基因組選擇及其在作物育種中的應用
2018-01-16 18:38:40姜淑琴孫炳蕊潘大建范芝蘭陳文豐
廣東農業科學 2017年9期
陳 雨,姜淑琴,孫炳蕊,潘大建,范芝蘭,陳文豐,李 晨
(1.廣東省農科院水稻研究所/廣東省水稻育種新技術重點實驗室,廣東 廣州 510640;2.中國農業大學農學院,北京 100193)
品種(系)選育是遺傳改良的重要手段之一,在人類農業生產中占著重要地位。傳統育種主要根據個體表型和個人經驗,結合簡單的統計方法,對個體進行選擇,存在周期長、效率低、部分性狀難以測量等缺點。隨著遺傳學及分子生物學技術的發展,分子標記輔助選擇(marker-assisted selection,MAS)應運而生;育種工作者可以直接通過檢測分子標記,利用標記與目標性狀基因緊密連鎖的特點,選擇優質基因(型)或者強性狀優勢個體[1]。MAS育種方法主要依賴于QTL定位的準確性,然而現有的QTL分析在尋找基因的廣度、深度和精度等方面尚有不足:一是基于雙親的作圖群體所含優良目標基因有限[2],二是雖然對少量效應大的遺傳變異具有較好的適應性,但大部分重要的性狀還受大量微效基因的協同作用,且容易受環境的影響[3]。近年來,由于高通量測序技術的迅猛發展,開發了大量豐富且廉價的SNP標記,全基因組關聯分析(Genome-wide association study,GWAS)成為一種新型的MAS方法[4-5]。與QTL分析相比,GWAS通過對全基因組進行掃描,鑒定出與目標性狀有關聯的標記,不需要構建專門的作圖群體,能同時檢測多個序列變異,實現QTL的精細定位[6]。但由于受群體結構、因避免假陽性而采用嚴格顯著性閾值等因素的影響,只有少數顯著位點被利用。針對數量性狀由微效多基因控制這一育種問題,基因組選擇方法利用覆蓋全基因組的分子標記,對個體進行了遺傳評估與選擇。相對于GWAS,GS方法充分反映了目標性狀的遺傳變異,不需要做顯著性檢驗,即使標記效應微小,其導致的遺傳變異也能被捕獲。2001年Meuwissen等利用模擬數據通過GS方法對育種值的估計準確性達到85%[7]。本文對GS的原理和方法進行歸納,并基于影響GS的各種因素探討了提高選擇準確性的各種途徑,展望了GS在作物育種的應用前景,旨在為GS在作物育種中的應用提供一定的參考。
1 基因組選擇的原理和基本策略
基因組選擇的概念最早由Haley和Visscher在第六屆世界遺傳學應用于畜牧生產國際會議上提出[8],并最早于2001年由Meuwissen等完善并應用于動物育種模擬數據分析中[7]。GS根據連鎖不平衡原理(linkage disequilibrium,LD),假設影響性狀的每個基因至少與一個標記緊密連鎖,從而用該標記間接反映基因的效應[9]。GS的實現主要分3個步驟:首先,對參與分析的所有個體,利用覆蓋其全基因組范圍的分子標記進行基因型分型、過濾,獲得基因型數據;其次,選擇合適的訓練群體(Training Population,TP),調查其表型,根據表型和基因型數據構建數學模型,估計每個分子標記的效應;再次,利用這些效應值對僅有基因型數據的育種群體(Breeding Population,BP)估計其對應的基因組育種值(Genomic Estimated Breeding Value,GEBV),并篩選出育種值較大的個體。基因組選擇考慮了全基因組范圍的SNP標記,有效地提高了選擇的準確性,尤其是對低遺傳力的數量性狀[10]。基因組選擇不需要測量所有個體的表型,可以同時對多個性狀進行選擇,顯著地提高了育種效率,降低成本。
2 育種值估計方法
基因組選擇的核心是育種值的估計,目前用于GS的方法主要分3類。
2.1 基于混合線性模型的BLUP(Best Linear Unbiased Prediction)方法
