多花黃精生殖生物學特性研究
2017-11-07 02:47:05劉艷羅敏秦民堅謝國勇章文偉陳強肖波李娜
中國中醫(yī)藥信息雜志 2017年11期
關鍵詞:研究
劉艷,羅敏,秦民堅,謝國勇,章文偉,陳強,肖波,李娜
?
多花黃精生殖生物學特性研究
劉艷1,羅敏1,秦民堅2,謝國勇2,章文偉1,陳強1,肖波1,李娜1
1.重慶市藥物種植研究所,中國醫(yī)學科學院藥用植物研究所重慶分所,重慶 408435; 2.中國藥科大學,江蘇南京 210009
研究多花黃精的生殖生物學特性。在花期定點定株觀察記錄多花黃精的開花習性,用TTC法檢測其開花各時期的花粉活力,采用聯(lián)苯胺-過氧化氫法檢測不同開花時期柱頭的可授性,估算雜交指數并計算其花粉胚珠比,通過6種不同的人工授粉方式處理確定多花黃精的繁育系統(tǒng)類型。多花黃精在4月5日左右現蕾,4月10日左右開花,5月上旬陸續(xù)開始結實,花期約30~40 d。自花藥未開裂時花粉即開始具有活力,花朵枯萎后花粉失去活力,開花當日花粉活力達73.06%±5.19%。自花朵未開放時柱頭即具有活性,花朵開始衰敗時柱頭活性減弱消失。多花黃精的雜交指數為4,據Dafni標準其繁育系統(tǒng)為部分自交親和,異交,需要傳粉者;其花粉胚珠比為1799.20±445.54,據Cruden標準其繁育系統(tǒng)為兼性異交。人工控制授粉試驗結果表明多花黃精具有無融合生殖、自交和異交3種生殖方式,其結實率受外界條件影響較大,總體結實率較低,自然狀態(tài)下僅為17.46%。多花黃精具有無融合生殖、有性生殖與無性生殖3種生殖方式。
多花黃精;開花習性;花粉活力;柱頭活性;繁育系統(tǒng)
多花黃精Hua隸屬于百合科Liliaceae黃精屬,為《中華人民共和國藥典》收載黃精的原植物之一。屬藥食同源植物,為我國傳統(tǒng)大宗中藥材。其性平,味甘,入脾、肺、腎經。功效養(yǎng)陰潤肺、補脾益氣、滋腎填精,主治陰虛勞嗽、肺燥咳嗽、脾虛乏力、食少口干、內熱消渴、腎虧腰膝酸軟、陽痿遺精、耳鳴目暗、須發(fā)早白、體虛羸瘦、風癩癬疾。多花黃精為多年生草本植物,生于山林、灌叢、溝谷旁的陰濕肥沃土壤中,分布于我國中南地區(qū)及江蘇、安徽、浙江、江西、福建、四川、重慶、貴州等地。
目前對多花黃精的研究包括對其生理生態(tài)學方面的研究[1-2]、種植技術研究[3-7]、種子繁殖技術研究[8]、分子生物學研究[9]、組培快繁方面的研究[10]等。目前對多花黃精生殖生物學特性以及繁育系統(tǒng)方面的研究報道非常少,田啟建等[11-12]研究了栽培多花黃精的物候期及開花結實特性,發(fā)現多花黃精蕾期為45 d左右,花期約20 d,結實率為58.37%,落果嚴重,田間成熟果實量非常小。
