黑龍江省水稻品種抗性與稻瘟病病菌致病性年際變化趨勢分析
2017-11-30 21:13:08馬軍韜張國民張麗艷鄧凌韋王永力王英宮秀杰任洋
江蘇農業科學 2017年20期
馬軍韜+張國民+張麗艷+鄧凌韋+王永力+王英+宮秀杰+任洋

摘要:為分析黑龍江省水稻品種抗性及稻瘟病病菌致病性在年際間的波動及變異趨勢,以8個水稻品種及24個抗瘟基因為試驗材料,以180株稻瘟病病菌為選擇壓力,應用噴霧接種方式進行試驗。結果表明,以2009—2011年的病菌為選擇壓力,水稻品種的抗性頻率分別介于43.33%~93.33%、28.33%~73.33%、25.00%~78.33%之間,龍盾104抗性較好;聯合抗病性方式下,各水稻組合的聯合抗病性系數平均值分別為0.57、0.33、0.35,綏粳4與龍盾104組合抗性較好。以2009—2011年的病菌為選擇壓力,各基因的抗性頻率分別介于6.67%~95.00%、5.00%~8667%、6.67%~91.67%之間,Pi-z5、Pi-9基因抗性較好;聚合Pi-z5與Pi-9基因條件下,品種抗性頻率平均值分別為10000%、90.83%、96.46%。2009—2011年病菌的致病性相似系數分別介于0.35~0.97、0.19~1.00、0.20~1.00之間,致病率分別介于8.33%~87.50%、8.33%~95.83%、12.50%~95.83%之間,無毒基因Pi-9和Pi-z5出現的頻率較高且相對穩定。
關鍵詞:黑龍江省;水稻品種;抗瘟基因;稻瘟病菌;致病性;變化趨勢
中圖分類號: S435.111.4+1 文獻標志碼: A 文章編號:1002-1302(2017)20-0109-05
黑龍江省是我國水稻生產大省,同時也是稻瘟病為害的重災區,常年減產在5%以上[1-2]。目前,當地生產上對該病害的控制以化學防控為主,農業防控為輔,但前者污染環境,后者技術水平要求高,效果均不理想。進一步分析發現,水稻種質由于自身抗瘟基因的存在,對稻瘟病具有較高的抗性權重,應用潛力較大。但是,稻瘟病病菌受內在遺傳機制及外界環境影響,致病性存在分化,對不同類型種質侵染力差別顯著。因此,要想實現水稻種質的生態控病,首要條件是明確稻瘟病病菌致病性的分化情況,獲得水稻種質抗性信息,然后通過合理布局或抗性育種等生態方式實現病害高效控制。
目前,國內外相關研究較多。馬輝剛等以江西省67個稻瘟病病菌菌株為選擇壓力,對35個水稻品種的抗性進行了評價,明確了兩優培九的高抗源地位;應用聯合抗病性方式,明確了兩優培九與汕優晚3的優勢組合地位[3]。張國民等以12個日本鑒別品種和24個水稻單基因系為鑒別體系,對黑龍江省178個稻瘟病病菌菌株的致病性進行了分析,將其劃分為104個生理小種,明確了Pi-9的高抗源地位,同時獲得了墾稻10、上育397等品種的抗性改良信息[4]。楊秀娟等以41個已知抗性基因的水稻品種為鑒別體系,對福建省87個稻瘟病病菌菌株的致病性進行分析,明確了無毒基因AvrPid-2、AvrPik的高頻率地位[5]。此外,張亞玲等也相繼進行了類似研究[6-8],對科研及生產起到了一定的促進作用。但是,多數研究由于缺乏種質抗性或病菌致病性在年際間變化及波動趨勢方面的分析,從而難以明確種質抗性或病菌致病性的未來發展趨勢,不利于抗性布局及育種策略的適時調整,有待改進。