BLUP主要是通過系譜構建個體親緣關系矩陣A,然后基于表型和系譜A,利用混合線性模型(Mixed Linear Model,MLM)計算個體的估計育種值(Estimated Breeding Value,EBV)。目前基于BLUP的GS方法主要有兩種:一種是通過已測定的基因型計算個體間的相關關系矩陣G,然后用G代替系譜關系矩陣A,估算個體的育種值,即GBLUP(Genomic Best Linear Unbiased Prediction)方法[11];另一種是基于等位基因效應的RRBLUP(Ridge Regression Best Linear Unbiased Prediction),該法將標記效應作為隨機效應,假定標記效應服從標準正態分布,有共同的方差,然后利用混合模型求解,每個個體的育種值等于其所在基因組標記效應的總和[7]。全基因組選擇中因變量數p(標記數)往往大于反應變量數n(個體數),這種情況下常規的最小二乘估計法預測能力較低,很可能導致多重共線性和過度擬合[12],嶺回歸就是一種專用于共線性數據分析的改良最小二乘法,用于提高預測的準確性。當個體效應矩陣等于標記系數矩陣與標記效應乘積時,GBLUP與RRBLUP等同。
2.2 基于MCMC(Markov chain Monte Carlo)和Gibbs抽樣的貝葉斯(Bayes)方法
在全基因組SNP中,并不是所有標記方差都相同。另外,只有少數SNP與影響性狀的QTL連鎖,其效應值或大或小;多數SNP與QTL不連鎖,沒有效應[13]。基于此,研究者提出了Bayes方法[7]。BayesA假設每個SNP都有特定的方差,且方差服從逆卡方分布;BayesB對標記進行了選擇,認為大部分位點(π)沒有遺傳方差,而少數標記(1-π)有各自不同的效應方差,方差服從逆卡方分布[7];BayesC將π作為未知參數,假定其服從均勻分布U(0,1);BayesCπ在BayesC的基礎上,假定標記效應方差相同;BayesDπ在BayesC的基礎上,將逆卡方分布中的尺度參數作為未知參數[14];貝葉斯壓縮法(Bayesian least absolute shrinkage and selection operator,Bayesian LASSO)相對于BayesB來說,假定標記效應服從雙指數分布[15]。根據以上的描述發現,貝葉斯方法的改進主要是在經典BayesA和BayesB的基礎上,或優化參數,或變更標記效應先驗分布。貝葉斯方法在某些性狀上優于BLUP方法[7,16],但仍存在計算時間長、先驗分布超參數優化等問題。
2.3 人工智能領域的機器學習方法
目前常用的GS機器學習方法主要有支持向量機(support vector machine,SVM)、隨機森林(Random Forest,RF)、人工神經網絡(Artificial Neural Network,ANN)等[17-18]。機器學習不需要假設標記效應的分布和方差,依靠訓練和學習過程,挖掘事物內部存在的規律,考慮了標記間的線性及非線性關系,在某些性狀上優于傳統參數方法[17,19]。盡管如此,GS方法預測效果依然無法令人完全滿意,仍需根據不同的情況進行改進,并依據不同的標記、不同的性狀等選擇不同的方法。
3 GS的影響因素
影響GS預測準確性的主要因素有統計方法、標記密度和類型、性狀遺傳力的大小、訓練群體的大小、訓練群體和育種群體之間的關系、連鎖不平衡程度等。其基本公式表述如下[20]:
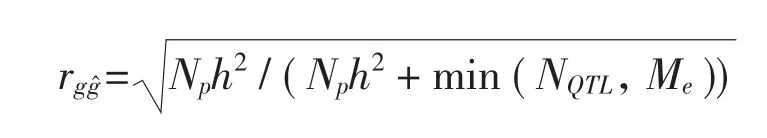
3.1 標記的選擇
