灰葡萄孢BcKMO基因的原核表達分析
2017-12-12 09:03:20劉媛媛姜婷婷時翠平邢繼紅董金皋
華北農學報 2017年1期
王 敏,劉媛媛,周 帆,姜婷婷,鄭 旭,張 靖,時翠平,邢繼紅,董金皋
(河北農業大學,真菌毒素與植物分子病理學實驗室,河北 保定 071001)
灰葡萄孢(Botrytiscinerea)是一種死體營養型植物病原真菌,其寄主范圍廣泛,可引起蔬菜、水果等200多種植物的灰霉病,給農業生產造成巨大經濟損失[1-3]。目前,灰霉病預防以化學藥劑為主,但其連續使用,易使病菌產生抗藥性,且危害人畜安全,對環境造成壓力,嚴重破壞生態平衡[4-5]。因此,尋找一種快速抑制病原菌、特異性強、對環境友好的新型生物殺菌劑迫在眉睫[6]。犬尿氨酸單加氧酶(Kynurenine 3-monooxygenase,KMO)是NADPH依賴的黃素蛋白羥化酶,位于犬尿氨酸途徑的中心。KMO催化犬尿氨酸在它的酚環第3位羥基化,生成3-羥基犬尿氨酸(3-HK)[7-9]。犬尿氨酸途徑主要存在于色氨酸分解代謝,廣泛存在于真核生物和原核生物中。犬尿氨酸代謝產生一些生物活性中間體,帶有不同生理行為,參與各種神經退行性疾病的發病機理,包括阿爾茨海默病、帕金森癥和亨廷頓疾病[10-11]。近年來,在一些微生物中也發現了類似的犬尿氨酸途徑,如熒光假單胞菌(Pseudomonasfluorescens)和哈氏噬纖維菌(Cytophagahutchinsonii)[12-14]、綠膿單胞菌(Pseudomonasaeruginosa)[15]、金羊毛鏈霉菌(Streptomyceschrysomallus)[16],犬尿氨酸途徑在安曲霉素生物合成中起重要作用[17]。由此,筆者推測灰葡萄孢中也存在犬尿氨酸途徑[18]。河北農業大學真菌毒素與植物分子病理學實驗室前期研究中,克隆了灰葡萄孢BcKMO基因,明確了BcKMO正調控灰葡萄孢的生長、發育,負調控灰葡萄孢的致病力[19];確定了BcKMO通過調控灰葡萄孢的胞壁降解酶活性、毒素活性、產酸能力及致病相關基因的表達而影響病菌的致病力[20]。但是BcKMO基因編碼產物的功能及其與病菌致病力調控之間的關系尚未明確。因此,本試驗對灰葡萄孢BcKMO基因進行原核表達分析,獲得純化的BcKMO蛋白,為下一步的酶活力測定及其互作蛋白的篩選奠定基礎,并為闡明單加氧酶及其所在的犬尿氨酸途徑與病菌致病力之間的關系提供理論依據。
1 材料和方法
1.1 試驗材料
灰葡萄孢野生型菌株BC22由河北農業大學真菌毒素與植物分子病理學實驗室保存;大腸桿菌DH5α和BL21、限制性內切酶、T4 DNA連接酶購自TaKaRa公司;pMD19-T載體、pGEX4T-1載體購自Pharmacia Biotech公司;RNA提取試劑盒、反轉錄試劑盒購自北京全式金公司。
1.2 試驗方法