多花黃精種子成熟率低,目前在藥材生產中多花黃精的栽培主要以無性繁殖為主(依靠地下莖)[2,11],但是長期的無性繁殖弊端比較大,如品種退化、病蟲害發(fā)生嚴重等,使藥材產量和質量受到嚴重影響。本研究著重對多花黃精的花粉活力、柱頭活性及繁育系統(tǒng)等進行檢測,研究其有性生殖特性及繁育系統(tǒng)特征,為多花黃精在規(guī)范化栽培中能盡快解決無性繁殖的弊端、提高有性繁殖的比率提供依據,并為多花黃精后續(xù)的種質資源開發(fā)提供繁育系統(tǒng)方面的技術參考。
1 材料
試驗地位于重慶市南川區(qū)三泉鎮(zhèn)石門溝,為重慶市藥物種植研究所的黃精種質資源圃,栽植面積為360 m2。所用的多花黃精栽種2年,以根莖為繁殖材料,種莖來自湖南省新化縣,開花時經重慶市藥物種植研究所植物分類學專家劉正宇研究員鑒定為多花黃精Hua。于2015年4-5月和2016年4-5月進行開花、結實物候觀察及人工控制授粉試驗等。
2 方法與結果
2.1 開花習性觀察
從2015年4月1日開始,每3 d觀察1次現蕾情況,現蕾之后,隨機選取5株進行標記,觀察記錄其開花順序、初花、盛花、末花及結實時間等。同時對即將開放的6朵花蕾掛牌,定位觀察并記錄花朵個體水平的開花進程。
多花黃精的花蕾與花均按從下往上的順序生出和開放,花期為4月上旬至5月下旬。2015年4月5日左右開始現蕾,4月10日左右開始開花,單花花期2~2.5 d。花朵于晚上9~10點開始張開,至次日早8點完全張開,持續(xù)24 h后開始凋謝,花朵凋謝時花瓣先閉合,1 d后整朵花枯萎。盛花期為4月底至5月初,持續(xù)約7~10 d。5月上旬陸續(xù)開始結實,有明顯的落花落果現象,結實率不高。
2.2 花粉活力及柱頭可授性檢測
于盛花期隨機采集多花黃精開花前后不同大小的新鮮花朵,將少量花粉置于載玻片上,滴1~2滴0.5%TTC溶液,充分混勻后蓋上蓋玻片,置于35 ℃恒溫箱中15 min,之后在顯微鏡下觀察。選取5個視野,每個視野花粉數約為70~100粒,凡被染成紅色的花粉為具有活力的花粉,淡紅色或無色者為不具有生活力的花粉。花粉活力(%)=紅色花粉數÷花粉總數×100%。結果見表1。
采用聯(lián)苯胺-過氧化氫法檢測不同開花時期柱頭的可授性。隨機選取開花前至開花后不同生長時期的花朵,在凹面載玻片中加入體積比為1%聯(lián)苯胺∶3%過氧化氫∶水=4∶11∶22的反應液,將柱頭浸入其中,觀察柱頭是否呈現藍色并產生大量氣泡。結果見表1。
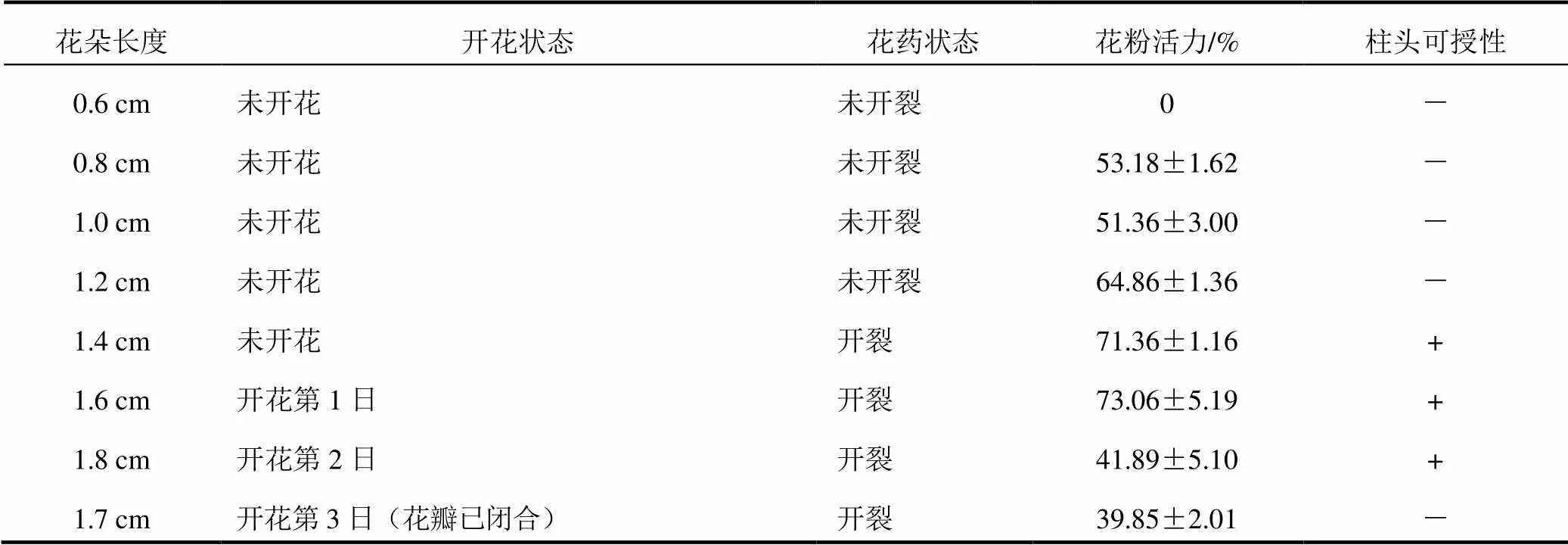
表1 不同開花時期多花黃精的花粉活力與柱頭可授性
