棉花光合基因GhRCAα啟動子的克隆及活性分析
2017-12-12 09:03:16晁毛妮張志勇張金寶溫青玉郭書磊馬亮王清連
華北農學報 2017年1期
關鍵詞:植物
晁毛妮,張志勇,張金寶,溫青玉,郭書磊,馬亮,王清連
(1.河南科技學院 生命科技學院, 現代生物育種河南省協同創新中心,河南 新鄉 453003;2.河南省農業科學院,河南 鄭州 450002)
通過調控植物的光合作用來增加植物的產量,是繼通過傳統育種技術、栽培措施及農業實踐等方法后,實現植物產量增加的一個新的重要的方向[1]。在大多數植物中,卡爾文循環在植物碳代謝中占據重要位置,調控這個途徑對于提高植物的光合效率和產量意義重大。
Rubisco作為卡爾文循環中的關鍵酶,不僅可以催化2個具有競爭性的反應,同時它也是一個具有較低周轉次數的低效酶,它的有效活化需要Rubisco活化酶(RCA)的參與[2]。因此,對RCA基因的研究已成為近年來提高植物產量的一個新的重要目標。通過調控RCA基因的表達來增加Rubisco的活性,在改善植物光合效率和提高植物產量方面具有很大的潛力[3-4]。許多研究發現,RCA基因的表達受光、植物激素、組織特異性及晝夜模式的調控[5-8]。與RCA基因具有組織特異性和受光調控的轉錄模式一致,菠菜[9]、擬南芥[10]、馬鈴薯[11]和水稻[12]的RCA基因啟動子在高等植物的綠色組織及光誘導下具有活性。RCA基因啟動子的光誘導和組織特異性在應對葉食害蟲或葉片抗病植物基因工程方面具有特定的優勢。例如,在馬鈴薯基因工程方面,馬鈴薯的RCA基因啟動子可用于表達只在植物綠色組織表達的防御基因來特異性的防御白粉病或者蚜蟲[11]。因此,研究植物RCA基因的啟動子,不僅有助于了解該基因的表達模式及調控機制,而且對于農業生產也具有重要意義。
啟動子是影響基因表達效率的重要因子,其順式作用元件決定了基因的表達量及表達模式[13]。啟動子區的順式作用元件可以響應光、植物激素、逆境脅迫和生物鐘等,從而調控下游基因的表達。對啟動子區進行順式作用元件分析,有助于了解下游基因的表達模式及調控機制。棉花作為重要的經濟作物,在農業生產中具有重要地位。目前,關于棉花RCA基因的研究主要集中在結構、活性和熱脅迫條件下的蛋白表達等方面[14-15],關于其轉錄調控的分子機制研究很少。為此,本研究利用棉花基因組測序結果,以陸地棉品種百棉1號為材料,通過PCR方法從基因組中擴增GhRCAα啟動子序列,并利用PlantCARE對其順式作用元件進行分析。同時為了驗證該啟動子的功能,利用熒光定量PCR技術研究了該基因的表達特性,并通過瞬時表達的方法進一步研究其活性。旨在摸清RCA表達調控的分子機制,從而為提高棉花光合速率和產量提供理論基礎。