1.2.1 引物設計 根據BcKMO基因序列,采用Primer 5.0軟件設計兩端帶有NotⅠ和SmaⅠ酶切位點的特異性引物yhKMO-F(5′-ATAAGAATGCGG CCGCATGCCGTCCTTGTTGATC-3′)和yhKMO-R(5′-TCCCCCGGGAATGTCTGCACCATCCTCTAAAG-3′)。其中下劃線的部分分別是NotⅠ和SmaⅠ的酶切位點。
1.2.2BcKMO基因的克隆 以灰葡萄孢野生型菌株BC22為試材,提取RNA,反轉錄成cDNA,用高保真Taq酶擴增BcKMO基因。PCR反應體系:模板cDNA 2 μL、yhKMO-F(10 μmol/L) 0.5 μL、yhKMO-R(10 μmol/L) 0.5 μL、LATaq酶0.5 μL、10×LA PCR Buffer Ⅱ 2.5 μL、10 mmol/L dNTP 2 μL、ddH2O 18 μL,共25 μL。經PCR擴增,瓊脂糖凝膠電泳檢測,目的條帶回收后與載體pMD19連接,連接后的產物轉化大腸桿菌DH5α感受態細胞,陽性克隆測序正確后進行質粒pMD19-T-BcKMO的提取。
1.2.3 原核表達載體的構建與鑒定 用NotⅠ和SmaⅠ分別對pMD19-T-BcKMO和pGEX4T-1質粒進行雙酶切,回收目的片段后連接,轉化大腸桿菌DH5α感受態細胞,通過菌落PCR、酶切驗證及測序驗證后,確定原核表達載體pGEX4T-1-BcKMO-GST構建成功。將pGEX4T-1-BcKMO-GST和pGEX4T-1分別轉化大腸桿菌BL21,確定陽性克隆進行蛋白的誘導表達。
1.2.4 不同濃度的IPTG誘導對BcKMO蛋白表達的影響 將轉化了pGEX4T-1-BcKMO-GST和pGEX4T-1的大腸桿菌BL21分別在LB培養基中培養8 h,37 ℃轉接,培養至OD600約為0.7,分別加入不同濃度的IPTG至終濃度為0.1,0.2,0.4,0.8,1.0 mmol/L,28 ℃誘導14 h,收集菌體,棄上清,用pH值8.8 Tris-HCl和5×SDS-PAGE Loading Buffer重懸沉淀,沸水浴10 min,離心取上清進行SDS-PAGE,檢測目的蛋白的表達情況,進而確定最適IPTG誘導濃度。
1.2.5 不同誘導時間對BcKMO蛋白表達的影響 利用上述得到的最適IPTG誘導濃度,28 ℃誘導pGEX4T-1-BcKMO-GST和pGEX4T-1的表達,誘導時間分別為4,8,12,16,20 h,收集菌體,提取蛋白,進行SDS-PAGE,確定誘導表達的最佳時間。
1.2.6 BcKMO蛋白的純化 利用上述得到的最適IPTG誘導濃度和最佳誘導時間,在28 ℃進行誘導表達,收集菌體,棄上清,用pH值8.8 Tris-HCl重懸菌體沉淀,加入蛋白酶抑制劑,利用超聲細胞破碎儀破碎菌體,超聲功率100 W,工作10 s,間歇20 s,循環30次,離心收集上清,4 ℃保存備用。