GS的原理是利用與基因或QTL緊密連鎖的標記,用標記效應反映基因效應,因而篩選標記的方法主要有:根據LD值過濾冗余SNP或者選擇tag SNP或者block tag SNP,利用GWAS篩選與性狀顯著相關的significant SNP。另外,標記密度越高,預測的準確性越大;標記的密度主要決定于LD的跨度及樣本大小,異花授粉植物的LD跨度低于自花授粉,需要更高密度的分子標記,標記密度玉米>高粱>水稻[21]。當標記達到一定數目時,預測準確性處于平衡狀態,而后增加或減少標記數目,可能會降低準確性[22]。標記的類型對GS預測的準確性有很大的影響。單倍型是由數個相鄰SNP組成,更能捕獲QTL,優于SNP標記模型,但是當LD程度足夠大時,兩者之間沒差別[23]。但是SNP等位基因具有二態性,其結果的重復性較好,且使用起來更方便。
3.2 訓練群體
對于訓練群體的選擇主要考慮TP的遺傳結構、TP的大小、TP與BP的親緣關系遠近。TP的遺傳多樣性越豐富,預測準確性越高[24]。由于自交繁殖或者雙親雜交產生后代等因素,植物種群的遺傳基礎較動物狹窄,動物需要的訓練群體大于植物;自然群體樣本大于雙親群體。群體結構是影響GS預測準確性的一個重要因素。Isidro等[25]和Giovanny[26]闡述了基因組選擇過程中如何在考慮群體結構的情況下選擇最優訓練集,包括隨機抽樣法、基于PCA的相似法、最大決定系數平均值法、最小預測誤差方差平均值法、混合法等。結果表明,不同的作物、不同的性狀、不同的群體結構,最優訓練集的選擇標準不一樣。另外,TP和BP相關越強,預測的準確性越高。研究表明,對于多親本的小麥單交群體,以下群體預測的準確性從高到低依次為TP與BP來源于共同父母本、TP與BP共享雙親之一、TP與BP沒有共同雙親[27]。
3.3 統計方法
目前常用于GS預測的模型其準確性取決于個體間相關關系、分子標記效應及方差的假設分布。不同的群體、不同的性狀、不同的模型,其準確性不同。模擬數據研究表明,準確性BayesB>BayesA>RRBLUP[7]。在抗小麥銹病育種中,Bayesian Lasso和RRBLUP比SVM模型表現更好[28]。對于BLUP法,考慮到混合線性模型對具有較大效應的標記效應收縮得厲害,可以將此類標記作為固定效應,在一定程度上可以提高預測準確性。研究發現,將GWAS分析篩選出來的顯著與性狀相關的標記作為固定效應,放入GS模型中可以提高GEBV的準確性,尤其是GBLUP法[26,29-30]。另外,結合GWAS結果提供的信息構建標記權重對角矩陣,不同的SNP賦予不同的權重,并將所有標記分為兩組,一組位于已關聯到的QTL區域標記,一組為未關聯到的標記)也可以提高GEBV的準確性[31]。GS估計育種值往往只考慮加性效應,但是標記間可能存在一定的相關性,存在上位效應,并且對于F1群體,還需要考慮到顯性效應。當非加性遺傳方差存在時,在模型中如不加以考慮,將會高估加性效應。研究表明,當顯性方差占表型總方差的比例達到0.2,包含加性和顯性效應的預測模型顯著優于加性模型[32]。GBLUP和傳統的BLUP法分別從基因組和系譜的角度計算了個體間的相關關系,對于既有系譜關系又有基因組數據的群體,可以考慮將系譜和基因組數據相結合,完善關系矩陣,提高GEBV的準確性。從性狀表型的角度考慮,對于那些不符合正態分布或者離散型性狀,我們還可以考慮廣義混合線性模型用于提高預測的準確性。
4 基因組選擇在作物育種中的應用
自基因組選擇的概念提出以來,GS方法在動物育種中應用越來越廣泛,如小鼠、牛、雞等,尤其是奶牛,在全球各國的奶牛產業中均得到應用[33-36]。近年來,GS在植物育種中也逐漸引起了重視,已相繼開展了多種作物的GS模擬和驗證實驗,2007年Piyasatian等[37]用自交系雜交模擬了選擇效率,結果表明GS效率高于傳統的MAS。GS在玉米育種中使用最為廣泛,Riedelsheimer等[38]、Zhao等[39]、Guo[40]等在玉米自交、測交、雜交F1、RIL等群體中,使用RRBLUP模型,針對玉米籽粒含水量、產量、一般配合力(General Combining Ability,GCA)、代謝物含量等性狀,對個體GEBV進行了估計。