冬小麥高花藥培養力基因型的篩選
2017-11-08 07:36:55趙林姝劉錄祥古佳玉謝永盾郭會君趙世榮李軍輝熊宏春
麥類作物學報 2017年10期
趙林姝,劉錄祥,古佳玉,謝永盾,郭會君,趙世榮,李軍輝,熊宏春
(中國農業科學院作物科學研究所,北京 100081)
冬小麥高花藥培養力基因型的篩選
趙林姝,劉錄祥,古佳玉,謝永盾,郭會君,趙世榮,李軍輝,熊宏春
(中國農業科學院作物科學研究所,北京 100081)
為了篩選小麥花培育種骨干親本,減輕花培育種的基因型依賴性問題,對74個冬小麥品種(系)的5個花藥培養力性狀進行了鑒定,并對5個性狀進行了相關性分析。結果表明,74個基因型的愈傷組織誘導率、綠苗分化率、綠苗產率、白苗分化率及白苗產率變化范圍分別為0~43.17%、0~139.29%、0~20.83%、0~63.33%、0~7.17%,各花藥培養力性狀在所研究基因型中差異明顯,存在基因型依賴性,其中綠苗分化率基因型間差異最大。基因型愈傷組織誘導特性與綠苗、白苗的分化正相關,愈傷再生分化成綠苗或是白苗沒有相關性。篩選出綠苗產率高于1.0%的基因型22個,其中,SPLM2、衡96851、石4185、邯6172、河農6425五個基因型農藝性狀較好,綠苗產率依次為8.17%、5.44%、2.39%、2.00%、0.72%,可作為花培育種的骨干親本。
小麥;花藥培養力;基因型
單倍體是指含有配子體染色體數的個體,經自然或人工加倍后可獲得純合的二倍體植株,應用于育種中可縮短育種年限、提高育種效率。單倍體自然發生頻率較低,人工方法誘導是獲得單倍體的主要途徑[1]。花藥離體培養是通過人工途徑獲得小麥單倍體的主要途徑之一,在小麥新品種選育中發揮著重要作用。目前,國內外育種家利用花藥離體培養技術已選育出京花1號[2]、AC Andrew[3]、陜農28[4]、H6756[5]、花培8號[6]、生選6號[7]、寧春50號[8]、隴春31號[9]等一批小麥新品種并在生產上應用。
基因型依賴性在一定程度上限制了花藥離體培養技術在育種工作中優勢潛力的充分發揮[10-11]。研究表明,通過選擇高花藥培養力小麥基因型作為橋梁親本組配組合可以減輕育種實踐中一些配組綠苗率極低或根本不出綠苗的問題,從而提高小麥花培育種的效率[12-14]。一些研究單位開展了高花藥培養力小麥基因型的創制和篩選,例如趙林姝等[15-16]利用誘變及花藥培養方法創制出具有高綠苗再生能力的冬小麥基因型H307和SPLM2號等;韓曉峰等[17]通過愈傷組織誘導率的測定,認為石4185及新春9 號小麥品種花藥愈傷組織誘導率較高,是開展花藥離體培養的適宜基因型。我國冬小麥種植面積和總產量均占常年小麥總面積和總產的90%以上,其中,北方冬麥區(包括北部冬麥區和黃淮冬麥區)小麥面積及總產通常占全國冬麥總量的60%以上,為冬小麥主產區[18]。為減輕北部冬麥區花培育種工作中的基因型依賴問題,本研究對北方冬麥區部分育種單位育成品種(系)的花藥培養力性狀進行鑒定,篩選出適于該麥區種植的高花藥培養力性狀基因型,進一步根據農藝性狀及配合力分析篩選出優異花培育種親本,利用優異親本與生產上主栽品種配制組合并用于花培育種工作,以期提高花培育種的效率。
1 材料與方法
1.1 試驗材料
供試材料為74個適于北部及黃淮冬麥區種植的新品種(系)(表1),現均保存于中國農業科學院作物科學研究所。2011-2012年度將所有供試材料種植于中國農業科學院作物科學研究所試驗農場(北京),每個品種(系)種植1行,按常規方法進行田間管理。