本研究以黑龍江省主栽水稻品種和單基因系作為水稻種質,以連續3年的稻瘟病病菌菌株作為選擇壓力,在獲得水稻種質抗性及稻瘟病病菌致病性準確的信息前提下,明確其年際間變異及波動趨勢,為相關的持續研究提供基礎信息,同時,應用聯合抗病性分析及基因聚合分析的方式為抗性布局及抗瘟育種工作提供優化方案,從而生態控制病害。
1 材料與方法
1.1 試驗材料
8個水稻品種分別為松粳9、松粳6、龍稻7、墾稻12、龍盾104、綏粳4、龍稻4、空育131,均由黑龍江省農業科學院耕作栽培研究所提供。
24個水稻單基因系各含有1個已知抗瘟基因,感病對照品種為麗江新團黑谷,不含任何抗瘟基因,均由黑龍江省農業科學院耕作栽培研究所提供。
稻瘟病病菌:總計180株單孢菌株,來源于黑龍江省第一、第二、第三積溫帶的水稻品種。其中,2009年在24個水稻品種上分離獲得60株單孢菌株,2010年在21個水稻品種上分離獲得60株單孢菌株,2011年在22個水稻品種上分離獲得60株單孢菌株,均由黑龍江省農業科學院耕作栽培研究所分離并保存。
培養基:菌株分離與活化使用燕麥片番茄汁培養基,配方為30 g/L燕麥片、150 mL/L番茄汁、17~20 g/L 瓊脂;菌株產孢使用高粱粒培養基,將約200 g高粱粒煮熟后置于三角瓶中,121 ℃濕熱滅菌30 min后備用。
1.2 試驗方法
1.2.1 稻瘟病病菌單孢子的分離及培養 將病葉用無菌水清洗后,浸泡4~6 h,放入恒溫箱培養至產生霉層。鏡檢挑取單個分生孢子點接于燕麥片番茄汁培養基中,25 ℃培養3 d,將菌絲塊挑入裝有高粱粒培養基的三角瓶中,25 ℃培養20~25 d,待高粱粒表面長滿菌絲后,無菌水洗脫菌絲,將高粱粒平鋪于已滅菌鐵托盤中,蓋上無菌紗布,待高粱粒表面形成大量分生孢子后,用含0.025%吐溫-20的自來水洗脫孢子,采用血球計數板計數,將孢子懸浮液濃度調至5×105 個/mL,供接種使用[9]。
1.2.2 水稻品種抗性分析方法 水稻品種抗性分析通過幼苗3葉1心期噴霧接種方式進行。幼苗以草碳土、黑土為基質,用育秧盤育苗,個體保苗15株,3次重復。待幼苗長至3葉1心期時,接種孢子懸浮液,劑量為100 mL/盤,然后在 26 ℃、相對濕度為100%條件下暗培養30 h,光照培養96~144 h,待發病完全后按表1中的標準調查發病情況[10]。調查后,按Hayashi等的標準[11],將0~3級歸為抗病反應型,4~5級歸為感病反應型。各重復間接種結果若抗、感病表現不一致,則按感病表現進行統計。
用抗性頻率表示水稻品種抗性強弱,抗性頻率=(非致病病菌菌株數/總菌株數)×100%,用聯合抗病性[3,12]評價水稻品種搭配種植后抗性效果,它包括聯合抗病性系數和聯合致病性系數2個指標,以水稻品種組合既同時高抗稻瘟病病菌(聯合抗病性系數最大)又不會同時被稻瘟病病菌侵染(聯合致病性系數最小)為優劣評價標準,聯合抗病性系數=水稻品種組合共同非致病病菌菌株數/總菌株數;聯合致病性系數=水稻品種組合共同致病病菌菌株數/總菌株數。endprint
1.2.3 水稻抗瘟基因抗性分析方法 水稻抗瘟基因抗性分析通過幼苗3葉1心期噴霧接種方式進行,具體方法同“1.2.2”節。用聚合后的抗性頻率表示抗瘟基因與水稻品種聚合后可能獲得的最佳抗性,通過常規雜交技術完成,聚合后抗性頻率=[(總菌株數-聚合后共同致病的菌株數)/總菌株數]×100%。
1.2.4 稻瘟病病菌致病性分析方法 稻瘟病病菌致病性分析通過幼苗3葉1心期噴霧接種方式進行,具體方法同“1.2.2”節。