GS在小麥育種中也有較多的涉獵,如Heffner等[22]的研究表明,在小麥雙親育種群體中,隨著高通量基因型分型技術的發展,GS較表型選擇和傳統MAS節約成本、縮短育種年限;Crossa等[41]對599個小麥品系產量進行GS分析,結果表明GS比傳統的BLUP預測能力提高了7.7%~35.7%;Zhao等[42]利用RRBLUP、BayesA、BayesB、BayesC以及BayesCπ方法,考慮加性和顯性效應,對90個小麥雜交種進行了GS分析。GS在水稻、大麥、甘蔗等其他作物中也有研究報道[43-45]。
基因組選擇方法的主要目的是對個體育種值進行準確的估計。針對不同的群體,GS還可以用于對所有遺傳變異和遺傳效應進行檢測和估計[46],以及雜種優勢的預測[47]。TP與BP親緣關系越近,GS預測準確性越高。對與TP間隔世代的BP進行預測,GS仍具有一定的預測力。研究表明,在與BP間隔3代以內,GS準確率每世代下降約5%[7],3代以后再重新估計標記效應。利用GS還可以對個體的適應性及穩產性進行估計。農業生產中對品種的評價,不僅需要考慮高產性,還需要考慮適應性及穩產型。GS在作物育種中的應用前景廣闊,但其發展仍處于初級階段,面臨著眾多挑戰,如不同的群體需要選擇不同的模型,基于基因與環境的互作、群體結構等因素需要改善模型等,這些都需要進一步完善。基于以上總結和探索,可以考慮構建一個GS平臺,一方面加強與各單位的合作,收集的豐富的表型和基因型數據;另一方面,可以收集已有的QTL信息,改善模型,將其融入GS分析當中。GS平臺的目的不僅用于育種值的估計,還可用作遺傳變異和效應估計、雜種優勢預測、品種評價。
[1]Xu Y B,Crouch J H.Marker-assisted selection in plant breeding:from publications to practice[J].Crop Science,2008,48(2):391-407.
[2]孔繁玲.植物數量遺傳學[M].北京:中國農業大學出版社,2006.
[3]Rex B.Molecular markers and selection for complex traits in plants:learning from the last 20 years[J].Crop Science,2008,48(5):1649-1664.
[4]Atwell S,Huang Y S,Vilhjalmsson B J,et al.Genome-wide association study of 107 phenotypes in a common set of Arabidopsis thaliana inbred lines[J].Nature,2010,465(7298):627-631.
[5]Huang X H,Zhao Y,Wei X H,et al.Genomewide association study of flowering time and grain yield traits in a worldwide collection of rice germplasm[J].Nature Genetics,2012,44(1):32-39.
[6]Zhu C S,Michael G,Yu J M,et al.Status and prospects of association mapping in plants[J].The Plant Genome,2008,1(1):5-20.
[7]Meuwissen T H E,Hayes B J,Goddard M E.Prediction of total genetic value using genomewide dense marker maps[J].Genetics,2001,157(4):1819-1829.