表1 74個基因型的花藥培養力Table 1 Anther culture ability of seventy-four genotypes %
(續表1Continuedtable1)
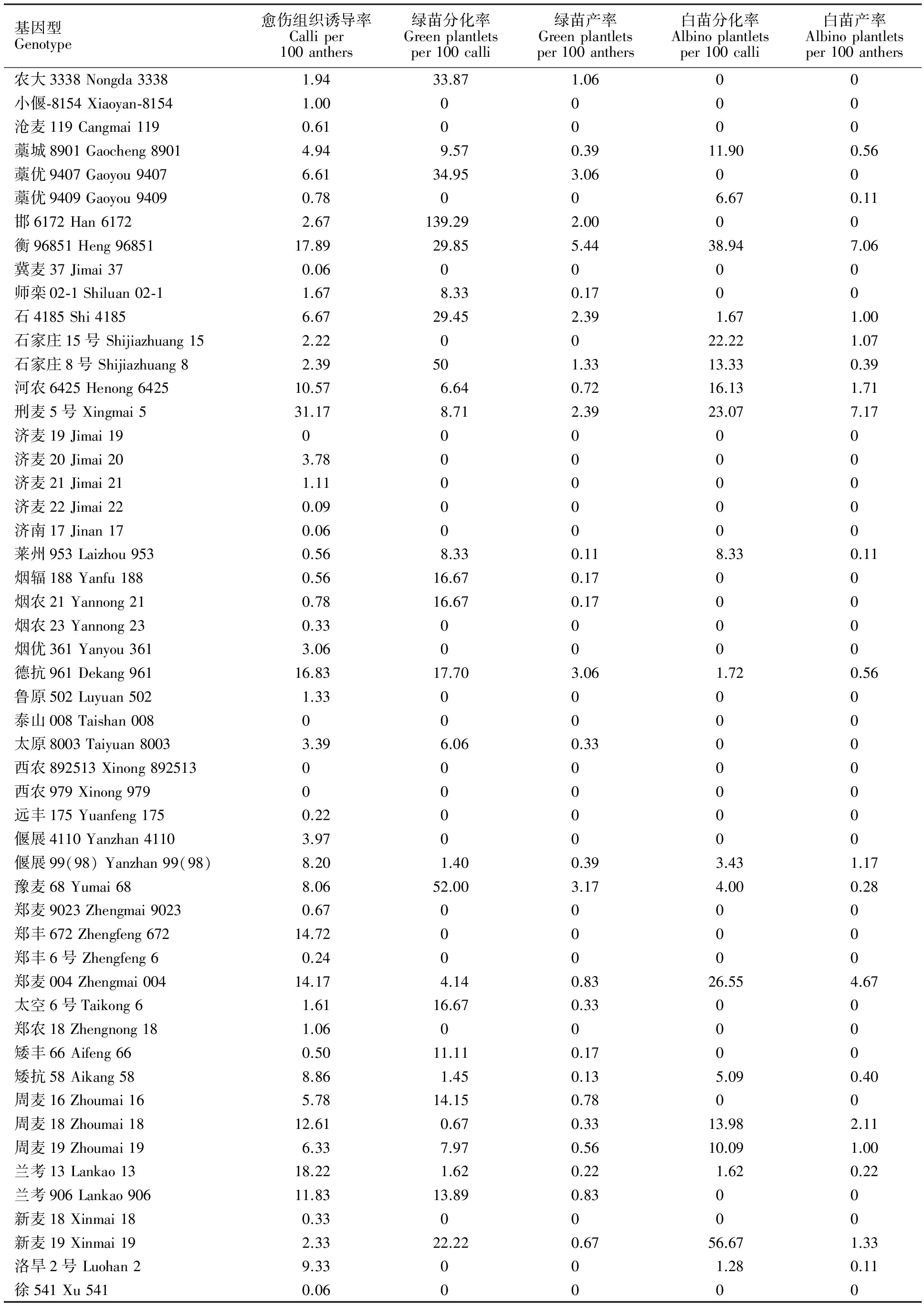
基因型Genotype愈傷組織誘導率Calliper100anthers綠苗分化率Greenplantletsper100calli綠苗產率Greenplantletsper100anthers白苗分化率Albinoplantletsper100calli白苗產率Albinoplantletsper100anthers農大3338Nongda33381.9433.871.0600小偃?8154Xiaoyan?81541.000000滄麥119Cangmai1190.610000藁城8901Gaocheng89014.949.570.3911.900.56藁優9407Gaoyou94076.6134.953.0600藁優9409Gaoyou94090.78006.670.11邯6172Han61722.67139.292.0000衡96851Heng9685117.8929.855.4438.947.06冀麥37Jimai370.060000師欒02?1Shiluan02?11.678.330.1700石4185Shi41856.6729.452.391.671.00石家莊15號Shijiazhuang152.220022.221.07石家莊8號Shijiazhuang82.39501.3313.330.39河農6425Henong642510.576.640.7216.131.71刑麥5號Xingmai531.178.712.3923.077.17濟麥19Jimai1900000濟麥20Jimai203.780000濟麥21Jimai211.110000濟麥22Jimai220.090000濟南17Jinan170.060000萊州953Laizhou9530.568.330.118.330.11煙輻188Yanfu1880.5616.670.1700煙農21Yannong210.7816.670.1700煙農23Yannong230.330000煙優361Yanyou3613.060000德抗961Dekang96116.8317.703.061.720.56魯原502Luyuan5021.330000泰山008Taishan00800000太原8003Taiyuan80033.396.060.3300西農892513Xinong89251300000西農979Xinong97900000遠豐175Yuanfeng1750.220000偃展4110Yanzhan41103.970000偃展99(98)Yanzhan99(98)8.201.400.393.431.17豫麥68Yumai688.0652.003.174.000.28鄭麥9023Zhengmai90230.670000鄭豐672Zhengfeng67214.720000鄭豐6號Zhengfeng60.240000鄭麥004Zhengmai00414.174.140.8326.554