使用ProteinIsoTMGST Resin對收集的上清進行目的蛋白的純化,取1 mL GST beads加入20 mL Binding Buffer (50 mmol/L Tris-HCl pH值8.0,150 mmol/L NaCl)進行平衡,4 ℃混勻10 min,700 r/min離心5 min,棄上清,加入20 mL Binding Buffer重懸,重復洗3次,棄上清,冰上備用;將菌體上清加入上述平衡的GST beads,4 ℃混勻2 h,700 r/min離心5 min,棄上清,留沉淀;向沉淀中加入20 mL Binding Buffer(含0.1% TritonX-100),4 ℃混勻10 min,700 r/min離心5 min,棄上清,重復洗滌2次,棄上清,留沉淀;向沉淀中加入500 μL洗脫緩沖液(50 mmol/L Tris-HCl pH值8.0,10 mmol/L 谷胱甘肽),4 ℃混勻1 h,700 r/min離心5 min,收集洗脫液,重復洗2次,分別得到洗脫液,進行SDS-PAGE,檢測目的蛋白的純化情況。
1.2.7 Western Blot分析 利用Western Blot技術,對目的蛋白進行檢測。分別將純化后的蛋白和菌體的全蛋白進行SDS-PAGE,利用轉膜儀轉到PVDF膜上,用5%脫脂奶粉封閉2 h,用PBST洗滌4次,按1∶1 000加入GST Tag Antibody,雜交1 h,用PBST洗滌4次,每次5 min,按1∶10 000加入goat anti-mouse HRP antibody,雜交1 h,用PBST洗滌5次,每次5 min,然后在PVDF膜上加入Western Blot發光檢測液A、B混合液1 mL,用保鮮膜封好,放在暗匣內,在暗室進行顯影[21]。
2 結果與分析
2.1 BcKMO基因原核表達載體的構建
提取灰葡萄孢野生型菌株BC22總RNA,反轉錄成cDNA,用高保真Taq酶擴增BcKMO基因(圖1-A)。回收PCR擴增產物,與pMD19-T載體連接,轉化大腸桿菌,得到pMD19-T-BcKMO陽性克隆。用NotⅠ和SmaⅠ進行酶切檢測,獲得了2條與目的片段大小一致的條帶(圖1-B),表明BcKMO基因已連接pMD19-T載體。將pGEX4T-1載體和測序正確的pMD19-T-BcKMO載體分別進行NotⅠ和SmaⅠ雙酶切,回收目的片段,用T4連接酶連接,轉化大腸桿菌,通過酶切驗證獲得了與目的條帶大小一致的條帶(圖1-C),表明原核表達載體pGEX4T-1-BcKMO-GST構建成功。
2.2 不同濃度的IPTG誘導對BcKMO蛋白表達的影響
為確定IPTG誘導使用的最適濃度,37 ℃轉接培養至OD600約為0.7,分別加入不同濃度的IPTG至終濃度為0.1,0.2,0.4,0.8,1.0 mmol/L,28 ℃誘導14 h,收集菌體,以未經誘導的菌體為對照,進行SDS-PAGE檢測。結果發現,轉化pGEX4T-1載體的菌株經誘導表達出約26 kDa的蛋白條帶,與GST蛋白大小一致(圖2-A),而帶有pGEX4T-1-BcKMO-GST載體的菌株誘導表達出約71 kDa的蛋白條帶(圖2-B),與融合蛋白大小一致(GST蛋白為26 kDa,BcKMO蛋白45 kDa)。當IPTG濃度為0.1 mmol/L時,融合蛋白的表達水平較低;當IPTG濃度為0.2~1.0 mmol/L時,融合蛋白的表達水平沒有明顯變化(圖2-B),由此確定IPTG誘導使用的最適濃度為0.2 mmol/L。