1 材料和方法
1.1 供試材料
用于GhRCAα啟動子克隆或組織表達模式分析的材料為陸地棉品種百棉1號,該材料由河南科技學院棉花研究所提供,于2015年4月20日種植于河南科技學院喬謝試驗田。在幼苗期,當第3復葉完全展開時,取完全展開葉并立即速凍于液氮中,用于基因組DNA的提取。在盛花期取棉花植株的根、莖、葉和花(花瓣和萼片)及初鈴期的鈴殼和纖維用于棉花GhRCAα的組織表達模式分析。
1.2 DNA的提取及GhRCAα啟動子的擴增
使用基因組DNA提取試劑盒(TIANGEN, China, DP321)提取百棉1號葉片DNA。以基因組DNA為模板進行GhRCAα啟動子序列的擴增。引物序列為:GhRCAα-Promoter-F:5′- TCAGGGTATCTTCAGGGGTA-3;GhRCAα-Promoter-R:5′-GTCCTTTTCGGTCTGTGTCT-3′。
GhRCAα啟動子擴增采用50 μL反應體系,包括:10 μL 5×PrimeSTAR GXL Buffer (Mg2+plus),4.0 μL 2.5 mmol/LdNTPs,1.5 μL 10 mmol/L正反向引物,1 μL 1.2 U/μL PrimeSTAR GXL DNA Polymerase (TaKaRa, Japan),4.0 μL DNA模板及28.0 μL ddH2O。PCR反應程序:94 ℃ 2 min;98 ℃ 20 s, 61 ℃ 2.5 min, 共35個循環;最后4 ℃保溫10 min。PCR擴增產物經1%的瓊脂糖凝膠電泳分離檢測,根據比對Marker條帶,鑒定為目的基因的PCR產物經膠回收試劑盒(Axygen, China, AP-GX-4)純化后送深圳華大公司測序。
1.3 序列分析
利用植物順式作用元件數據庫PlantCARE(http://bioinformatics.psb.ugent.be/webtools /plantcare/html/)[16]在線預測分析所克隆序列的順式作用元件。
1.4 RNA 的提取和cDNA的合成
