朱砂葉螨危害對木薯幼苗光合作用及葉綠素熒光參數的影響
2018-01-16 18:38:16梁瓊月董蒙蒙顧明華
廣東農業科學 2017年9期
梁瓊月,董蒙蒙,何 冰,顧明華,陳 立,楊 娟,賴 艷,曾 瑋
(1.廣西大學農學院,廣西 南寧 530004;
2.廣西大學植物科學國家級實驗教學示范中心,廣西 南寧 530004)
木薯(Manihot esculentaCrantz)被稱為世界三大薯類作物之一,除了作為糧食食用以外,其另一大用處為生物燃料——乙醇的生產原料。近年來,木薯栽培遭到以朱砂葉螨(Tetranychus cinnabarinus)為首的螨蟲危害嚴重,其中華南等地區木薯朱砂葉螨危害成災,嚴重時可致木薯產量降低50%~70%[1]。朱砂葉螨民間稱之為紅蜘蛛,是遍布全世界的害蟲[2],木薯植株受取食后將出現生長發育不良,減產嚴重,甚至顆粒無收。國內外對木薯害螨的研究熱點集中于木薯單爪螨(Mononychellus tanajoa)方面[3-6],國內偏向于防控檢疫性害螨,如木薯單爪螨侵入危害方面[7-13],關于朱砂葉螨取食危害與寄主木薯的光合作用生理反應的研究鮮見報道。
植物生長過程中必須依靠光合作用來提供能量物質。光合作用作為植物生理學方面研究的關鍵性問題[14],這一生理過程極易受外界環境變化的影響[15]。葉綠素熒光參數是植物光合作用和環境的關系的內在反映,從更深程度解析植物進行光合作用時光系統對光能的吸收到利用的整個過程深入反映其內在本質特征[16-17]。因此,我們研究了朱砂葉螨危害對木薯4個品種(華南205、南植199、桂熱891、華南8號)幼苗光合作用和葉綠素熒光參數的影響,以期為木薯朱砂葉螨的防治及抗性品種的篩選提供理論依據。
1 材料與方法
1.1 試驗材料
供試植物為盆栽木薯4個品種:華南8號、南植199、桂熱891、華南205。各品種木薯分別種植于塑料盆(上口徑 15 cm)內,每盆2株,以無菌泥炭土為種植基質,用防蟲罩各盆隔離栽培于玻璃溫室內,溫度28(±2)℃,相對濕度70(±5)%,水分、肥料管理按常規栽培進行,植株生長至7~9片完全展開葉時進行試驗處理。
供試昆蟲為朱砂葉螨,于2016年5月在武鳴采集,隔離飼養于室內。試驗蟲齡為2~3 齡蟲,從飼養室內繼代種群中隨機挑取,用毛筆將其移接至試驗木薯正4~8葉片上。
1.2 試驗方法
2016年4月20日栽種木薯,6月15日于每個木薯品種中選取未受螨蟲或其他病蟲危害的3盆(每盆1株)長勢相當的木薯健康植株,接種蟲量10頭/株,隨機釋放,讓其自由取食葉片,接種后的各木薯植株分別采用防蟲罩隔離栽培。每個處理3次重復。分別在接種螨蟲后5、10 d對木薯葉片進行光合作用相關指標和葉綠素熒光參數的測定,以健康植株為對照。試驗地點為廣西大學農學院教學科研基地玻璃網室內。
光合作用的測定采用便攜式光合作用測定儀(LI-6400XT,LI-COR,美國)分別對木薯葉片(正5葉,功能葉)光合相關指標進行測定,測定時,設定光強PAR為1 500 umol/m2·s,溫濕度采用自然狀態。試驗采用不同處理交替進行測量避免環境因素變化影響測量結果。試驗時間在6月1~7日上午9:00~11:00,天氣晴朗無云。
葉綠素熒光參數測定采用葉綠素熒光分析儀(PAM-2500,WALZ,德國)分別對木薯葉片(正5葉,功能葉)葉綠素熒光參數進行測定。試驗時間在6月1~7日凌晨。
試驗結果采用Excel、SPSS軟件進行處理分析。
2 結果與分析
2.1 朱砂葉螨危害對木薯葉片氣體交換參數的影響