用致病率表示稻瘟病病菌對水稻品種或抗瘟基因的致病力強弱,致病率=(致病菌株數/總菌株數)×100%。
用稻瘟病病菌致病性聚類分析方式評價菌株間致病性的分化情況,菌株間致病性相似系數差值越大,表示其致病性分化越劇烈。將水稻品種與稻瘟病病菌的反應型作為性狀,抗病反應型記為1,感病反應型記為0,構成二態性狀。相似系數=1-相異系數。
1.3 數據分析
應用Excel 2010對數據進行統計與計算,利用DPS v7.05分析軟件,以不加權算術平均法(UMPGA)完成分析。
2 結果與分析
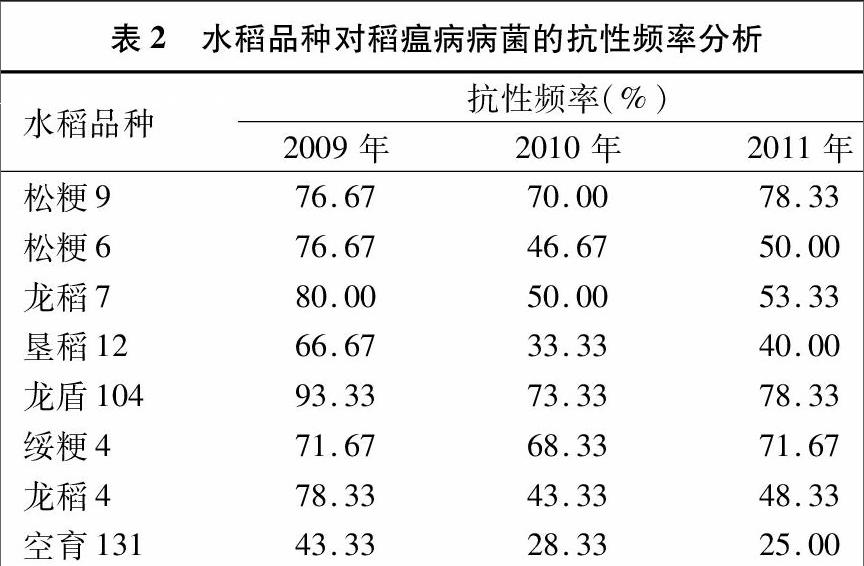
2.1 水稻品種對不同年際稻瘟病病菌抗性的分析
如表2所示,各水稻品種的抗性在穩定的前提下,年際間存在較大波動。以2009—2011年的病菌為選擇壓力,水稻品種的抗性頻率分別介于43.33%~93.33%、28.33%~7333%、25.00%~78.33%之間,平均值分別為73.33%、5167%、5562%。其中,空育131的抗性在年際間呈持續下降的趨勢,其他7個品種的抗性在年際間均呈先下降后上升的趨勢,龍盾104與空育131一直分別為各年份抗性最好與最差的水稻品種。2011年與2009年相比,松粳9的抗性略有上升,上升了2.17%,其他7個品種的抗性均呈持平或下降趨勢,龍稻4的抗性降幅最大,為38.30%。
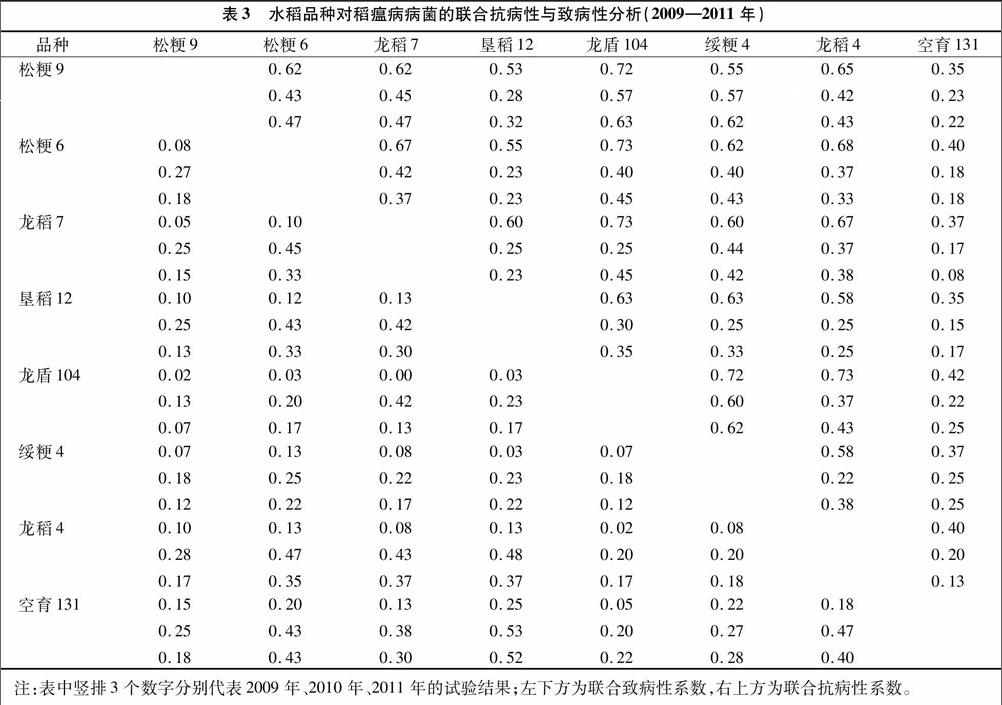
由表3可知,以2009—2011年病菌為選擇壓力,各水稻組合的聯合抗病性系數均值分別為0.57、0.33、0.35,聯合致病性系數均值分別為0.10、0.31、0.24。其中,各年際間相對較好的組合分別為龍稻7與龍盾104、龍稻4與龍盾104、綏粳4與龍盾104、松粳9與龍盾104。綜合比較可知,綏粳4與龍盾104為聯合抗病性較好、年際間波動較小的水稻組合,可以廣泛推廣應用。
2.2 抗瘟基因對不同年際稻瘟病病菌抗性分析
由表4可知,24個抗瘟基因的抗性在年際間存在較大的波動,以2009—2011年的病菌為選擇壓力,抗瘟基因的抗性頻率分別介于6.67%~95.00%、5.00%~86.67%、6.67%~9167%之間,平均值分別為44.17%、30.42%、3049%。其中,2009—2011年抗性頻率≥70%的基因分別有5、3、2個,依次為Pi-9、Pi-z5、Pi-ta2、Pi-12、Pi-20,Pi-zt、Pi-z5、Pi-9,Pi-z5、Pi-9;2009—2011年抗性頻率≤30%的基因分別有11、14、14個,Pi-19、Pi-a、Pi-t分別為各年抗性最差的基因。綜合評價可知,Pi-z5、Pi-9基因的抗性較好,年際間表現穩定,為優質抗源。依據抗性的變化趨勢,可將24個抗瘟基因劃分為5種類型:Pi-kp等6個基因的抗性呈持續下降趨勢;Pi-k等3個基因的抗性呈持續上升趨勢;Pi-zt、Pi-t基因的抗性呈先上升后下降趨勢;Pi-ta2基因的抗性呈先下降后持平趨勢;Pi-a等12個基因的抗性呈先下降后上升趨勢。這間接說明了黑龍江省稻瘟病病菌的致病性變異趨勢復雜。此外,2011年與2009年相比,Pi-k基因的抗性升幅最大,上升了109.11%,Pi-ta2基因的抗性降幅最大,下降了81.63%。