使用RNA提取試劑盒(BioFLUX, BSC65S1B),對所取樣品進行總RNA提取。再用1%瓊脂糖凝膠電泳確定RNA質量,而后RNA儲存于- 80 ℃備用。使用TaKaRa反轉錄試劑盒對約2 μg RNA進行反轉錄,生成cDNA。反轉錄生成的cDNA采用棉花內參基因Actin檢測質量,并保存于-20 ℃。
1.5 基因表達量測定
以反轉錄得到的各組織cDNA為模板,以棉花Actin基因為內參,進行熒光定量PCR分析。棉花GhRCAα和Actin基因的特異性熒光定量PCR引物分別為:QGhRCAα-F:5′-AGCCGCCGACATAATCAAAAAG-3′; QGhRCAα-R:5′-ACGGATGAGAGGAGCATACA GT-3′; QActin-F:5′-ATCCTCCGTCTTGACCTTG-3′; QActin-R:5′-TGTCCGTCAGGCAACTCAT-3′。
熒光定量PCR擴增體系(20 μL):2 × SYBR Premix ExTaq(TaKaRa) 10 μL,正反引物各1 μL,模板cDNA 2 μL,ddH2O 6 μL。每個樣品設置3個重復。擴增反應在Bio-rad Chromo4熒光定量PCR儀上運行,擴增條件為95 ℃ 2 min;95 ℃ 30 s,56 ℃ 30 s,72 ℃ 30 s,40 個循環。采用2-ΔΔCt法分析數據,確定基因的相對表達量。
1.6 載體的構建及農桿菌介導的轉化
已克隆的2 000 bp啟動子片段通過載體相應的酶切位點(EcoRⅠ和PstⅠ)連入含有GUS報告基因的雙元載體pCAMBIA1381Z(Cambia, Australia)中。構建好的雙元載體(GhRCAα∷GUS)以及載體pCAMBIA1301(陽性對照:CaMV35S∷GUS)和pCAMBIA1381Z(陰性對照:無啟動子)分別轉化至根癌農桿菌菌株EHA105中,并通過農桿菌介導煙草轉化的方法進行遺傳轉化。
1.7 組織化學染色
GUS活性檢測參照Jefferson等[17]的方法并略加修改。取待檢測的植物組織,加入GUS染色液,37 ℃溫育過夜24 h。接著,用體積分數為70%,80%,90%,100%的乙醇系列進行脫色。最后,在體式顯微鏡下觀察組織的染色情況并拍照。
2 結果與分析
2.1 GhRCAα啟動子的克隆