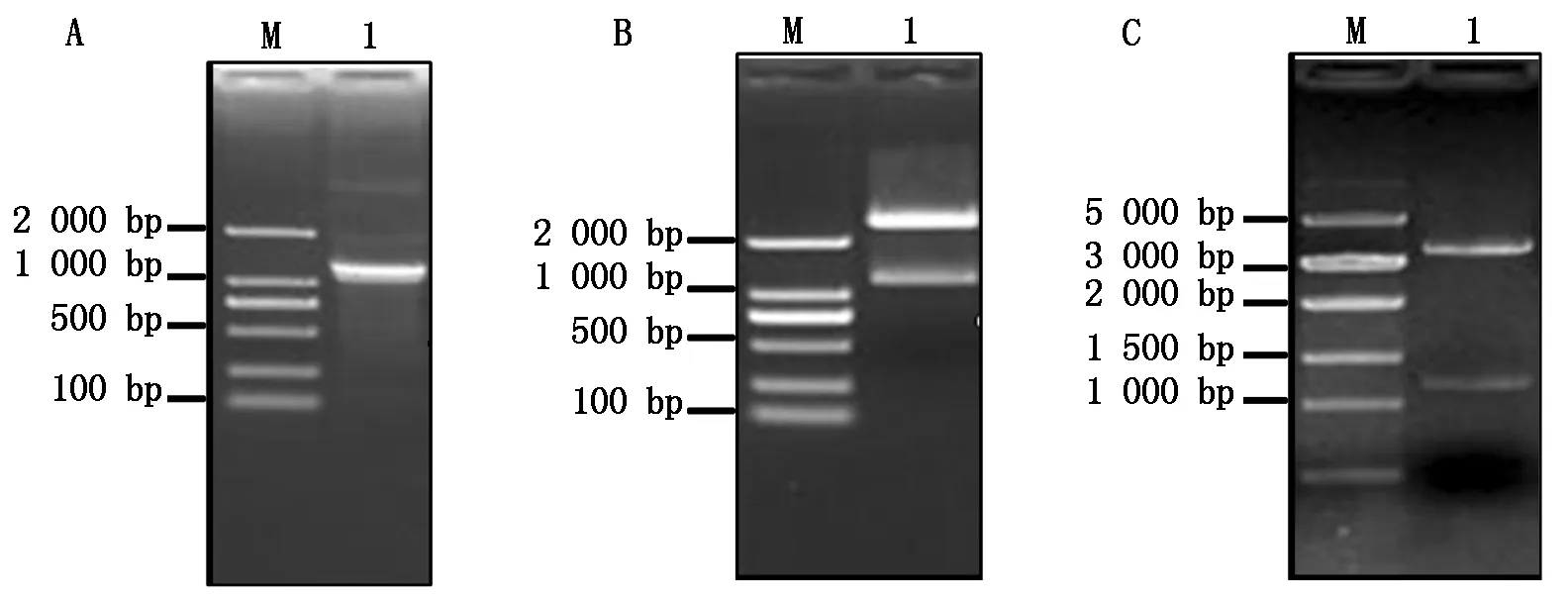
A.BcKMO基因的擴增;B.pMD19-T-BcKMO載體的酶切鑒定;C.pGEX4T-1-BcKMO-GST載體的酶切鑒定。A.PCR amplification of BcKMO;B.Identification of pMD19-T-BcKMO by restriction enzyme digestion;C.Identification of pGEX4T-1-BcKMO-GST by restriction enzyme digestion.
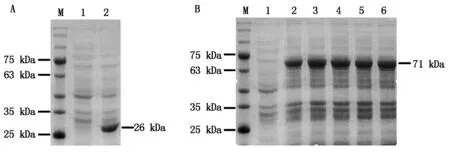
M.蛋白分子量標準;A.誘導表達pGEX4T-1載體;1.IPTG未誘導;2.IPTG誘導GST蛋白;B.pGEX4T-1-BcKMO-GST的誘導表達;1.IPTG未誘導;2~6.IPTG誘導濃度分別為0.1,0.2,0.4,0.8,1.0 mmol/L。M.Protein Marker;A.Expression of pGEX4T-1 with IPTG induction;1.Without IPTG;2.IPTG induces GST proteins;B.Expression of pGEX4T-1-BcKMO-GST with IPTG induction;1.Without IPTG;2-6.Expression of pGEX4T-1-BcKMO-GST with 0.1,0.2,0.4,0.8,and 1.0 mmol/L IPTG.
2.3 不同誘導時間對BcKMO蛋白表達的影響
使用0.2 mmol/L IPTG誘導pGEX4T-1-BcKMO-GST表達,誘導時間分別為4,8,12,16,20 h,收集菌體,以未誘導的菌體作為對照,進行SDS-PAGE檢測。結果發現,在不同誘導時間下均可以誘導表達71 kDa的蛋白;當誘導時間為0~12 h時,隨著誘導時間的延長,目的蛋白的表達水平逐漸增加;當誘導時間為12~20 h時,目的蛋白的表達水平沒有明顯變化(圖3)。由此確定,蛋白誘導表達的最佳時間為12 h。
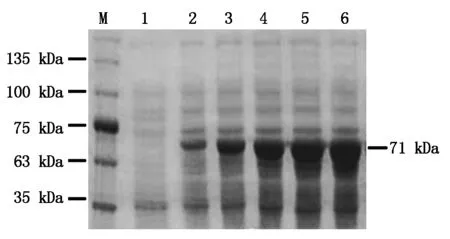
M.蛋白分子量標準;1.IPTG未誘導;2~6.IPTG誘導時間分別為4,8,12,16,20 h。M.Protein Marker;1.Without IPTG;2-6.Expression of BcKMO-GST with IPTG induction for 4,8,12,16,and 20 h.