[8]Meuwissen T H E.Genomic selection:marker assisted selection on a genome wide scale[J].Journal of Animal Breeding and Genetics,2007,124(6):321-322.
[9]Goddard M E,Hayes B J.Mapping genes for complex traits in domestic animals and their use in breeding programmes[J].Nature Reviews Genetics,2009,10(6):381-391.
[10]Resende R M S,Casler M D,de Resende M D V.Genomic selection in forage breeding:accuracy and methods[J].Crop Science,2014,54(1):143-156.
[11]VanRaden P M.Efficient methods to compute genomic predictions[J].Journal of Dairy Science,2008,91(11):4414-4423.
[12]唐金梅,陳建國.全基因組選擇在植物育種中的研究進展[J].貴州農業科學,2016,44(8):1-5.
[13]王重龍,丁向東,劉劍鋒,等.基因組育種值估計的貝葉斯方法[J].Hereditas(Beijing),2014,36(2):111-118.
[14]Habier D,Fernando R L,Kizilkaya K,et al.Extension of the bayesian alphabet for genomic selection[J].BMC Bioinformatics,2011,12(1):1-12.
[15]Yi N J,Xu S Z.Bayesian LASSO for quantitative trait loci mapping[J].Genetics,2008,179(2):1045-1055.
[16]Moser G,Tier B,Crump R E,et al.A comparison of five methods to predict genomic breeding values of dairy bulls from genome-wide SNP markers[J].Genetics Selection Evolution,2009,41(1):1-16.
[17]Ornella L,Perez P,Tapia E,et al.Genomicenabled prediction with classification algorithms[J].Heredity,2014,112(6):616-626.
[18]Ogutu J O,Piepho H P,Schulz-Streeck T.A comparison of random forests,boosting and support vector machines for genomic selection[J].BMC Proceedings,2011,5(3):1-5.
[19]束永俊,吳磊,王丹,等.人工神經網絡在作物基因組選擇中的應用[J].作物學報,2011,37(12):2179-2186.
[20]Daetwyler H D,Pongwong R,Villanueva B,et al.The impact of genetic architecture on genomewide evaluation methods[J].Genetics,2010,185(3):1021-1031.
[21]Gupta P K,Rustgi S,Kulwal P L.Linkage disequilibrium and association studies in higher plants:present status and future prospects[J].Plant Molecular Biology,2005,57(4):461-485.
[22]Heffner E L,Jannink J L,Iwata H,et al.Genomic selection accuracy for grain quality traits in biparental wheat populations[J].Crop Science,2011,51:2597-2606.
[23]Calus M P L,Veerkamp R F.Accuracy of breeding values when using and ignoring the polygenic effect in genomic breeding value estimation with a marker density of one SNP per cM[J].Journal of Animal Breeding and Genetics,2007,124(6):362-368.
[24]Heslot N,Yang H P,Sorrells M E,et al.Genomic selection in plant breeding:a comparison of models[J].Crop Science,2012,52:146-160.
[25]Isidro J,Jannink J L,Akdemir D,et al.Training set optimization under population structure in genomic selection[J].Theoretical and Applied Genetics,2015,128(1):145-158.
[26]Giovanny C P.Genome-assisted prediction of quantitative traits using the R package sommer[J].Plos One,2016,11(6):1-15.
[27]Technow F,Schrag T A,Schipprack W,et al.Genome properties and prospects of genomic prediction of hybrid performance in a breeding program of maize[J].Genetics,2014,197(4):1343-1355.
[28]Ornella L,Sukhwinder S,Perez P,et al.Genomic prediction of genetic values for resistance to wheat rusts[J].The Plant Genome,2012,5(3):136-148.
[29]Rex B.Genomewide selection when major genes are known[J].Crop Science,2014,54(1):68-75.
