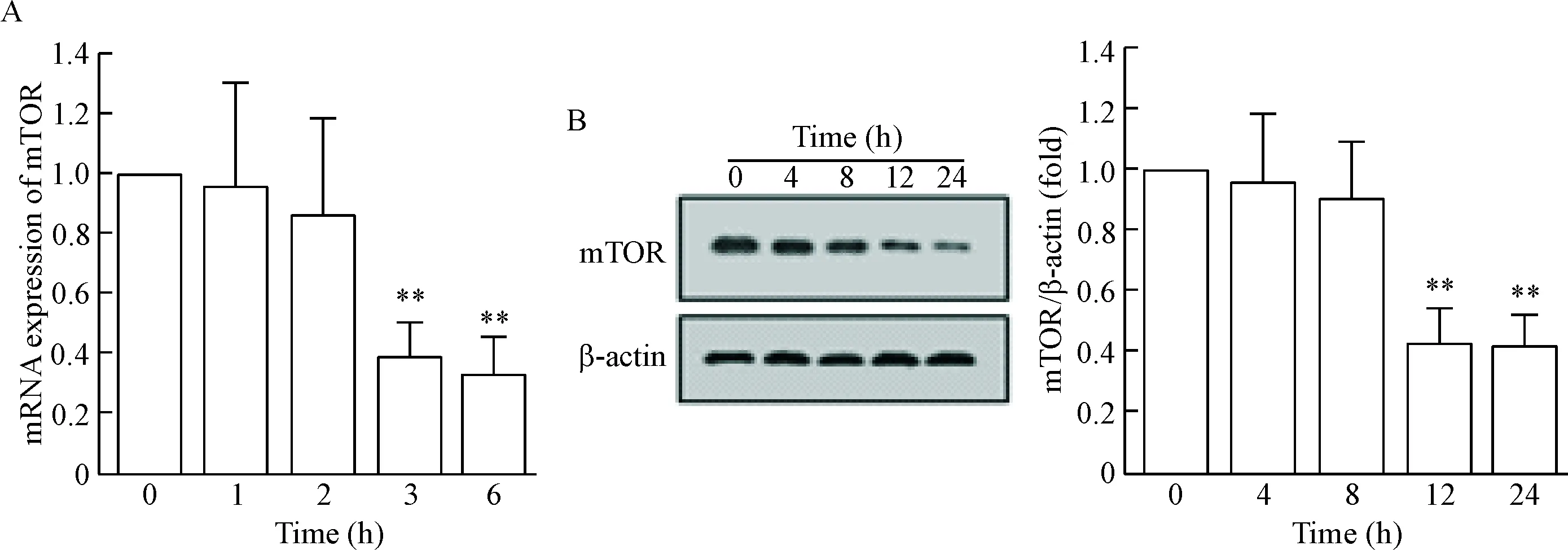
Figure 2. The change of mTOR mRNA (A) and protein (B) expression in the PCAs after treatment with DA. Mean±SD.n=3.*P<0.05,**P<0.01vs0 h.
圖2DA干預PCAs后mTOR的mRNA和蛋白表達水平的變化
3DA對mTOR抑制/激活后的PCAsEAAT2表達的影響
mTOR拮抗劑雷帕霉素與DA干預的PCAs共培養后的RT-qPCR和Western blot實驗分析結果顯示,EAAT2的mRNA水平和蛋白表達都顯著降低;mTOR激活劑MHY1485與DA干預的PCAs共培養后,EAAT2的mRNA水平和蛋白表達顯著上升,見圖3。

Figure 3. The change of EAAT2 mRNA (A) and protein (B) expression in the PCAs after treatment with rapamycin/MHY1485 and DA. Mean±SD.n=3.*P<0.05,**P<0.01vsDA alone.