依據抗性較好且相對穩定的原則,選取Pi-z5、Pi-9基因通過基因聚合方式對水稻品種進行抗性改良分析。由表5可知,聚合Pi-z5與Pi-9基因組合前提下,水稻品種的抗性提升效應明顯。以2009—2011年的病菌為選擇壓力,各品種的聚合后抗性頻率平均值分別為100.00%、90.83%、9646%,其平均值分別比聚合前提高26.67%、39.16%、4084%。聚合單個基因前提下,以2009—2011年的病菌為選擇壓力,各年份抗性聚合的首選基因分別是Pi-9、Pi-z5、Pi-z5,龍盾104、龍盾104、綏粳4分別為各年份聚合后抗性頻率較高的品種,依次為100.00%、95.00%、98.33%。此外,空育131一直是聚合后抗性頻率提升幅度最大的品種。
2.3 不同年際稻瘟病病菌致病性分析
黑龍江省稻瘟病病菌致病性在年際間存在波動,從病菌致病性分化角度分析,2009—2011年病菌的致病性相似系數分別介于0.35~0.97、0.19~1.00、0.20~1.00之間,其差值分別為0.62、0.81、0.80。由圖1可知,以相似系數0.36為閾值,2009—2011年病菌分別被劃分為2、10、3個致病類群;以相似系數0.51為閾值,病菌分別被劃分為6、10、9個致病類群;以更高的相似系數為閾值,不同年際的病菌將被劃分為更多的致病類群。從病菌致病力角度分析,2009—2011年,病菌對24個抗瘟基因的致病率分別介于8.33%~87.50%、833%~95.83%、12.50%~95.83%之間,平均值分別為5583%、69.58%、69.51%。此外,2010、2011年開始有相同致病型的菌株群出現,致病率平均值分別為9107%、8472%。從病菌含有無毒基因角度分析,與24個抗瘟基因相對應的無毒基因全部出現,但年際間出現頻率不同,2009—2011年單個菌株含有無毒基因的平均數量依次為10.60、730、732個。綜合比較,無毒基因Pi-9和APi-z5出現的頻率較高且相對穩定,分別占當年總出現頻率的8.96%、959%、10.25%與7.70%、10.50%、12.53%。endprint
3 討論
黑龍江省為粳稻主產省,本研究選用的水稻品種在其域內累計推廣面積巨大,后代種質分布廣泛;水稻單基因系品種具有粳稻血緣,常規雜交結實率高、后代種質穩定快;稻瘟病病菌采自當地主產區,數量較多,具備年際間準確信息。因此,試驗靶標代表性強、可操作性好、應用價值大,在為理論研究奠定基礎的同時可通過生態手段解決現實難題。
從水稻品種抗性角度分析,存在2種波動趨勢:空育131的抗性呈逐年下降的趨勢;其他品種的抗性呈先急劇下降后略有上升的趨勢,除松粳9外,其他多數品種的抗性難以恢復到2009年的水平。從品種搭配后的聯合抗病性分析,同樣呈先下降后上升的趨勢,這是因為搭配種植的基礎源自水稻品種的自身抗性。除綏粳4與龍盾104等個別組合外,絕大部分組合的聯合抗病性系數偏低,聯合致病性系數偏高,不適宜推廣應用。馬軍韜等曾以2006年黑龍江省的稻瘟病病菌為選擇壓力,對13個水稻品種的抗性及聯合抗病性進行了分析,其抗性頻率介于47.19%~93.82%之間,平均值為7537%,聯合抗病性系數、致病性系數平均值分別為0.55、0.15[13],本研究2009年的結果與之基本持平,明顯高于2010年和2011年的研究結果。雖然兩者選用的水稻品種和病菌存在一定的差別,仍可從側面說明病菌的致病力已發生一定程度的改變。此外,水稻品種搭配后,自身抗性均強的品種共有抗病性較高,自身抗性均差的品種共有感病風險也較高,說明供試品種間抗性相似度較高,從側面說明其抗性遺傳基礎相對狹窄。水稻品種的抗性主要源自本身含有的抗瘟基因,一般可穩定遺傳,對相同類型病菌的抵抗能力基本無變化,其抗性之所以有波動,主要是病菌致病力及致病性在年際間發生改變引起的。