由圖1可知,供試木薯品種受朱砂葉螨危害后葉片凈光合速率(Pn)、氣孔導度(Gs)和蒸騰速率(Tr)均低于健康植株,差異顯著,且隨著受害時間的延長呈下降趨勢;而胞間CO2濃度(Ci)則反之,受害植株均高于正常健康植株,且受害時間越長其差異也越明顯。4個不同基因型木薯在受害10 d后,氣體交換參數的變化趨勢一致,其中受害葉片Pn、Gs和Tr比健康植株降低最為顯著的是桂熱891,分別降低91.3%、95.0%和80.1%,而胞間Ci則以華南205上升最顯著,達45.2%。
2.2 朱砂葉螨危害對木薯葉片葉綠素熒光參數的影響
由圖2可知,木薯對朱砂葉螨的危害的反應并非因品種而異,受危害后F0含量均呈現上升趨勢,兩個不同時間段含量均顯著高于健康植株;而Fm值則有所下降,受危害后5 d以華南205下降最為顯著,其他各品種與健康植株未達差異顯著水平,受害后10 d 4個品種受害植株與健康植株間Fm值未達差異顯著水平;同時Fv/Fm也下降,變化趨勢與Fm一致;隨著木薯的生長發育,F0和Fm有所降低。同時本試驗4個不同基因型木薯在受害后10 d,各受害植株間、各健康植株間葉綠素熒光參數F0,Fm和Fv/ Fm均未達差異顯著水平。有研究認為F0值增大說明光系統Ⅱ中RC受損壞,或永久失活。通常植株受到脅迫,F0值增大,本研究結果也顯示木薯植株受危害后F0值有所上升。可見F0、Fm和Fv/Fm值在植物生長發育過程中并非簡單的增加或降低,其變化趨勢因植物種類而異。
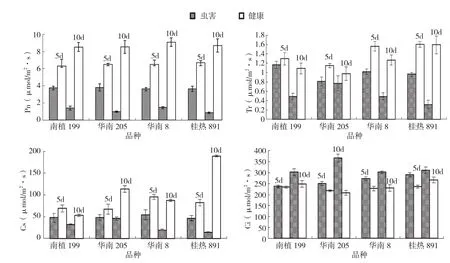
圖1 朱砂葉螨危害對木薯葉片氣體交換參數的影響

圖2 朱砂葉螨危害對木薯葉片葉綠素熒光參數的影響
3 結論與討論
植物生長發育過程中,光合作用在生理代謝進程中發揮極其重要的作用,光合水平直接影響植物的生長發育及其抗逆性[18]。植物光合作用過程中對二氧化碳和水的運輸及調控是通過氣孔完成,氣孔限制或非氣孔限制均直接致使光合能力降低。植物葉片Ci是評估其光合作用是受氣孔或非氣孔限制的關鍵參數,Ci降低為氣孔限制,Ci 上升為非氣孔限制[19]。同時前人發現,植物受病蟲侵害后,致使葉片Gs值上升,Pn減弱,如昆蟲危害云杉[20]、葉甲危害紅荊[21];同時也有研究表明植株受蟲害導致葉片Gs降低,如鱗翅目取食蘋果樹[22]、癭螨危害糖楓樹[23]和纓翅目危害梨樹[24]。而二斑葉螨對棉花危害初期Gs降低,后期Gs上升[25]。可見,病蟲危害對植物光合指標的影響因植物種類、危害時期和強度而異。本研究中,不同木薯品種受朱砂葉螨取食危害后,葉片的Pn、Gs均降低,而Ci增加,可見Pn降低可能是由于非氣孔限制引起的葉肉細胞同化能力降低。
植物長期處于較嚴重的脅迫狀態,光合強度受限多為非氣孔限制所致,光合代謝過程受抑制和損傷,如葉片光合放氧能力、葉綠體Hill反應和葉綠素熒光強度顯著降低等[19]。葉綠素熒光參數反映了光合系統PSⅡ的電子傳遞吸收光化學量子效率。光系統Ⅱ的光能轉化效率是反映植物潛在光合效率高低的指標,常被用于評價植物受環境脅迫程度的指標。有研究表明不同植物種類受病蟲危害后,葉片Fv/Fm值因植物種類而異,如綠斑病藻侵染夏橙[26]、黃瓜花葉病毒侵染煙草[27]均降低了Fv/Fm值;而鱗翅類幼蟲取食冬青[28]、茶尺蠖取食茶樹[29]和大蝸牛危害黃瓜[30]后Fv/Fm值則升高。Fo和Fm表示光系統Ⅱ反應中心處于完全打開或完全關閉狀態的熒光產量,PS反應中心受損壞壞或可逆失活致使F0值上升,而PS熱能耗散增多F0值則降低。本研究結果表明,供試木薯品種受朱砂葉螨危害后Fv/Fm值降低,且隨著危害時間的延長而下降,說明在朱砂葉螨的危害下木薯葉片發生了光抑制或PSⅡ損害,同時F0水平呈上升的趨勢,說明光抑制或PSⅡ損害程度較嚴重。