注:“+”表示柱頭具有可授性,“-”表示柱頭不具可授性
檢測結果顯示,多花黃精自花藥未開裂時花粉即開始具有活力,經TTC染色后花粉粒呈現紅色,花朵枯萎后,花粉失去活力。根據試驗結果(見表1),花長0.6 cm以下的花粉不具有活力,花長0.8 cm以上開始具有活力,且隨著花朵的逐漸成熟花粉活力逐漸增強,至開花當天花粉活力最高,為73.06%±5.19%,而開花次日至花朵凋謝,花粉活力逐漸下降,開花第2日花粉活力已下降至41.89%±5.10%,遠不及開花當日的花粉活力。
多花黃精自花朵未開放時柱頭即具有活性,一直延續(xù)至開花時,當花朵開始衰敗時,柱頭活性減弱消失。根據聯(lián)苯胺-過氧化氫法檢測結果(見表1),花長1.3 cm以下的柱頭沒有可授性,花長1.4 cm以上的柱頭才具有可授性,柱頭浸入液體中呈現藍色,柱頭周圍產生大量氣泡。直至開花第3日花朵開始凋謝,柱頭活性消失,不再具有可授性。
2.3 雜交指數的估算
按照Dafni的標準[13]進行花朵直徑、花朵大小及開花行為的測定及繁育系統(tǒng)的評判。①花朵或花序直徑:<1 mm記為0,1~2 mm記為1,>2~6 mm記為2,>6 mm記為3。②花藥開裂時間與柱頭可授期之間的時間間隔:同時或雌蕊先熟記為0;雄蕊先熟記為1。③柱頭與花藥的空間位置:同一高度記為0;空間分離記為1。以上三者之和為雜交指數(outcrossing index,OCI)。OCI=0時,繁育系統(tǒng)為閉花受精;OCI=1時,繁育系統(tǒng)為專性自交;OCI=2時,繁育系統(tǒng)為兼性自交;OCI=3時,繁育系統(tǒng)為自交親和,有時需要傳粉者;OCI=4時,繁育系統(tǒng)為部分自交親和,異交,需要傳粉者。
多花黃精的花朵直徑<6 mm,測量值記為2;雄蕊先熟,花粉在柱頭具有可授性之前已經具有活力,測量值記為1;開花時柱頭與花藥在空間位置上是分離的,柱頭位置比花藥高約1 mm,測量值記為1。因此,多花黃精的OCI值為4,其繁育系統(tǒng)為部分自交親和,異交,需要傳粉者。
2.4 花粉胚珠比
隨機選取5朵花藥未開裂的花。將1朵花中所有的花藥置于離心管中,加200 μL染液,將花藥搗碎使花粉散出成為懸濁液,在渦旋儀上震蕩60 s,然后用移液槍取5份1 μL的樣本于載玻片上,于顯微鏡下觀察計數,取平均值。1朵花中總的花粉數=1 μL染液中的花粉數×200。將1朵花的子房在解剖鏡下仔細剖開,計數胚珠數。花粉胚珠比(P/O)=花粉數÷胚珠數。根據Cruden的標準[14]進行繁育系統(tǒng)的判斷:P/O為2.7~5.4,繁育系統(tǒng)為閉花受精;P/O為18.1~39.0,為專性自交;P/O為31.9~396,為兼性自交;P/O為244.7~2588,為兼性異交;P/O為2108~195 525,為專性異交。