2.4 BcKMO-GST蛋白的純化
利用上述得到的最佳誘導條件,進行BcKMO-GST蛋白的誘導表達,并使用ProteinIsoTMGST Resin對收集的上清進行蛋白的純化。純化后的蛋白進行SDS-PAGE分析,獲得了單一的蛋白條帶,大小與目的蛋白大小一致(圖4),表明BcKMO-GST蛋白純化成功。
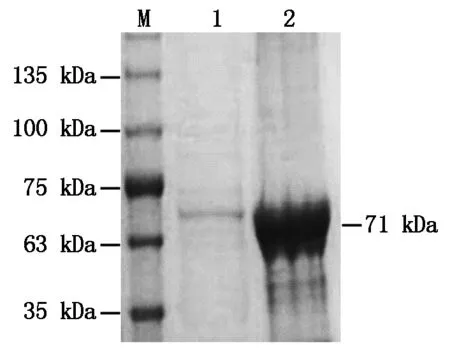
M.蛋白分子量標準;1.純化的BcKMO-GST蛋白;2.純化之前的上清液。M.Protein Marker;1.Purified protein BcKMO-GST;2.Supernatant before purification.
2.5 BcKMO-GST蛋白的Western Blot分析
以GST Tag antibody為一抗,goat anti-mouse HRP antibody為二抗,對純化獲得的蛋白進行Western Blot分析。結果發現,純化的蛋白獲得了單一的陽性條帶,大小與目的蛋白大小(71 kDa)一致(圖5),表明BcKMO蛋白體外誘導表達成功。
3 討論
外源基因在大腸桿菌體內能否高效表達受到很多因素的影響和限制,如外源基因自身的理化性質、大腸桿菌菌株的類別、所用表達載體種類、誘導物濃度、誘導溫度及誘導時間等[22]。一般誘導物選用IPTG[23],低溫有利于增強蛋白的穩定性和正確折疊,容易獲得可溶性的蛋白,而較高溫度則易形成包涵體[24],不利于目的蛋白的純化,因此,本試驗采用28 ℃長時間誘導,降低合成速度,使該蛋白具有充分的時間進行折疊,使二硫鍵正確配對,使蛋白達到足夠的溶解度。本試驗通過設置不同IPTG濃度和不同誘導時間進行試驗[25],確定了pGEX4T-1-BcKMO-GST融合蛋白誘導表達的最適濃度和最佳時間。超聲破碎后使用GST親和的beads進行目的蛋白的純化,經SDS-PAGE檢測和Western Blot分析確定BcKMO蛋白體外誘導表達成功。但是,蛋白純化的條件仍需進一步優化,進而獲得更高含量的目的蛋白,為后續進行該單加氧酶的酶活力測定及其互作蛋白的篩選奠定基礎。
文獻報道,犬尿氨酸單加氧酶的穩態動力學參數通過酶活性的初始速率測量獲得。KMO催化NADPH依賴的L-犬尿氨酸羥基化成為3-羥基犬尿氨酸,酶促反應能通過接下來的NADPH在340 nm吸光度的降低來監測。速率分析實施采用1 cm路徑長度石英微量吸收測定人KMO是37 ℃,釀酒酵母KMO是30 ℃[11]。GST pull-down試驗可用于捕獲與靶蛋白相互作用的目的蛋白,從而證實2種蛋白之間的相互作用或篩選相應的蛋白。試驗后期擬利用Pull-down技術篩選BcKMO蛋白的互作蛋白,為闡明BcKMO及其所在的犬尿氨酸途徑與病菌致病力之間的關系提供理論依據。