圖3雷帕霉素/MHY1485與DA干預PCAs后EAAT2的mRNA和蛋白表達水平的變化
4DA對mTOR抑制/激活后的PCAs對Glu攝取能力的影響
如圖4所示,雷帕霉素與DA干預的PCAs共培養后,細胞外的Glu水平顯著上升。MHY1485與DA干預的PCAs共培養后,其細胞外的Glu含量顯著下降。
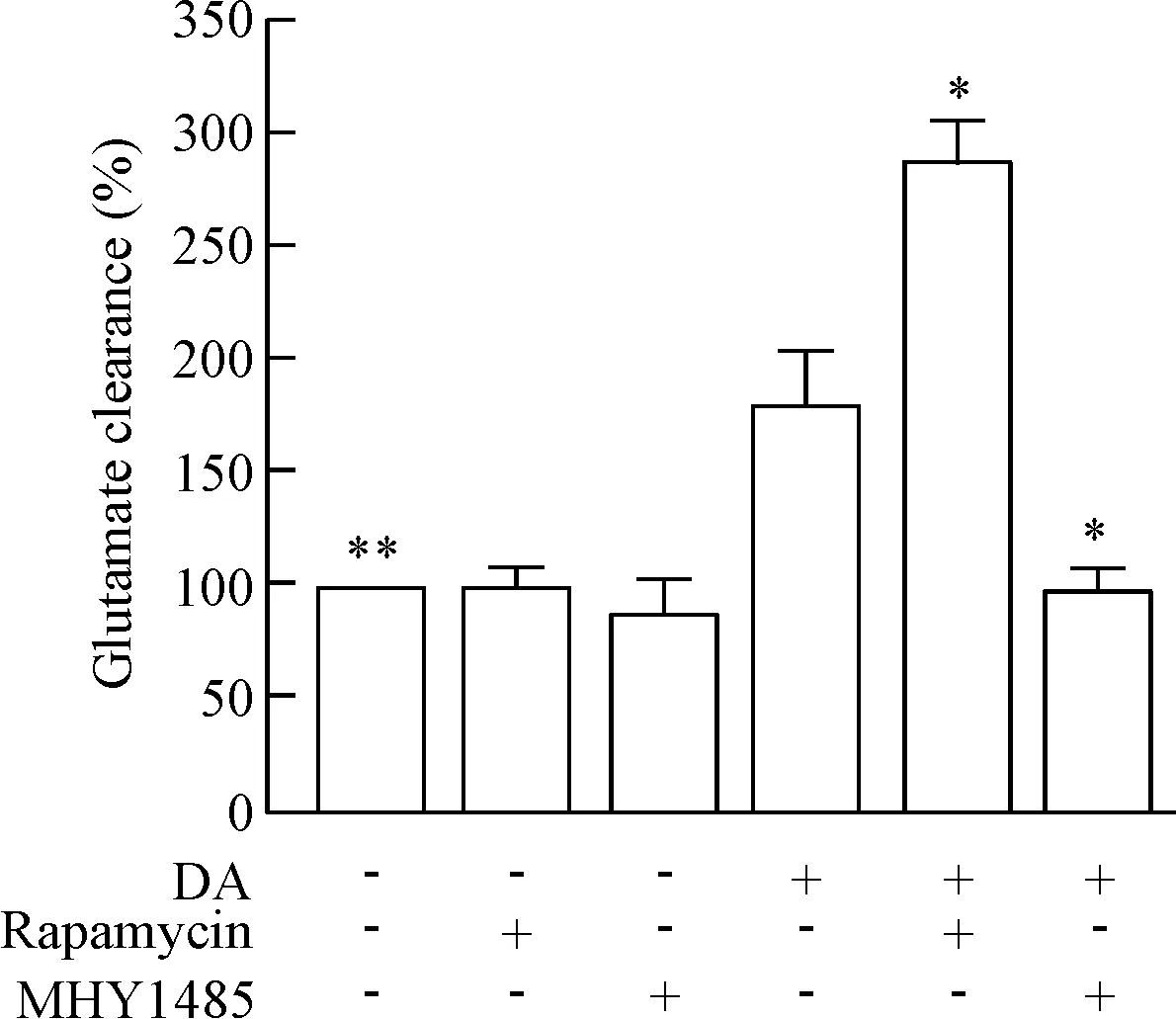
Figure 4. The Glu content in the culture supernatant of the PCAs after treated with rapamycin/MHY1485 and DA. Mean±SD.n=3.*P<0.05,**P<0.01vsDA alone.