結果多花黃精P/O為1799.20±445.54。依據Cruden的標準可以判斷,多花黃精的繁育系統(tǒng)為兼性異交。
2.5 繁育系統(tǒng)檢測
于盛花期隨機選取花朵,分6個處理進行人工控制授粉試驗。①不去雄,不套袋:檢測自然狀態(tài)下授粉情況。②去雄,套袋,不授粉:檢測是否具有無融合生殖。③不去雄,套袋:檢測是否具有自發(fā)的自花授粉。④人工自花授粉:檢測自花自交情況。⑤人工同株異花授粉:檢測同株異花自交情況。⑥人工異株異花授粉:檢測異交情況。
開花前進行去雄操作并用6 cm×8 cm硫酸紙袋將單花套住,開花時完成人工控制授粉試驗后繼續(xù)將單花套住,套袋及授粉用的花朵均選取完整沒有開裂的花藥。自然對照采用315朵花,其余每處理幾十朵花不等。3周后統(tǒng)計結實率,子房膨大者視為結實。結實率(%)=果實數÷花數×100%。結果見表2。
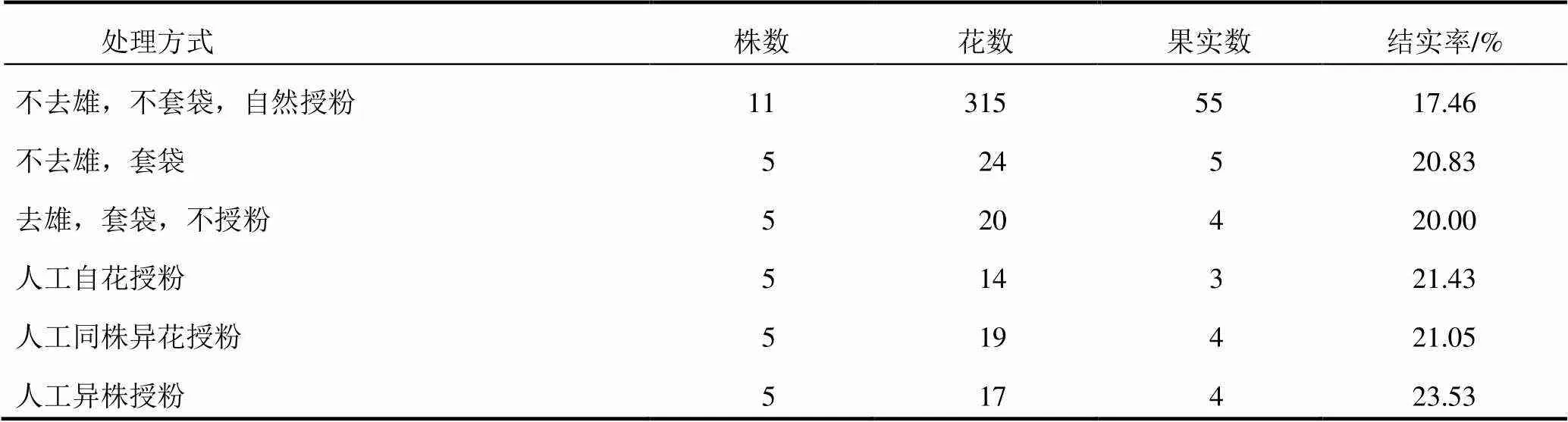
表2 多花黃精人工控制授粉結實情況
人工控制授粉試驗結果顯示:去雄、套袋的多花黃精可以結實,結實率為20.00%,說明其存在無融合生殖現象;不去雄、套袋的多花黃精結實率為20.83%,與去雄、套袋的結實率無明顯差異,說明缺少傳粉媒介輔助時自花花粉對結實率的貢獻較小;人工自花授粉、同株異花授粉和異株異花授粉的結實率分別為21.43%、21.05%和23.53%,均略高于去雄、套袋的結實率,說明經過傳粉媒介的作用,自花、同株異花、異株異花的花粉均能相應提高結實率。多花黃精具有無融合生殖、自交和異交3種繁殖方式。多花黃精自然授粉狀態(tài)下的結實率為17.46%,其結實率受外界條件影響較大,大風大雨都會導致嚴重的落花落果,因此總體而言多花黃精的結實率比較低。
3 討論
多花黃精的花期約為2個月,單花能開2~2.5 d,盛花期持續(xù)約7~10 d,這與田啟建等[11-12]對栽培多花黃精的物候期研究結果基本吻合。多花黃精在花長0.8 cm以上時花粉即開始具有活力,開花當日花粉活力達到73.06%±5.19%,一直延續(xù)至花朵枯萎后,花粉失去活力。花長1.5 cm以上的柱頭具有可授性,當花朵開始衰敗時,柱頭活性減弱消失。多花黃精的結實率受外界條件影響較大,大風大雨都會導致嚴重的落花落果,本研究6種人工控制授粉試驗中多花黃精的結實率均比較低,約為20%,這與田啟建等[11-12]研究得出的“落果嚴重,田間成熟果實量非常小”的結論一致。本研究中的結實率比該研究發(fā)現的多花黃精結實率58.37%要低得多,其原因可以歸結為以下幾點:①試驗地位于重慶市南川區(qū)三泉鎮(zhèn),常年風力在2級以上,經常有持續(xù)強風,大風加強了落花落果。②試驗地的土壤相對干燥,土壤肥力不高。③栽培年限對多花黃精植株的開花數和結實率具有顯著影響[11],試驗用多花黃精利用塊莖栽種了2~3年,屬于低齡,因此造成結實率相對偏低。