以已克隆出的陸地棉GhRCAαcDNA序列(GenBank 登錄號:AF329935)為查詢序列BlastN陸地棉全基因組[18],得到起始密碼子ATG上游2 000 bp調控序列,然后根據該序列設計引物,在百棉1號DNA(圖1-A)中擴增GhRCAα啟動子。GhRCAα的理論大小為2 309 bp,所得片段電泳結果顯示,與預期的片段大小相符(圖1-B)。經Blast比對確認,該片段為GhRCAα啟動子序列。
2.2 GhRCAα啟動子的序列分析
將分離獲得的棉花GhRCAαATG上游2 000 bp的啟動子序列提交到啟動子預測網站plantCARE在線分析啟動子區的順式作用元件組成。結果表明,許多重要的順式作用元件,包括TATA-box、 HSE、 G-box、5UTR Py-rich和ciradian等特異地存在于GhRCAα啟動子區(圖2),且不同順式作用元件的拷貝數也存在很大差異(表1)。進一步分析表明,這些順式作用元件主要分為以下6類(表1)。第1類是光響應元件,包括G-box、I-box和GT1-motif等可能參與GhRCAα光調控的順式作用元件;第2類是生物鐘調控元件ciradian,該元件可能參與GhRCAα的晝夜表達模式調控;第3類是與植物激素響應有關的GARE-motif和ABRE元件,表明GhRCAa的表達可能受植物激素的調節;第4類是與逆境響應有關的元件,包括響應干旱脅迫的MBS元件,熱激響應元件HSE等;第5類是基礎表達相關的元件,包括CAAT-box (與增強轉錄效率有關)和TATA-box等,說明GhRCAα啟動子是一個典型的由RNA 聚合酶Ⅱ結合的啟動子;第6類是包括CGTCA-motif、TGACG-motif、as1、Skn-1_motif以及CAT-box等其他類順式作用元件,其中as1具有參與根特異性表達的功能,CGTCA-motif和TGACG-motif具有參與茉莉酸響應的功能。以上這些結果表明GhRCAα的表達可能受光、生物鐘、逆境及植物激素等的調控。
2.3 GhRCAα表達模式的分析
由于GhRCAα啟動子區包含許多響應光、特定組織或生物鐘的順式作用元件,為了摸清這些元件的存在對GhRCAα表達有何影響,采用熒光定量PCR方法來研究GhRCAα在棉花不同組織中的表達情況。結果表明:GhRCAα在葉片中大量表達,而葉片是光合作用進行的主要位置,在纖維、莖、萼片、根、花瓣和鈴殼中表達量很低(圖3)。先前對棉花GhRCAα進行晝夜表達模式分析發現,GhRCAα基因的表達具有晝夜節律性,在光周期開始后1 h表達量最高,午夜表達量最低[15]。這些結果表明,GhRCAα的表達具有組織特異性且受晝夜模式的影響。該結果與啟動子區的順式作用元件分析結果相一致。