圖4雷帕霉素/MHY1485與DA干預PCAs后上清液Glu含量的比較
討 論
谷氨酸是大腦主要的興奮性神經遞質,過多的Glu蓄積在細胞間隙會引起神經興奮性中毒而受損,將會引發多種神經退行性疾病。星形膠質細胞能夠通過突觸間隙的EAAT2清除排到細胞外大約90%的谷氨酸[10]。已有報道星形膠質細胞mTOR分子水平的改變與星形膠質細胞EAAT2表達水平和功能相關[11]。
維持Glu內穩態的主要通路是通過EAAT1和EAAT2再攝取[12],Glu釋放入突觸間隙后,在神經元活動期間需要通過興奮性氨基酸轉運體快速攝取,EAAT1和EAAT2會優先清除位于星形膠質細胞外大部分的Glu[12]。這些轉運體在控制興奮信號和阻止細胞外Glu興奮毒性聚集方面起著至關重要的作用[12-13],所以EAAT1和EAAT2的失活可能會導致Glu異常的累積,從而損傷星形膠質細胞。已有報道表明,EAAT2的長期損傷會促進中樞神經系統功能紊亂[14]。非選擇性的EAATs抑制劑會誘導EAAT2功能紊亂,明顯影響行為反應[15]和誘發神經退行性病變[16]。在我們的實驗結果中,EAAT2的mRNA和蛋白質表達因DA干擾顯著降低,與Danbolt等[12]所報道的EAAT2蛋白表達降低后干擾Glu的清除相符。我們的實驗結果顯示Glu的清除能力會因DA干擾減弱。基于大腦釋放增加的DA會導致大腦興奮性氨基酸轉運異常的現象,我們觀測到DA干預星形膠質細胞對Glu攝取的最明顯改變是EAAT2的翻譯和轉運功能下降,從而降低對Glu攝取能力。前期實驗中我們已經發現DA通過誘導星形膠質細胞中NADPH氧化酶活化產生活性氧,導致蛋白質酪氨酸酸化,改變星形膠質細胞功能[17]。
DA干預下的星形膠質細胞mTOR表達降低[18];mTOR可以調控Glu轉運體在星形膠質細胞的表達[18],mTOR可能涉及DA介導的EAAT2的下調。DA干預星形膠質細胞后協同mTOR下調EAAT2,從而降低Glu清除能力而導致Glu水平增加。已知雷帕霉素為mTOR競爭者,能夠調節Glu的清除能力[11]。因此mTOR在DA誘導的過量Glu的釋放中有著重要的作用。DA可以降低星形膠質細胞mTOR的表達,這會造成mTOR/EAAT2受損及伴隨著Glu清除的受阻。
綜上所述,本次我們闡明DA是通過mTOR-EAAT2這一信號通路影響Glu的攝取從而誘導Glu的蓄積,Glu在細胞間隙聚集會引起神經興奮毒性,從而使星形膠質細胞受損,這涉及很多的神經退行性疾病[19-20]。我們的實驗研究DA體外干預星形膠質細胞,明確DA抑制星形膠質細胞對Glu的攝取,引起膠質細胞興奮中毒,且星形膠質細胞作為中樞神經系統中含量最豐富的細胞在阿爾茨海默癥的發病過程中起十分重要的作用[21],可以更好地確定DA對腦內谷氨酸攝取能力的影響及相關疾病的發病機制。
鑒于星形膠質細胞在多種神經退行性疾病中的作用越來越受到重視,后續我們還需要進一步了解DA體內干預大鼠海馬組織的星形膠質細胞的具體機制,將有助于改善和治療神經退行性疾病。
[1] Negis Y, Karabay A. Expression of cell cycle proteins in cortical neurons: correlation with glutamate-induced neurotoxicity [J]. Biofactors, 2016, 42(4):358-367.
[2] Van Laar VS, Roy N, Liu A, et al. Glutamate excitoto-xicity in neurons triggers mitochondrial and endoplasmic reticulum accumulation of Parkin, and, in the presence ofN-acetyl cysteine, mitophagy [J]. Neurobiol Dis, 2015, 74:180-193.
[3] Mao XY, Zhou HH, Li X, et al. Huperzine a alleviates oxidative glutamate toxicity in hippocampal HT22 cells via activating BDNF/TrkB-dependent PI3K/Akt/mTOR signaling pathway [J]. Cell Mol Neurobiol, 2016, 36(6): 915-925.
[4] Lourhmati A, Buniatian GH, Paul C, et al. Age-depen-dent astroglial vulnerability to hypoxia and glutamate: the role for erythropoietin [J]. PLoS One, 2013, 8(10): e77182.
[5] 張 運, 王莉莉, 趙 茜, 等. GRK5對大鼠星形膠質細胞活化的作用及其機制研究[J]. 中國病理生理雜志, 2014, 30(4):577-583.
[6] 王 磊, 劉 平, 慕永平, 等. 二甲基亞硝胺大鼠肝纖維化中醫方證研究[J]. 中醫雜志, 2006, 47(12): 929-932.
[7] Ding S, Wang W, Wang X, et al. Dopamine burden triggers neurodegeneration via production and release of TNF-α from astrocytes in minimal hepatic encephalopathy [J]. Mol Neurobiol, 2016, 53(8):5324-5343.
[8] Ding S, Liu L, Jing H, et al. Dopamine from cirrhotic liver contributes to the impaired learning and memory ability of hippocampus in minimal hepatic encephalopathy [J]. Hepatol Int, 2013, 7(3):923-936.
[9] Bernabeu R, Schmitz P, Faillace MP, et al. Hippocampal cGMP and cAMP are differentially involved in memory processing of inhibitory avoidance learning [J]. Neuroreport, 1996, 7(2):585-588.
[10] Anderson CM, Swanson RA. Astrocyte glutamate transport: Review of properties, regulation, and physiological functions [J]. Glia, 2000, 32(1):1-14.
[11] Cisneros IE, Ghorpade A. Methamphetamine and HIV-1-induced neurotoxicity: role of trace amine associated receptor 1 cAMP signaling in astrocytes [J]. Neuropharmacology, 2014, 85:499-507.