從抗瘟基因抗性角度分析,波動趨勢多樣,主體呈急劇下降又略有回升的趨勢,僅少數基因抗性較好且相對穩定。張國民等以2006年黑龍江省的稻瘟病病菌為選擇壓力,對24個抗瘟基因的抗性進行了分析,發現其抗性頻率介于3.93%~9775%之間,平均值為43.48%,Pi-9、Pi-z5、Pi-ta2、Pi-12、Pi-20和Pi-5共6個基因抗性頻率超過70%[4],本研究中2009年的結果與之基本持平,明顯高于2010年和2011年的研究結果。就抗性較高的優質基因而言,Pi-9、Pi-z5基因抗性較好且持續穩定,可以廣泛推廣應用;Pi-12、Pi-20基因抗性已不再突出,但仍有一定抗性,可以有限度地推廣應用;Pi-ta2、Pi-12基因抗性已急劇下降,應限制應用。上述結論再次說明,病菌的致病力已發生一定程度的改變。從基因聚合角度分析,綏粳4和松粳9通過雜交手段完成聚合后,抗性改良效果明顯,后代種質可以大力推廣;空育131基因聚合后抗性升幅最大,后代種質可以謹慎推廣。雷財林等曾分析了7個抗瘟基因對黑龍江省12個水稻品種的抗性改良效果,認為Pi-9基因對品種的抗性具有極強的改良效果[14],如能進一步導入Pi-z5、Pi-12基因,各品種對稻瘟病可表現免疫,與本研究的結論具有較強的相似度。目前,國內外共發現約101個各類抗瘟基因[15-17],遠多于本研究的基因群體,可進一步引進Pi-gm、Pi-40等優質抗源以再次提升當地品種的抗性。
從稻瘟病病菌致病性角度分析,病菌的致病率平均值及致病性相似系數差值均呈先急劇上升后略有下降的趨勢,說明病菌的致病力及致病性分化均呈先急劇上升后略有下降的趨勢。關于病菌致病性變異,可能主要有2個方面原因:(1)內在遺傳機制影響。病菌受自身異核現象、突變和準性重組等細胞學機制影響,菌絲融合現象普遍,田間自然突變率高于10%,完成致病性變異。(2)外在環境條件影響。隨著黑龍江省水稻種植面積的增加,很多未經審定的低抗性品系開始占有一定的種植比例,受互作關系影響,加劇了病菌致病力及致病性分化的復雜性與嚴重程度。2006、2007年黑龍江省稻瘟病連續大發生,產量損失巨大。在隨后的2008、2009年,天氣條件不利于發病,農民防控措施得當,只個別地區零星發病。自2010年以來,稻瘟病沒有再次大發生,防控措施開始松懈,病菌定殖、擴繁能力增強,致病力隨之增強;隨著種植品種持續增加及區域間引種日趨頻繁,病菌致病性分化總體呈加劇趨勢是可能的。馬軍韜等分析了2009—2010年哈爾濱市稻瘟病病菌變異情況后認為,當地病菌的致病力增強、致病性分化加劇[18],本研究的結論與之一致。此外,以稻瘟病病菌含有的無毒基因為現實依據,以Flor的“基因對基因”假說[19]為理論基礎,可推導出品種含有抗瘟基因的情況,為抗性合理布局提供理論借鑒。結果顯示,龍稻4和龍稻7可能含有Pi-sh基因,墾稻12可能含有Pi-kS基因,但上述結論僅為推測,還須結合標記技術進行驗證。
參考文獻:
[1]宋成艷. 黑龍江省水稻新品種(系)抗稻瘟病性鑒定及利用[J]. 植物保護,2011,37(4):142-145.