圖1 棉花基因組DNA的提取(A)及GhRCAα啟動子序列的擴增(B)Fig. 1 The extraction of genomic DNA (A) and the PCR amplification (B) of GhRCAα promoter in cotton

ATG上游的2 000 bp片段定義為啟動子區域,部分預測的順式作用元件在圖中用灰色陰影表示;
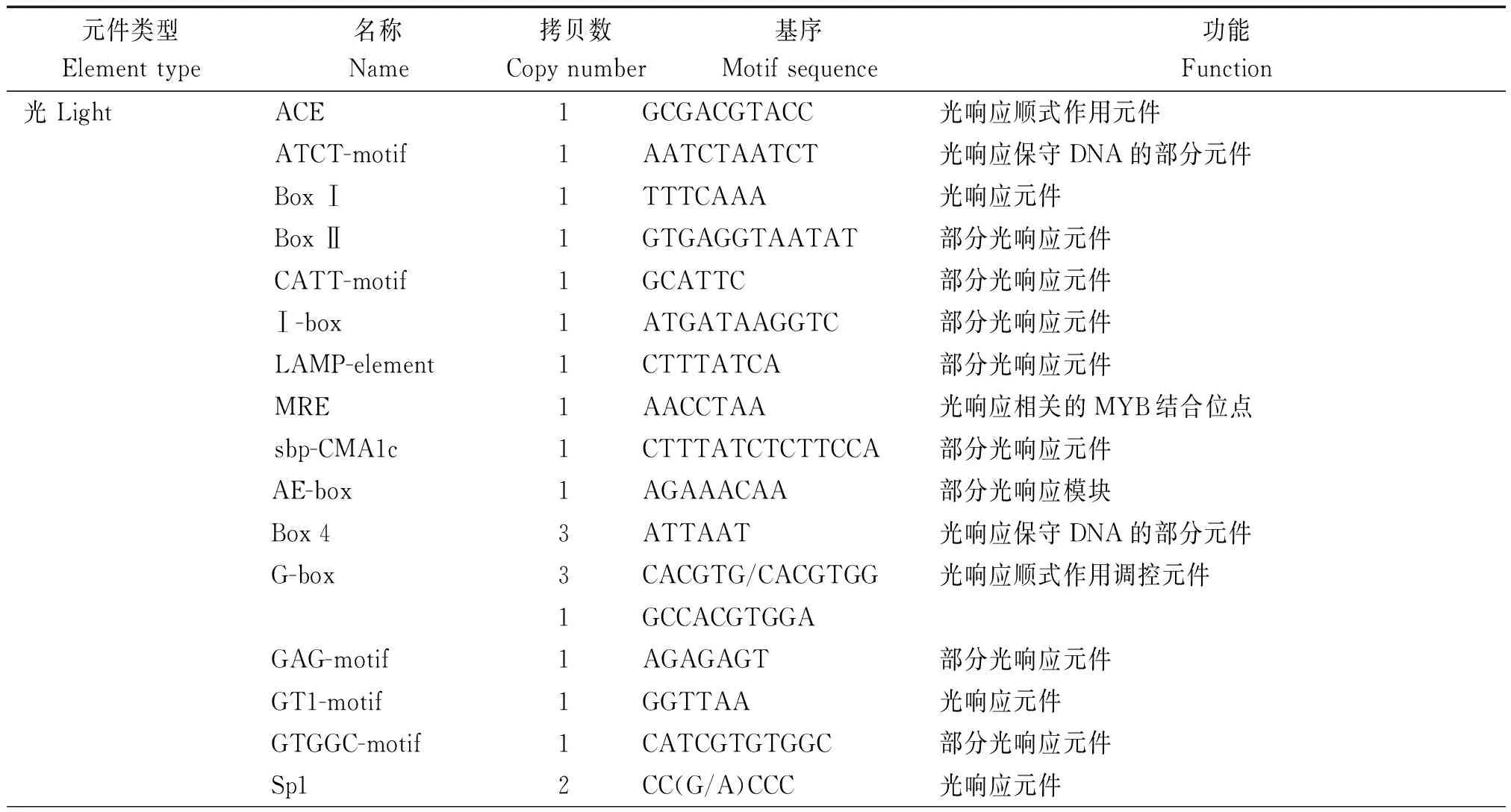
表1 GhRCAα啟動子區包含的順式作用元件Tab.1 Cis-acting elements in promoter of GhRCAα
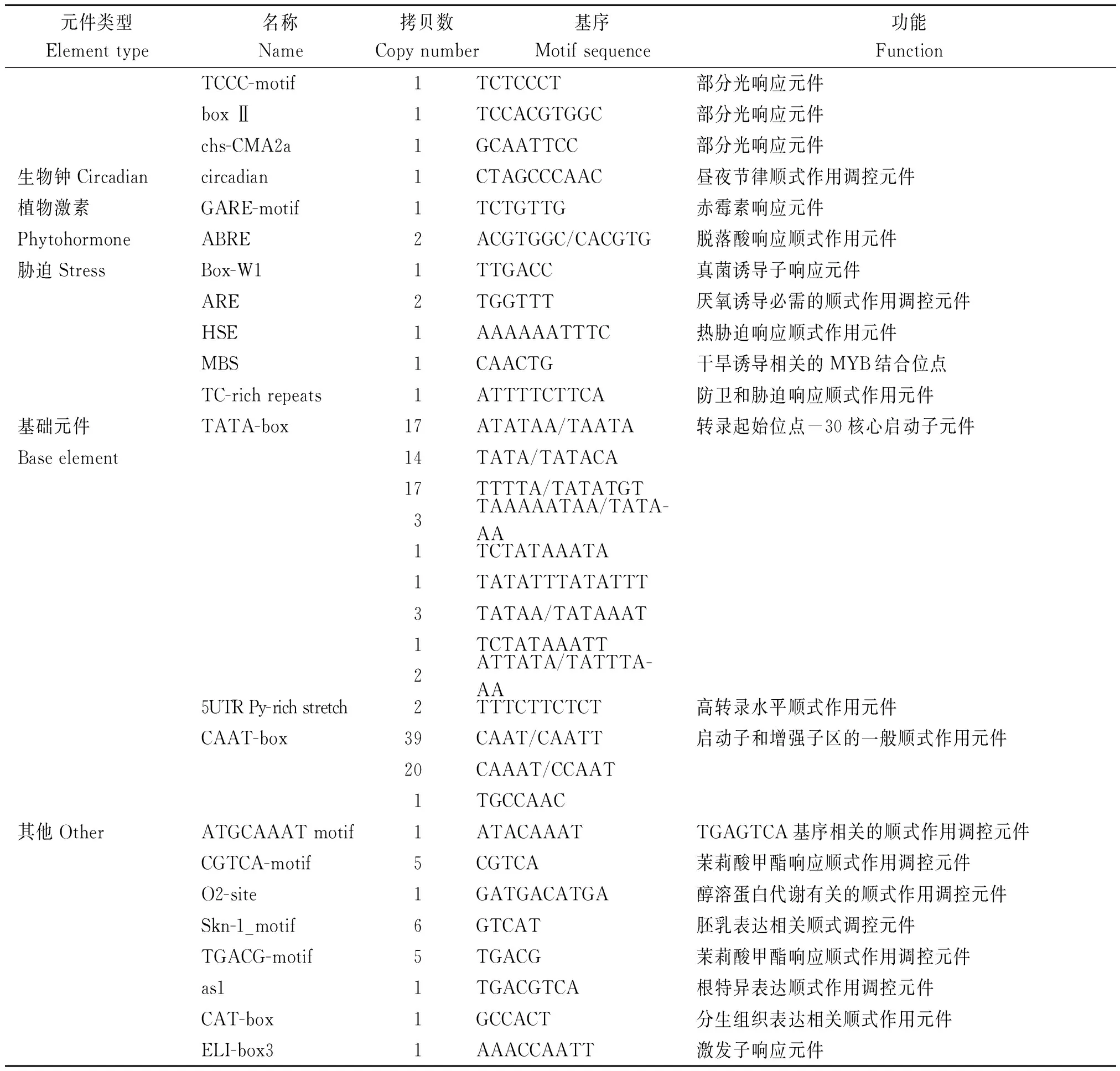
表1(續)
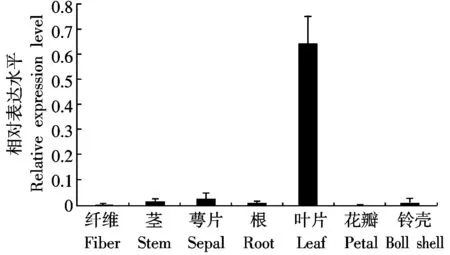
圖3 棉花 GhRCAα的組織特異性表達分析Fig. 3 Analysis of organ-specific expression of GhRCAα in cotton
2.4 植物表達載體的構建及活性分析
將克隆的2 000 bp的片段,用雙酶切法連接到植物表達載體pCAMBIA1381Z。對重組質粒進行雙酶切鑒定,得到1條約為2 000 bp的特異性條帶(圖4),與預期片段一致,表明已成功構建了由該啟動子驅動報告基因GUS的植物表達載體(GhRCAα∷GUS)。
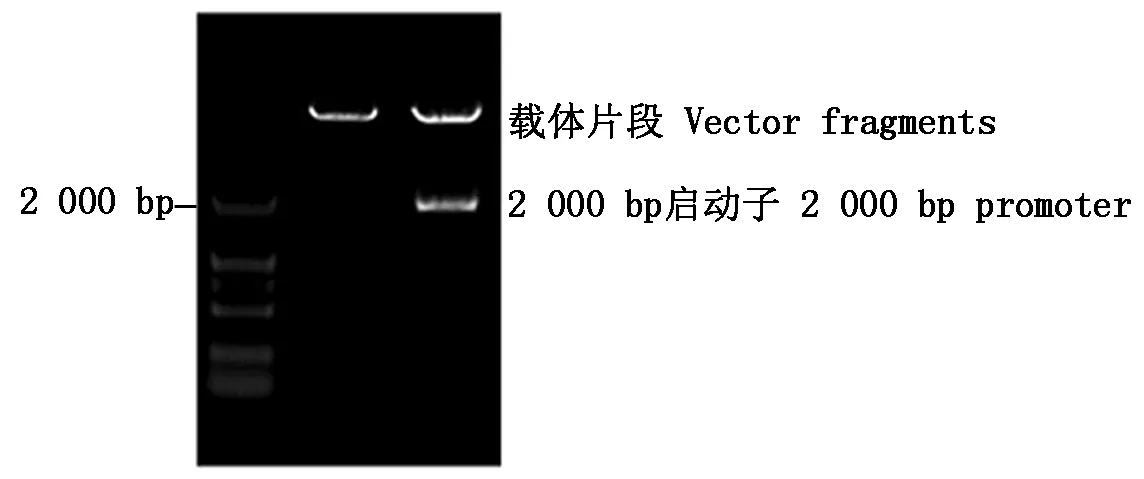
圖4 表達載體GhRCAα∷GUS 的酶切分析Fig.4 Restriction analysis of GhRCAα∷GUS vector
將重組表達載體(GhRCAα∷GUS)、pCAMBIA1301(陽性對照,CaMV35S∷GUS)以及pCAMBIA1381Z(陰性對照),通過農桿菌介導的煙草遺傳轉化進行瞬時表達。結果表明,陽性對照CaMV35S 啟動子(圖5-A)以及重組載體GhRCAα∷GUS(圖5-B)轉化的煙草葉片均能檢測到GUS的表達產物,但是與陽性對照(CaMV35S 啟動子)相比,目的載體染色弱于陽性對照,表明其活性比CaMV35S啟動子低;pCAMBIA1381Z 載體沒有啟動子來驅動GUS的表達,所以檢測不到GUS表達(圖5-C)。這些結果表明,本研究克隆的GhRCAα啟動子能夠驅動下游基因GUS的表達,具有驅動目標基因表達的活性。