多花黃精的雜交指數為4,根據Dafni的判定標準其繁育系統(tǒng)為部分自交親和,異交,需要傳粉者。其花粉胚珠比為1799.20±445.54,據Cruden的標準其繁育系統(tǒng)為兼性異交。這2種方法檢測到的繁育系統(tǒng)結果基本一致,也與人工控制授粉試驗的結果相吻合,多花黃精具有無融合生殖現象,并且經過傳粉媒介的作用,也能進行自交和異交。本研究觀察發(fā)現,多花黃精的傳粉昆蟲主要為蜂類,有熊蜂sp.和淡脈隧蜂sp.等。加之目前應用最多的地下莖繁殖,多花黃精既存在有性繁殖也存在無性繁殖。有性繁殖與無性繁殖的權衡與生境關系密切,在對典型植物珠芽蓼的研究中發(fā)現,在環(huán)境惡劣的生境中珠芽蓼減少了投資到營養(yǎng)生長的資源并增加了對繁殖器官的投資,而且對花的資源投資優(yōu)先于對珠芽的資源投資,表明高山植物在適應極端環(huán)境過程中有性繁殖比無性繁殖具有更為重要的作用[15]。無融合生殖是對不良環(huán)境的短暫適應,是有性生殖受阻的一種生殖補償,由于沒有外來基因導入,缺乏遺傳多樣性,在長期的自然選擇中會逐漸被淘汰[16]。多花黃精的多種繁殖方式并存,有利于其適應不同的生境類型從而存活下來,確保其種群的繁衍。
多花黃精種子的成熟率低,同時由于其存在較嚴重的休眠現象,需要2~3年才能出苗,所以目前多花黃精的栽培主要以無性繁殖為主,但長期無性繁殖所導致的品種退化、病蟲害嚴重等問題使藥材產量和質量受到嚴重影響。想破除這些弊端,唯有在實際栽培中加強多花黃精的有性繁殖方式,除了自然的昆蟲授粉外,輔助以人工授粉,提高其結實率。同時在實際生產中,針對落花落果問題要加強防護措施,如增加防護網、套種高大農作物等,減少自然環(huán)境條件導致的落花落果。本研究為多花黃精的規(guī)范化栽培生產及優(yōu)良種質資源的篩選提供了依據和技術參考,今后將重點關注多花黃精優(yōu)良品種的選育及如何打破其種子休眠,加快種子萌發(fā)并提高發(fā)芽率,為中藥材市場提供品種優(yōu)良的高質量種子。
[1] 駱緒美,余國清,郭婉琳,等.安徽4種黃精屬植物光合生理特性的研究[J].安徽林業(yè)科技,2012,38(4):3-5,12.
[2] 陳芳軟.不同生境條件下多花黃精生長特征及光合特性研究[D].福州:福建農林大學,2013.