從本研究可見,朱砂葉螨危害對木薯光合機制產生一定的影響,部分氣體交換參數和葉綠素熒光參數值降低,凈光合速率降低主要是由于非氣孔限制所致,葉綠素熒光參數值的變化可能是木薯葉片發生光抑制或PSⅡ損害導致。
[1]陳青,盧芙萍,黃貴修,等.木薯害蟲普查及其安全性評估[J].熱帶作物學,2010,31(5):819-827.
[2]李隆術,李云瑞.蜱螨學[M].重慶:重慶出版社,1988:4-129.
[3]Henrik Skovg?rd,Jonna Tomkiewicz,G?sta Nachman,et al.The effect of the cassave green miteMononychellus tanajoaon the growth and yeild of cassavaManihot esculentain a seasonally dry area in Kenya[J].Experimental & Applied Acarology,1993(17):41-58.
[4]Akinlosotu T A.Seasonal trend of green spider mite,Mononychellus tanajoapopulation on cassava,Manihot esculentaand its relationship with weather factors at moor plantation[J].Insect Sci.Application,1982,3(4):251-254.
[5]Costa E C,Teodoro A V,Rêgo A S,et.al.Population structure and dynamics of the cassava green miteMononychellus tanajoa(Bondar)and the predatorEuseius ho(DeLeon)(Acari:Tetranychidae,Phytoseiidae)[J].Arthropods,2012,1(2):55-62.
[6]Omorusi V I,Ayanru D K G.Effect of NPK fertilizer on diseases,pests and mycorrhizal symbiosis in cassava[J].Int.J.Agric.Biol,2011(13):391-395.
[7]盧輝,陳青,盧芙萍,等.基于Maxent模型的木薯單爪螨在云南的適生性評價(英文)[J].Agricultural Science & Technology,2011,12(12):1905-1908.
[8]盧輝,陳青,盧芙萍,等.木薯單爪螨全球潛在地理分布的MaxEnt預測[J].植物檢疫,2012,26(1):1-6.
[9]經福林,李遷,盧芙萍,等.木薯單爪螨的寄主選擇性研究[J].熱帶作物學,2013,34(2):344-347.
[10]盧芙萍,符悅冠,黃貴修,等.溫度對木薯單爪螨生長發育與繁殖的影響[J].熱帶作物學報,2011,32(9):1720-1724.
[11]盧芙萍,符悅冠,經福林,等.卵高溫脅迫對木薯單爪螨發育與繁殖的影響研究[J].中國農學通報,2012,28(21):229-236.
[12]黃潔,范偉峰,劉茂,等.531份木薯種質的朱砂時螨抗性評價[J].廣東農業科學,2011,38(2):27-28.