.67太空6號Taikong61.6116.670.3300鄭農18Zhengnong181.060000矮豐66Aifeng660.5011.110.1700矮抗58Aikang588.861.450.135.090.40周麥16Zhoumai165.7814.150.7800周麥18Zhoumai1812.610.670.3313.982.11周麥19Zhoumai196.337.970.5610.091.00蘭考13Lankao1318.221.620.221.620.22蘭考906Lankao90611.8313.890.8300新麥18Xinmai180.330000新麥19Xinmai192.3322.220.6756.671.33洛旱2號Luohan29.33001.280.11徐541Xu5410.060000
1.2 花藥培養
選取花粉粒發育至單核中晚期小麥幼穗,用潮濕的紗布包裹后置于4 ℃冰箱預處理2~3 d。接種前將穗子剝出,用0.1% HgCl2消毒8~10 min,超凈臺中無菌蒸餾水沖洗3~4次,用鑷子取出花藥置于誘導培養基上。每個基因型接種18個穗子,每穗選取中部小穗花藥進行接種,每3個穗子(100個花藥)接種1瓶,每重復200個花藥(2瓶),3次重復。
誘導培養采用C17培養基[19]+2.0 mg·L-12,4-D+0.5 mg·L-1KT+1.5 mg·L-1D-生物素+100.0 mg·L-1Casein Hydrolysate+90 g·L-1蔗糖+5 g·L-1瓊脂,pH 5.8。誘導培養采用暗培養,32±1 ℃處理3 d,之后27~29 ℃誘導愈傷。
當愈傷組織直徑長至0.5~1.0 mm時轉移到分化培養基上,分化培養采用MS培養基[20]+1.0 mg·L-1NAA+1.0 mg·L-1KT+30 g·L-1蔗糖+5 g·L-1瓊脂,pH 5.8。分化培養采用24~26 ℃光培養,每天光照14~16 h,光強2 000 lx。
1.3 花藥培養力性狀統計與數據分析
愈傷組織誘導率=(產生的愈傷組織塊數/接種花藥數)×100%;
綠苗分化率=(產生的綠苗數/轉分化愈傷數)×100%;
綠苗產率=(產生的綠苗數/接種花藥數)×100%;
白苗分化率=(產生的白苗數/轉分化愈傷數)×100%;
白苗產率=(產生的白苗數/接種花藥數)×100%。
利用EXCEL軟件繪制花藥培養力性狀頻次分布圖,利用SPSS統計分析軟件計算相關系數并進行顯著性測驗。
2 結果與分析
2.1 不同基因型愈傷組織誘導特性的差異
不同基因型間愈傷組織誘導率差異明顯,74個基因型的愈傷組織誘導率分布范圍為0~43.17%,平均數為7.7%,中位數為2.53%,其中70個基因型能脫分化出愈傷,但多數基因型愈傷組織誘導率偏低(表1、圖1A)。愈傷組織誘導率高于平均數的基因型有25個,其中,高于40%的基因型有2個(中優9701、中麥22),30%~40%之間的基因型有2個(SPLM2、刑麥5號),20%~30%之間的基因型有4個(H90265、H60329S、H90096、H90304),10%~20%之間的基因型有13個(具體見表1)。

圖1 74個基因型不同花藥培養力性狀的頻次分布
2.2不同基因型愈傷再生分化花粉植株特性的差異