[30]Abdollahi-Arpanahi R,Morota G,Valente B D,et al.Assessment of bagging GBLUP for wholegenome prediction of broiler chicken traits[J].Journal of Animal Breeding and Genetics,2015,132(3):218-228.
[31]Zhang Z,Ober U,Erbe M,et al.Improving the accuracy of whole genome prediction for complex traits using the results of genome wide association studies[J].Plos One,2014,9(3):1-12.
[32]de Almeida Filho J E,Guimaraes J F R,e Silva F F,et al.The contribution of dominance to phenotype prediction in a pine breeding and simulated population[J].Heredity,2016,117(1):33-41.
[33]Legarra A,Robert-Granie C,Manfredi E,et al.Performance of genomic selection in mice [J].Genetics,2008,180(1):611-618.
[34]Hayes B J,Bowman P J,Goddard M E,et al.Invited review:genomic selection in dairy cattle:progress and challenges[J].Journal of Dairy Science,2009,92(2):433-443.
[35]Chen C Y,Misztal I,Aguilar I,et al.Genomewide marker-assisted selection combining all pedigree phenotypic information with genotypic data in one step:an example using broiler chickens[J].Journal of Animal Science,2011,89(1):23-28.
[36]Loberg A,Durr J W.Interbull survey on the use of genomic information[J].Interbull Bull,2009,39:3-14.
[37]Piyasatian N,Fernando R L,Dekkers J C M.Genomic selection for marker-assisted improvement in line crosses[J].Theoretical and Applied Genetics,2007,115(5):665-674.
[38]Riedelsheimer C,Czedik-EysenbergA,Grieder C,et al.Genomic and metabolic prediction of complex heterotic traits in hybrid maize[J].Nature Genetics,2012,44(2):217-220.
[39]Zhao Y S,Gowda M,Liu W X,et al.Accuracy of genomic selection in European maize elite breeding populations[J].Theoretical and Applied Genetics,2012,124(4):769-776.
[40]Guo T T,Li H H,Yan J B,et al.Performance prediction of F1 hybrids between recombinant inbred lines[J].Theoretical and Applied Genetics,2013,126(1):189-201.
[41]Crossa J,de losCampos G,Perez P,et al.Prediction of genetic values of quantitative traits in plant breeding using pedigree and molecular markers[J].Genetics,2010,186(2):713-724.[42]Zhao Y S,Zeng J,Fernando R L,et al.Genomic prediction of hybrid wheat performance[J].Crop Science,2013,53(3):802-810.
[43]Xu S Z,Zhu D,Zhang Q F.Predicting hybrid performance in rice using genomic best linear unbiased prediction[J].PNAS,2014,111(34):12456-12461.
[44]Zhong S Q,Dekkers J C M,Fernando R L,et al.Factors affecting accuracy from genomic selection in populations derived from multiple inbred lines:a barley case study[J].Genetics,2009,182(1):355-364.
[45]Gouy M,Rousselle Y,Bastianelli D,et al.Experimental assessment of the accuracy of genomic selection in sugarcane[J].Theoretical and Applied Genetics,2013,126(10):2575-2586.
[46]Vitezica Z G,Varona L,Legarra A.On the additive and dominant variance and covariance of individuals within the genomic selection scope[J].Genetics,2013,195(4):1223-1230.
[47]Zhao Y S,Li Z,Liu G Z,et al.Genome-based establishment of a high-yielding heterotic pattern for hybrid wheat breeding[J].PNAS,2015,112(51):15624-15629.
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
兒童故事畫報(2019年5期)2019-05-26 14:26:14
中學物理·高中(2016年12期)2017-04-22 11:53:03
Coco薇(2016年2期)2016-03-22 02:42:52
Coco薇(2015年1期)2015-08-13 02:47:34
小雪花·成長指南(2015年7期)2015-08-11 15:03:12
小雪花·成長指南(2015年4期)2015-05-19 14:47:56
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