[2]支庚銀,張國民,雷材林,等. 黑龍江省2007年水稻稻瘟病生產調研及建議[J]. 黑龍江農業科學,2010(4):68-70.
[3]馬輝剛,曹九龍,胡水秀,等. 水稻品種對稻瘟病的抗性分析和利用評價[J]. 江西農業大學學報,2006,28(2):213-216.
[4]張國民,馬軍韜,肖佳雷,等. 已知抗瘟基因在黑龍江省寒地稻區的評價與利用[J]. 植物病理學報,2011,41(1):72-79.
[5]楊秀娟,阮宏椿,杜宜新,等. 福建省稻瘟病菌致病性及其無毒基因分析[J]. 植物保護學報,2007,34(4):337-342.
[6]張亞玲,王寶玉,臺蓮梅,等. 黑龍江省稻瘟病菌生理小種對水稻品種致病性分析[J]. 中國植保導刊,2014,34(2):22-26.
[7]趙正洪,周 政,吳偉懷,等. 湖南省稻瘟病菌生理小種的組成及其致病性[J]. 湖南農業大學學報(自然科學版),2014,40(2):173-177.endprint
[8]王世維,鄭文靜,趙家銘,等. 遼寧省稻瘟病菌無毒基因型鑒定及分析[J]. 中國農業科學,2014,47(3):462-472.
[9]周江鴻,王久林,蔣琬如,等. 我國稻瘟病菌毒力基因的組成及其地理分布[J]. 作物學報,2003,29(5):646-651.
[10]Mackill D J. Inheritance of blast resistance in near-isogenic lines of rice[J]. Phytopathology,1992,82(7):746-749.
[11]Hayashi N,Kobayashi N,Cruz C,et al. Protocols for the sampling of the diseases specimens and evaluation of blast disease in rice[J]. JIRCAS Working Report,2009,63:17-33.
[12]袁 潔,楊學輝,何海永. 部分水稻品種對稻瘟病菌群體的抗病性分析[J]. 種子,2006,25(8):14-16.
[13]馬軍韜,張國民,辛愛華,等. 黑龍江省水稻品種對稻瘟病的抗性分析及評價利用[J]. 中國農學通報,2008,24(2):332-334.
[14]雷財林,張國民,程治軍,等. 黑龍江省稻瘟病菌生理小種毒力基因分析與抗病育種策略[J]. 作物學報,2011,37(1):18-27.
[15]Wang Y,Wang D,Deng X,et al. Molecular mapping of the blast resistance genes Pi2-1 and Pi51(t) in the durably resistant rice ‘Tianjingyeshengdao[J]. Phytopathology,2012,102(8):779-786.
[16]Jiang H C,Feng Y T,Bao L,et al. Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding[J]. Molecular Breeding,2012,30(4):1679-1688.
[17]Xiao W M,Yang Q Y,Sun D Y,et al. Identification of three major R genes responsible for broad-spectrum blast resistance in an indica rice accession[J]. Molecular Breeding,2015,35(1):1-11.
[18]馬軍韜,張國民,辛愛華,等. 哈爾濱地區抗瘟基因抗性分析及水稻品種抗性評價與利用[J]. 植物保護學報,2015,42(2):160-168.
[19]Flor H H. Current status of the gene for gene concept[J]. Annual Review of Phytopathology,1971,9(1):275-296.endprint