不同基因型愈傷再生分化綠苗的特性差異明顯,74個基因型愈傷綠苗分化率分布范圍為0~139.29%,平均數為15.03%,中位數為4.63%,其中,42個基因型的愈傷能分化出綠苗,但多數基因型綠苗分化率偏低(表1、圖1B)。綠苗分化率高于平均數的基因型有24個,其中,高于100%的基因型有1個(邯6172),70%~100%之間的基因型有3個(H60148、H60329S、H261),60%~70%之間的基因型有1個(S6123),50%~60%之間的基因型有3個(H60279、豫麥68、石家莊8號),40%~50%之間的基因型有1個(H60017),15%~40%之間的基因型有15個(具體見表1)。
不同基因型愈傷再生分化白苗的特性差異明顯,74個基因型白苗分化率分布范圍為0~63.33%,平均數為6.91%,中位數為0,其中,33個基因型的愈傷再生出了白苗,但多數基因型白苗分化率偏低(表1、圖1D)。白苗分化率高于平均數的基因型有19個,其中,白苗分化率高于60%的基因型有1個(H6756),50%~60%之間的基因型有3個(新麥19、H90304、H90096),30%~40%之間的基因型有1個(衡96851),20%~30%之間的基因型有3個(鄭麥004、刑麥5號、石家莊15號),10%~20%之間基因型有9個(具體見表1)。
2.3不同基因型花藥經脫分化、分化過程再生花粉植株特性的差異
不同基因型花藥分化綠苗能力差異明顯,74個基因型綠苗產率分布范圍為0~20.83%,平均數為1.71%,中位數為0.17%,多數基因型綠苗產率偏低(表1、圖1C)。綠苗產率高于1%的基因型有20個,其中,綠苗產率高于20%的基因型有1個(H60329S),10%~20%之間的基因型有2個(H60148、H261),1.71%~10%之間的基因型有17個(具體見表1)。
74個基因型白苗產率分布范圍為0~7.17%,平均數為0.77%,中位數為0,多數基因型白苗產率偏低(表1、圖1E)。白苗產率高于平均數的基因型有19個,其中,白苗產率高于7%的基因型有2個(刑麥5號、衡96851),1%~5%之間的基因型有15個(具體見表1)。
2.4 花藥培養力性狀間的相關性
相關性分析結果(表2)表明,愈傷組織誘導率與綠苗分化率呈顯著正相關,與白苗分化率、綠苗產率及白苗產率呈極顯著正相關;綠苗分化率與綠苗產率呈極顯著正相關,與白苗分化率及白苗產率相關不顯著;綠苗產率與白苗產率呈顯著正相關,與白苗分化率相關不顯著;白苗分化率與白苗產率呈極顯著正相關。
2.5 花培育種骨干親本的農藝性狀
本研究通過對適于北部及黃淮冬麥區種植的74個基因型的花藥培養力性狀的鑒定,篩選出綠苗產率高于1.0%的基因型22個,通過對這些基因型的農藝性狀觀察,其中,SPLM2、衡96851、石4185、邯6172、河農6425共5個基因型農藝性狀較好(表3),可作為花培育種骨干親本。
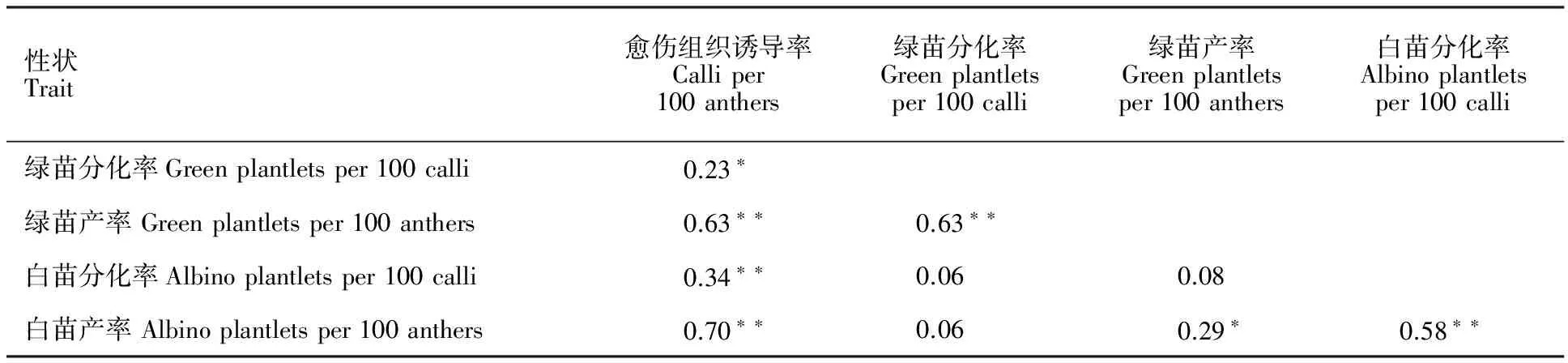
表2 5個花藥培養力性狀的相關系數Table 2 Correlation coefficients among five traits related to anther culture ability
*和**分別表示在0.05和0.01水平上相關顯著。
* and ** mean significant at 0.05 and 0.01 levels, respectively.