[13]韋威旭,覃維治,韋民政,等.木薯品種對二點紅蜘蛛的抗性觀察[J].廣西農業科學,2009,40(5):504-506.
[14]Nabity P D,Zavala J A,Lawlor D W.Musings about the effects of environment on photosynthesis[J].Annals of Botany,2009,103:543-549.
[15]Samsone I,Andersone U,Ievinsh G.Variable effect of arthropod-induced galls on photochemistry of photosynthesis,oxidative enzyme activity and ethylene production in tree leaf tissues[J].Environmental and Experimental Biology,2012,10:15 - 26.
[16]李曉,馮偉,曾曉春.葉綠素熒光分析技術及應用進展[J].西北植物學報,2006,26(10):2186-2196.
[17]陳建明,俞曉平,程家安.葉綠素熒光動力學及其在植物抗逆生理研究中的應用[J].浙江農業學報,2006,18 (1):51-55.
[18]惠紅霞,許興,李前榮.外源甜菜堿對鹽脅迫下枸杞光合功能的改善[J].西北植物學報,2003,23(12):2137 -2422.
[19]FarquharG D,SharkeyTD.Stomatal conductance and photosynthesis[J].AnnualReview PlantPhysiology and PlantMolecular,1982(33):317.
[20]Ozaki K,Saito H,Yamamuro K.Compensatory photosynthesis as a response to partial debudding in ezo spruce,Picea jezoensis seedlings[J].Ecol Res,2004,19(2):225-231.
[21]Pattison R R,D'Antonio C M,Dudley T L.Biological control reduces growth,and alters water relations of the saltcedar tree(Tamarixspp.)in western Nevada,USA[J].J Arid Environ,2011,75(4):346-352.
[22]Pincebourde S,Frak E,Sinoquet H,et al.Herbivory mitigation through increased wateruse efficiency in a leaf-mining moth-apple tree relationship[J].Plant Cell Environment,2006,29(12):2238-2247.
[23]Patankar R,Thomas S C,Smith S M.A gallinducing arthropod drives declines in canopy tree photosynthesis[J].Oecologia Nov,2011,167(3):701-709.
[24]Ellsworth D S,Tyree M T,Parker B L,et al.Photosynthesis and water-use efficiency of sugar maple(Acer saccharum)in relation to pear thrips defoliation[J].Tree Physiol,1994,14(6):619-632.
[25]Reddall A,Sadras V O,Wilson L J,et al.Physiological responses of cotton to two-spotted spider mite damage[J].Crop Sci,2004,44(3):835-846.
[26]王大平,曾明,朱鈞,等.綠斑病藻寄生對夏橙葉片光合作用特性的影響[J].應用生態學報,2006,17(6):1141-1144.
[27]王春梅,施定基,朱水芳,等.黃瓜花葉病毒對煙草葉片和葉綠體光合活性的影響[J].植物學報,2000,42(4):388-392.
[28]Zhang B,Zhou X,Zhou L,et al.A global synthesis of below-ground arbon responses to biotic disturbance:a meta-analysis[J].GlobalEcology and Biogeography,2014,24(2):126-138.
[29]韋朝領,童鑫,高香鳳,等.茶樹對茶尺蠖取食危害的補償光合生理反應研究[J].安徽農業大學學報,2007,34(3):355-359.
[30]Thomson V,Cunningham S,Ball M,et al.Compensation for herbivory by Cucumis sativus through increased photosynthetic capacityand efficiency[J].Oecologia,2003,134(2):167-175.
猜你喜歡
今日農業(2022年15期)2022-09-20 06:56:12
少兒科學周刊·少年版(2022年20期)2022-05-30 10:48:04
海峽姐妹(2020年12期)2021-01-18 05:53:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
創新作文(小學版)(2017年34期)2017-04-09 06:15:18
爆笑show(2016年7期)2017-02-09 09:36:13
民生周刊(2016年9期)2016-05-21 12:11:19
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