[12] Danbolt NC. Glutamate uptake[J]. Prog Neurobiol, 2001, 65(1):1-105.
[13] Karklin Fontana AC, Fox DP, Zoubroulis A, et al. Neuroprotective effects of the glutamate transporter activator (R)-(-)-5-methyl-1-nicotinoyl-2-pyrazoline (MS-153) following traumatic brain injury in the adult rat [J]. J Neurotrauma, 2016, 33(11):1073-1083.
[14] Cassano T, Serviddio G, Gaetani S, et al. Glutamatergic alterations and mitochondrial impairment in a murine mo-del of Alzheimer disease [J]. Neurobiol Aging, 2012, 33(6):1121.e1-1121.e12.
[15] Hayase T, Yamamoto Y, Yamamoto K. Brain excitatory amino acid transporters (EAATs) and treatment of methamphetamine toxicity [J]. Nihon Arukoru Yakubutsu Igakkai Zasshi, 2003, 38(6):498-511.
[16] Assous M, Had-Aissouni L, Gubellini P, et al. Progressive Parkinsonism by acute dysfunction of excitatory amino acid transporters in the rat substantia nigra [J]. Neurobiol Dis, 2014, 65:69-81.
[17] Ding S, Yang J, Liu L, et al. Elevated dopamine induces minimal hepatic encephalopathy by activation of astrocytic NADPH oxidase and astrocytic protein tyrosine nitration [J]. Int J Biochem Cell Biol, 2014, 55:252-263.
[18] María López-Colomé A, Martínez-Lozada Z, Guillem AM, et al. Glutamate transporter-dependent mTOR phosphorylation in Müller glia cells [J]. ASN Neuro, 2012, 4(5): e00095.
[19] Engel M, Do-Ha D, Muoz SS, et al. Common pitfalls of stem cell differentiation: a guide to improving protocols for neurodegenerative disease models and research [J]. Cell Mol Life Sci, 2016, 73(19):3693-3709.
[20] Abolhassani N, Leon J, Sheng Z, et al. Molecular pathophysiology of impaired glucose metabolism, mitochondrial dysfunction, and oxidative DNA damage in Alzheimer’s disease brain [J]. Mech Ageing Dev, 2016, 161(Pt A):95-104.
[21] 李夏春, 彭敏峰, 高麗華, 等. 上調星形膠質細胞中 PP2A 對APP/PS1 雙轉基因小鼠的神經保護作用[J]. 中國病理生理雜志, 2016, 32(7):1189-1194.
(責任編輯: 盧 萍, 羅 森)
Effectofdopamineonglutamate-uptakeabilityofastrocytesbyregulatingmTOR-EAAT2pathway
WEN Fang-fang, XU Zhu, LIU Le-ping, YANG Jan-jing, DING Sai-dan
(ZhejiangProvincialKeyLaboratoryofAgingandNeurologicalDiseaseResearch,TheFirstAffiliatedHospitalofWenzhouMedicalUniversity,Wenzhou325000,China.E-mail:firstdsdan@hotmail.com)
AIM: To investigate the effect of dopamine (DA) on the glutamate (Glu)-uptake ability of astrocytes, and the role of mammalian target of rapamycin (mTOR)-excitatory amino acid transporter 2(EAAT2) pathway in this process.METHODSExtracellular Glu levels in DA-treated primary cortical astrocytes (PCAs) were measured by a fluorimetric method. The relative expression of EAAT2 and mTOR at mRNA and protein levels was measured by RT-qPCR and Western blot. PCAs stimulated with or without DA in the presence or absence of mTOR antagonist rapamycin or mTOR agonist MHY1485 were used to determine the expression of mTOR and EAAT2, and Glu content in the culture supernatant was also measured.RESULTSThe expression of mTOR in DA-treated PCAs was decreased, the expression of EAAT2 was also decreased. Extracellular Glu levels of DA-treated PCAs were elevated significantly. When the PCAs were stimulated with DA in the presence of rapamycin, the expression of EAAT2 was decreased, and the levels of extracellular Glu was significantly increased. In the presence of MHY1485, the expression of EAAT2 was elevated, and significant decrease in the levels of extracellular Glu was also observed.CONCLUSIONDA interacts with mTOR-EAAT2 pathway to reduce the Glu-uptake ability of the astrocytes, and causes extracellular Glu accumulation, ultimately destroys the function of astrocytes.
Dopamine; Glutamate; Astrocytes
R741; R363
A
10.3969/j.issn.1000- 4718.2017.10.003
1000- 4718(2017)10- 1746- 05
2017- 01- 22
2017- 03- 27
國家自然科學基金資助項目(No. 81671042; No. 81300308)
△通訊作者 Tel: 15888209333; E-mail: firstdsdan@hotmail.com
雜志網址: http://www.cjpp.net