表3 5個基因型的農藝性狀Table 3 Agronomic traits of five genotypes
3 討 論
3.1小麥花藥培養力性狀的基因型依賴性及性狀間相關性
花藥離體培養再生花粉植株需經過二個階段,一是花藥內的花粉粒由配子體發育途徑轉變為孢子體發育途徑,脫分化形成愈傷組織或胚狀體,二是愈傷組織再分化出根和芽,最后形成花粉植株,或胚狀體直接再生出花粉植株。衡量小麥基因型花藥培養力的5個性狀指標中,愈傷組織誘導率表示的是花藥離體培養過程第一階段中花藥再生愈傷的能力;綠苗分化率和白苗分化率表示的是花藥離體培養過程第二階段中愈傷或胚狀體再生綠苗或白苗的能力;綠苗產率和白苗產率衡量的是花藥離體培養過程中第一階段和第二階段的綜合能力,即花藥經過脫分化愈傷(胚狀體)過程,最終再生出綠苗或白苗的能力。
研究表明,小麥花藥離體培養存在基因型依賴性,花藥離體培養的基因型依賴性是限制小麥花培技術廣泛應用的主要因素之一。韓曉峰等[17]對11個冬(春)小麥基因型花藥愈傷組織誘導特性的研究表明,愈傷組織誘導率差異顯著,基因型間分布范圍為1.9%~51.9%;王 煒等[21]對86 份甘肅主栽小麥和骨干親本的愈傷誘導及花粉苗分化情況的研究表明,愈傷組織誘導率、綠苗分化率、綠苗產率及白苗分化率基因型間差異顯著,其中,愈傷組織誘導率分布范圍為0~88.67%,綠苗分化率分布范圍為 0~143.74%,綠苗產率分布范圍為0~56.67%,白苗分化率分布范圍為0~60.92%。本研究對適于北方冬麥區種植的74個基因型的花藥培養力性狀鑒定結果表明,愈傷組織誘導率、綠苗分化率、綠苗產率、白苗分化率及白苗產率5個花培性狀基因型間均差異明顯,與前人研究結果相同。其中,基因型間差異最大的性狀是綠苗分化率,其次是白苗分化率、愈傷組織誘導率、綠苗產率、白苗產率。本研究結果與王 煒等[21]研究結果基本一致,基因型間差異最大的花培力性狀均為綠苗分化率,基因型間差異均超過100個百分點;由于綠苗產率和白苗產率衡量的是基因型由花藥脫分化愈傷(胚狀體)及再分化花粉植株的綜合能力,因此在所研究花培力性狀中差異相對較小。
王 煒等[21]對86份甘肅主栽小麥和骨干親本花藥離體培養的愈傷誘導及綠苗分化情況的相關分析研究表明,愈傷組織誘導率和綠苗分化率沒有相關性。本研究對適于北部及黃淮冬麥區種植的74個基因型的花藥培養力性狀相關性分析表明,愈傷組織誘導率和綠苗分化率呈正相關,此結果與王 煒等[21]研究結果不同,可能是因為所研究基因型不同導致,后續還有待利用更多的基因型進行驗證。
3.2 北方冬麥區小麥花培育種骨干親本的利用
花藥離體培養的最終目的是產生盡可能多的綠苗,不產生或盡可能的減少白苗的產生,因此,綠苗產率是評價小麥花藥培養力的重要指標。本實驗室已利用篩選出的高綠苗率且農藝性狀較好的5個花培育種骨干親本配制了一批組合,特別是在大穗、抗倒、抗病的自育品系SPLM2配制的組合后代中篩選出了一批苗頭品系,其中,航麥2566、航麥501、航麥287共3個新品系正在參加國家或省級生產試驗,航麥806等4個新品系正在參加省級第二年或第一年區試。
[1]GERMANA M A.Anther culture for haploid and doubled haploid production [J].PlantCellTissueandOrganCulture,2011,104(3):283.