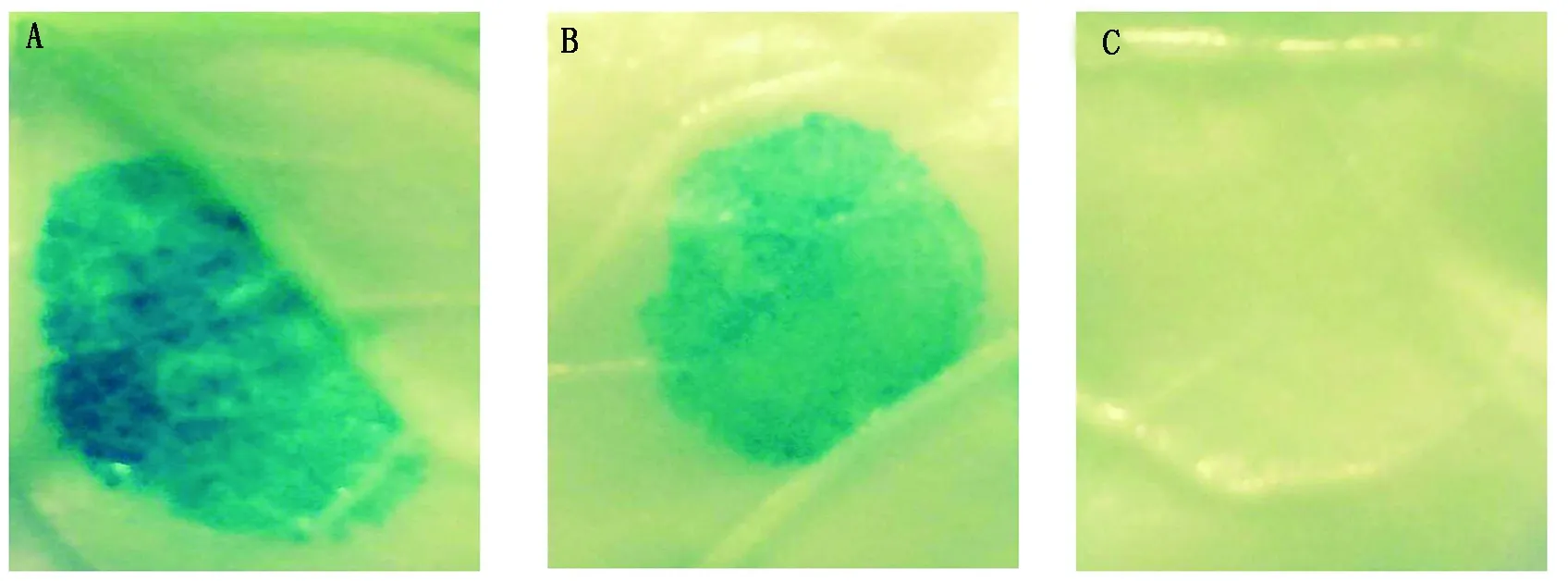
A.35S啟動子; B.GhRCAα啟動子;C.不含啟動子的對照。
3 結論與討論
啟動子是決定外源基因在受體植物中轉錄效率高低的關鍵因子,合適的啟動子將會促進外源基因的高效表達[19]。組成型啟動子能使外源基因在受體植物中高效、持續的表達,但在特定時間或特定組織部位的表達達不到預期效果[20]。因此,誘導型或特異型啟動子的研究越來越受到廣大研究者們的青睞,發掘一種組織特異型或誘導型啟動子對于轉基因作物育種具有重要意義[21-23]。RCA基因作為光合作用過程中的一個關鍵基因,其表達除了受光誘導和晝夜模式的影響以外,還具有組織特異性,且受發育時期和植物激素的影響[5-8]。與其表達模式相一致,RCA啟動子在高等植物的綠色組織及光誘導下具有活性,可作為一種組織特異型和誘導型啟動子[9-11]。因此,克隆植物RCA基因的啟動子,明確其組織特異性表達,可為轉基因育種提供一種組織特異型或誘導型啟動子。
近年來,關于棉花RCA基因啟動子的克隆及功能研究很少,而關于RCA的結構、活性和蛋白表達已開展很多研究[14-15,24]。本研究對棉花GhRCAα啟動子進行克隆及順式作用元件分析表明,GhRCAα啟動子除了具有基本的調控元件,還包含許多與光、生物鐘、植物激素以及逆境響應有關的調控元件,這些元件特異地存在于GhRCAα啟動區,并在拷貝數上存在很大差異。已有報道表明,啟動子誘導表達的強度主要依賴于順式作用元件的拷貝數,更具體的來說就是順式作用元件相對于TATA-box的間距[25-26]。對大豆的RCA研究表明,重要啟動子元件如G-box和pyrimidine-rich 5′-UTR在拷貝數目上的差異可能是導致GmRCAα和GmRCAβ表達量差異的一個原因[3]。棉花中GhRCAα啟動子區這些重要順式調控元件拷貝數變異可能參與了調控下游基因GhRCAα表達的強度及模式,但是關于其影響GhRCAα表達的分子機制還不清楚,在今后的研究中仍需進一步實驗驗證。