[3] 樊艷榮,陳雙林,楊清平,等.毛竹林下多花黃精種群生長和生物量分配的立竹密度效應[J].浙江農林大學學報,2013,30(2):199-205.
[4] 樊艷榮,陳雙林,楊清平,等.毛竹材用林林下植被群落結構對多花黃精生長的影響[J].生態(tài)學報,2014,34(6):1471-1480.
[5] 戴琴,王曉霞,黃勤春,等.毛竹林下多花黃精仿野生栽培技術[J].中國現代中藥,2014,16(3):205-207.
[6] 羅奮容.杉木、多花黃精復合經營技術探討[J].福建農業(yè)科技,2015, 46(6):25-28.
[7] 王聲淼,吳劍鋒,吳應齊.浙南閩北多花黃精規(guī)范化栽培技術[J].安徽農業(yè)科學,2015,43(22):14,47.
[8] 周建金,羅曉鋒,葉煒,等.多花黃精種子繁殖技術的研究[J].種子, 2013,32(1):111-113.
[9] 張紅梅,邵元華,龍?zhí)鹛?等.多花黃精ISSR反應體系的建立及正交優(yōu)化設計[J].安徽農業(yè)大學學報,2012,39(3):412-416.
[10] 萬學鋒,陳菁瑛.多花黃精組培快繁技術初探[J].中國現代中藥, 2013,15(10):850-852.
[11] 田啟建,趙致,谷甫剛.栽培黃精開花結實習性研究[J].種子,2009, 28(1):29-31.
[12] 田啟建,趙致,谷甫剛.栽培黃精物候期研究[J].中藥材,2010,33(2):168-170.
[13] DAFNI A. Pollination ecology[M]. New York:Oxford Univ Press, 1992:1-57.
[14] CRUDEN R W. Pollen-ovule ratio:a conservative indicator of breeding systems in flowering plants[J]. Evolution,1977,31(1):32-46.
[15] 范鄧妹,楊永平.不同生境下珠芽蓼(蓼科)的繁殖策略比較[J].云南植物研究,2009,31(2):153-157.
[16] 寧偉,張建,吳志剛,等.丹東蒲公英專性無融合生殖特性[J].植物學報,2014,49(4):417-423.
Study on Reproductive Biology ofHua
LIU Yan1, LUO Min1, QIN Min-jian2, XIE Guo-yong2, ZHANG Wen-wei1, CHEN Qiang1, XIAO Bo1, LI Na1
To study the reproductive biology of Chinese herbal medicineHua.The flowering habit ofwas observed and recorded regularly. The pollen viability at different flowering periods was detected by TTC. The stigma activity at different flowering periods was measured by the benzidine-hydrogen peroxide method. The hybridization index was estimated and pollen-ovule ratio was calculated. The breeding system ofwas determined by six different artificial pollination treatments.The flower buds ofappeared at April 5. They bloomed at April 10 and fruited early in May, flowering about 30–40 days. The pollen began to have viability since the anthers were not cracked, and loss viability after flowers withered. The pollen viability at the bloom day was about 73.06%±5.19%. The stigmas began to have activity before flowers opening and lost it when the flowers began to decline. The hybridization index was 4, so the breeding system ofHua was partly self-compatible and xenogamy needing pollinator according to Dafni’s standard. The pollen-ovule ratio was 1799.20±445.54, therefore the breeding system ofHua was facultative xenogamy according to Cruden’s standard. The results of artificial pollination experiments showed thatHua had three breeding methods, including apomixis, selfing and outcrossing. The fruit setting rate ofHua was greatly influenced by external conditions such as strong wind and rain. Generally,Hua had a low fruit-set percentage because of obvious flower and fruit dropping. The natural fruit setting rate was only 17.46%.The coexistence of apomixes, sexual and asexual reproduction inare the survival strategy adapting to different habitat types.
Hua; flowering habit; pollen viability; stigma activity; breeding system
10.3969/j.issn.1005-5304.2017.11.017
R282.2
A
1005-5304(2017)11-0071-04
(2017-03-10)
(2017-04-12;編輯:陳靜)
重慶市衛(wèi)生計生委中醫(yī)藥科技項目(zy201402124、zy201702147);西部之光訪問學者計劃項目(2016年)
章文偉,E-mail:ssf333@126.com
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