[2] 胡道芬,湯云蓮,袁振東,等.冬小麥花粉孢子體的誘導及京花1號的育成[J].中國農業科學,1983(1):29.
HU D F,TANG Y L,YUAN Z D,etal.The induction of pollen sporophyte of winter wheat and the development of the new variety Jinghua 1 [J].ScientiaAgriculturaSinica,1983(1):29.
[3]SADASIVAIAH R S,PERKOVIC S M,PEARSON D C,etal.Registration of 'Deloris' wheat [J].CropScience,2004,44:695.
[4] 王成社,李景琦,鄒淑芳,等.小麥新品種陜農28的選育與花培育種技術的改良[J].西北農林科技大學學報(自然科學版),2002,30(40):21.
WANG C S,LI J Q,ZOU S F,etal.Discussion on the improvement of haploid breeding from selecting new varieties of Shaannong 28 [J].JournalofNorthwestAgricultureandForestryUniversity(NaturalScienceEdition),2002,30(40):21.
[5]LIU L X,ZHAO L S,GUO H J,etal.A salt tolerant mutant wheat cultivar ‘H6756’ [J].PlantMutationReports,2007(1):50.
[6]KANG M H,HAI Y,HUANG B Y,etal.Breeding of newly licensed wheat variety Huapei 8 and improved breeding strategy by anther culture [J].AfricanJournalofBiotechnology,2011,10:19701.
[7] 姚金保,陸維忠.小麥新品種生選6號的豐產穩產性和赤霉病抗性分析[J].麥類作物學報,2012,32(4):793.
YAO J B,LU W Z.Analysis of yield potential,yield stability and resistance to Fusarium head blight of new wheat cultivar Shengxuan 6 [J].JournalofTriticeaeCrops,2012,32(4):793.
[8] 魏亦勤,葉興國,李紅霞,等.高產優質春小麥新品種寧春50號[J].麥類作物學報,2012,32(2):38.
WEI Y Q,YE X G,LI H X,etal.A high yield and good quality spring wheat variety Ningchun 50 [J].JournalofTriticeaeCrops,2012,32(2):38.
[9] 王 煒,楊隨莊,葉春雷,等.豐產優質花培春小麥新品種-隴春31號[J].麥類作物學報,2014,34(3):432.
WANG W,YANG S Z,YE C L,etal.A high yield and good quality spring wheat variety Longchun 31 of anther culture method [J].JournalofTriticeaeCrops,2014,34(3):432.
[10]BROUGHTON S.Ovary co-culture improves embryo and green plant production in anther culture of Australian spring wheat(TriticumaestivumL.) [J].PlantCellTissueandOrganCulture,2008,95:190.
[11]LANTOS C,WEYEN J,ORSINI J M,etal.Efficient application ofinvitroanther culture for different European winter wheat(TriticumaestivumL.) breeding programmes [J].PlantBreeding,2013,132:149.
[12]TUVESSON S,LJUNGBERG A,JOHANSSON N,etal.Large-scale production of wheat and triticale double haploids through the use of a single-anther culture method [J].PlantBreeding,2000,119:458.
[13] 張艷敏,郭北海,李洪杰,等.小麥花藥培養的基因型差異與雜交組合配制[J].華北農學報,2002,17(2):18.