利用熒光定量PCR對GhRCAα表達特性進行分析表明,GhRCAα在棉花的不同組織表達量存在很大差異,在光合作用進行的主要位置葉片中表達量最高,其他組織表達量很低,其表達具有組織特異性。與此相一致的是大量光響應元件(G-box、ACE及I-box等),組織特異表達元件(as1等)特異地存在于GhRCAα啟動子區。已有研究表明,植物的組織特異性表達可能是來源于根特異性表達元件的負調控,也可能是來源于葉特異性表達元件的正調控[10,27-28]。GhRCAα啟動子區根特異性表達元件as1的存在可能是作為一個負調控元件參與組織特異性表達,但仍需實驗驗證。在植物的生長發育過程中,光不但作為一種重要的信號來調控相關基因的表達,而且還直接參與光合作用[29]。GhRCAα啟動子區光響應元件的大量存在可能參與GhRCAα的光調控及光合組織葉片中GhRCAα的大量表達。另外,晝夜節律響應元件circadian及同時參與生物鐘及光響應的調控元件I-box[30-32]均存在于GhRCAα的啟動子區,說明其表達可能受到光周期的調節,這與先前報道的GhRCAα的表達具有晝夜節律性的結果相一致[15]。對棉花[15]、水稻[33]和菠菜[34]的RCA研究表明,45 kDa(RCAα)亞型可能在光合機構適應體外中度熱脅迫方面起著重要作用,熱激響應元件HSE特異存在于GhRCAα啟動子區可能賦予該基因在熱脅迫條件維持下Rubisco活性方面的重要作用。總的來說,與其他物種一樣[5-8],棉花的GhRCAα表達具有組織特異性,且可能受熱脅迫和晝夜模式的影響,該基因啟動子區相關調控元件的存在可能決定了該基因的表達特性。然而,GhRCAα啟動子區哪些核心元件參與調控GhRCAα的表達仍不清楚。大量逆境響應元件及植物激素響應元件存在GhRCAα啟動子區是否意味著該基因在植物應對逆境脅迫方面也具有重要作用,在今后的研究中,通過啟動子區的缺失試驗分析、定點突變及逆境條件下基因表達特性研究,將有助于了解GhRCAα啟動子調控目標基因表達的分子機制。
本研究克隆獲得的GhRCAα啟動子可以驅動GUS基因的表達,表明該啟動子片段具有驅動目標基因表達的活性,因此,有望用于植物的遺傳轉化,進而更好地調控重要基因的特異性表達。本研究結果不僅有助于進一步深入認識棉花GhRCAα基因功能及其表達調控規律,也可為植物遺傳轉化提供一種組織特異型或誘導型的啟動子。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