ZHANG Y M,GUO B H,LI H J,etal.Genotype difference in wheat anther culture response and selection of cross combination [J].ActaAgriculturaeBoreali-Sinica,2002,17(2):18.
[14]YILDIRIM M,BAHAR B,GENC I.Reciprocal effects in anther cultures of wheat hybrids [J].BiologiaPlantarum,2008,52(4):781.
[15]ZHAO L S,LIU L X,WANG J,etal.Development of a new wheat germplasm with high anther culture ability by using a combination of gamma-ray irradiation and anther culture [J].JournaloftheScienceofFoodandAgriculture,2015,95:122.
[16] 趙林姝,劉錄祥,郭會君,等.三個高花藥培養力小麥材料培養力性狀的配合力分析[J].麥類作物學報,2012,32(3):427.
ZHAO L S,LIU L X,GUO H J,etal.Combining ability analysis on anther culture traits of three wheat genotypes with high regeneration ability [J].JournalofTriticeaeCrops,2012,32(3):427.
[17] 韓曉峰,陶麗莉,殷桂香,等.基因型和環境條件對小麥花藥培養效果的影響[J].作物學報,2010,36(7):1209.
HAN X F,TAO L L,YIN G X,etal.Effect of genotype and growing environment on anther culture in wheat [J].ScientiaAgriculturaSinica,2010,36(7):1209.
[18] 趙廣才.中國小麥種植區劃研究(一)[J].麥類作物學報,2010,30(5):888.
ZHAO G C.Study on Chinese wheat planting regionalization(Ⅰ) [J].JournalofTriticeaeCrops,2010,30(5):888.
[19] 王 培,陳玉蓉.C17培養基在花藥培養中應用的研究[J].植物學報,1986,28(1):38.
WANG P,CHEN Y R.A study on the application of C17medium for anther culture [J].ActaBotanicaSinica,1986,28(1):38.
[20]MURASHIGE T,SKOOG F.A revised medium for rapid growth and bioassays with tobacco tissue cultures [J].PhysiologiaPlantarum,1962,15:478.
[21] 王 煒,陳 琛,葉春雷,等.甘肅主栽小麥品種及骨干親本花藥培養特性評價及分析[J].核農學報,2016,30(6):1061.
WANG W,CHEN S,YE C L,etal.Evaluation and analysis of anther culture characteristics in Gansu wheat cultivars and backbone parents [J].JournalofNuclearAgriculturalSciences,2016,30(6):1061.
ScreeningofWinterWheatGermplasmswithHighAntherCultureAbility
ZHAOLinshu,LIULuxiang,GUJiayu,XIEYongdun,GUOHuijun,ZHAOShirong,LIJunhui,XIONGHongchun,
(Institute of Crop Science, Chinese Academy of Agricultural Sciences,Beijing 100081, China)
In order to screen the genotype with good anther culture response and agronomic traits, and decrease the genotype dependence, we characterized five anther-culture associated traits for 74 winter wheat varieties (lines) and conducted correlation analysis on these traits. The results showed that the variation range of the proportion of calli per 100 anthers, green plantlets per 100 calli, green plantlets per 100 anthers, albino plantlets per 100 calli and albino plantlets per 100 anthers was 0-43.17%, 0-139.29%, 0-20.83%, 0-63.33%, and 0-7.17%, respectively. All the five traits showed great genotype dependence, among which, the proportion of green plantlets per 100 calli displayed greatest genotype dependence. Correlation analysis results indicated that there was positive correlation between callus induction capacity of each genotype and green- and albino-seedlings differentiation. It did not show any correlation whether callus would differentiate to green seedlings or albino seedlings. In this study, the proportion of green plantlets per 100 anthers of 22 genotypes screened was higher than 1%. Five of these 22 genotypes, such as SPLM2, Heng 96851, Shi 4185, Han 6172, and Henong 6425, are suggested to serve as backbone parents for anther culture based breeding approach, as they have good agronomic traits.
Wheat; Anther culture ability; Genotype
時間:2017-10-11
網絡出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20171011.1601.006.html
2017-03-10
2017-07-31
“十三五”國家重點研發計劃項目(2016YFD0102101);中國農業科學院作物科學研究所中央級公益性科研院所基本科研業務費專項
E-mail:zhaolinshu@caas.cn
劉錄祥(E-mail:liuluxiang@caas.cn)
S512.1;S330
A
1009-1041(2017)10-1294-07
