辣椒熱激轉錄因子家族在馴化過程中的演變
2018-01-23 08:42:52龔記熠孫媛媛
西北大學學報(自然科學版) 2018年1期
關鍵詞:植物
李 菲,龔記熠,孫媛媛,乙 引
(貴州師范大學 生命科學學院/貴州省植物生理與發育調控重點實驗室, 貴州 貴陽 550001)
不能移動的植物生存在變動的環境中,需要應對各種不利的環境條件。目前針對植物抗性的研究,往往聚焦于某一單一逆境。轉錄組學分析發現,61%的雙重脅迫誘導的基因在任何一個單一脅迫環境下都不會誘導表達[1]。因此,有必要研究植物響應多重脅迫的機制[2-3]。
植物應對多種逆境的方法之一是利用多效熱激轉錄因子(Heat Shock Transcription Factors,HSFs)——熱激蛋白/分子伴侶(heatshock protein/chaperone)信號傳導和基因調控網絡。逆境影響植物正常生長的分子機制,是阻礙細胞內蛋白質正常進行生理功能。蛋白質變性是任何逆境都會引發的后果。所以,任何會引起蛋白質錯誤折疊的逆境,都需要植物基因組中熱激轉錄因子(HSFs)信號傳導和基因調控網絡來應對[4]。熱激轉錄因子(HSFs)信號傳導途徑的作用不僅限于熱應激響應,而且還涉及多種脅迫,如冷、滲透、干旱、鹽、紫外線、高光、氧化和病原體侵染脅迫[5]。
HSF存在于所有真核生物中,但與釀酒酵母中的1個HSF或人類中7個HSFs基因相比,植物基因組中普遍存在大量HSFs基因(大豆中38個,水稻中25個,擬南芥21個)[6]。植物中HSF家族的多樣性造成了研究的困難。然而,表達模式、基因突變和過表達研究顯示了擬南芥HSFs家族基因在抗多種逆境脅迫和發育中的獨特和重疊的功能[7]。
熱激轉錄因子(HSFs)分為A,B和C3類[8-10]。大量的HSFs成員基因及其以異源多聚體的形式發揮功能的特征,使得特定基因的功能研究非常困難。多個HSFs基因在植物干旱、鹽、強光、冷脅迫等的信號傳導和基因表達調控中起作用。它們可以被ABA依賴性或非依賴的信號通路誘導。HSFA3是DREB2A轉錄因子)的靶點,由多種脅迫如干旱、鹽、熱和冷以及氧化脅迫、紫外光UVB和傷害誘導[11-12]。AtHSFA6a的啟動子包含兩個ARE(ABA反應元件),并在體外結合3種ABA響應轉錄因子:AREB1,ABF3和AREB3[13]。過表達AtHSFA6a的植物表現出抗鹽脅迫和抗旱能力。B類和C類HSFs基因缺少A類HSFs基因調節轉錄活性所必需的結構域AHA (aromatic hydrophobic acidic),因此被認為是抑制性轉錄因子。
在非模式植物中,HSFs轉錄因子的功能也有研究[14]。辣椒營養豐富,是我國西南省份重要的經濟作物。利用重要的栽培辣椒品種遵辣一號(C.apsicumannuumLZunla1),及其野生祖先品系Chiltepin(C.annuumvar.glabriusculum)的基因組測序和注釋結果。本研究分析了野生辣椒和栽培辣椒基因組中HSFs基因家族的成員組成、系統分類和可能功能,以及馴化過程中辣椒基因組中HSFs基因家族發生的演變。本研究為辣椒品系應對多種逆境的機制研究和品種改良提供了基因資源。
1 材料與方法
1.1 擬南芥HSFs基因家族序列和辣椒基因組序列來源
擬南芥HSFs基因家族序列來源于擬南芥數據庫TAIR。栽培辣椒遵辣一號和其野生祖先品系Chiltepin的基因組序列及其注釋信息可以從NCBI Genome數據庫下載。
1.2 辣椒基因組中HSFs基因的檢索和篩選
辣椒基因組中HSFs基因的檢索工具是本地BLAST軟件。將下載到的栽培辣椒遵辣一號和其野生祖先品系Chiltepin的基因組序列及其注釋信息,構建本地核苷酸和氨基酸數據庫。以擬南芥HSFs家族全部序列信息為檢索序列,進行本地BLAST。保留檢索到的E值小于0001的序列。
下一步對這些序列進行結構域分析。結構域分析是在Pfam在線蛋白質結構域數據庫中,進行這些序列的結構域檢索。保留含有HSFs特征性結構域:HSF-DNAbind結構域的序列,用于后續分析。
1.3 栽培辣椒和野生辣椒HSFs基因家族的染色體定位比較
栽培辣椒和野生辣椒HSFs基因家族成員的染色體定位信息可以根據基因序列號從基因組序列信息中查詢到。根據基因的染色體定位信息,使用MapInspect軟件分別構建出栽培辣椒和野生辣椒HSFs基因家族成員的染色體定位圖譜,并進行比對和解讀。
1.4 辣椒基因組中HSFs基因的系統進化分析
對擬南芥、栽培辣椒遵辣一號和野生辣椒品系Chiltepin 全部HSFs基因序列進行序列比對和系統進化樹構建,使用的軟件是MEGA70。序列比對使用的算法是Clustal W。系統發育樹構建使用的算法是Neighbor Joining。
2 結果與討論
2.1 辣椒基因組中HSFs基因的鑒定和特征初步分析
在西南各省廣泛種植的栽培辣椒品種遵辣一號(C.apsicumannuumLZunla1),和其野生祖先品系Chiltepin(C.annuumvar.glabriusculum)的基因組測序已經完成,序列及注釋信息均公開(http://peppersequencegenomicscn/page/species/indexjsp)。使用擬南芥HSFs基因家族的基因序列,可以從辣椒基因組中挖掘出其中的HSFs基因。
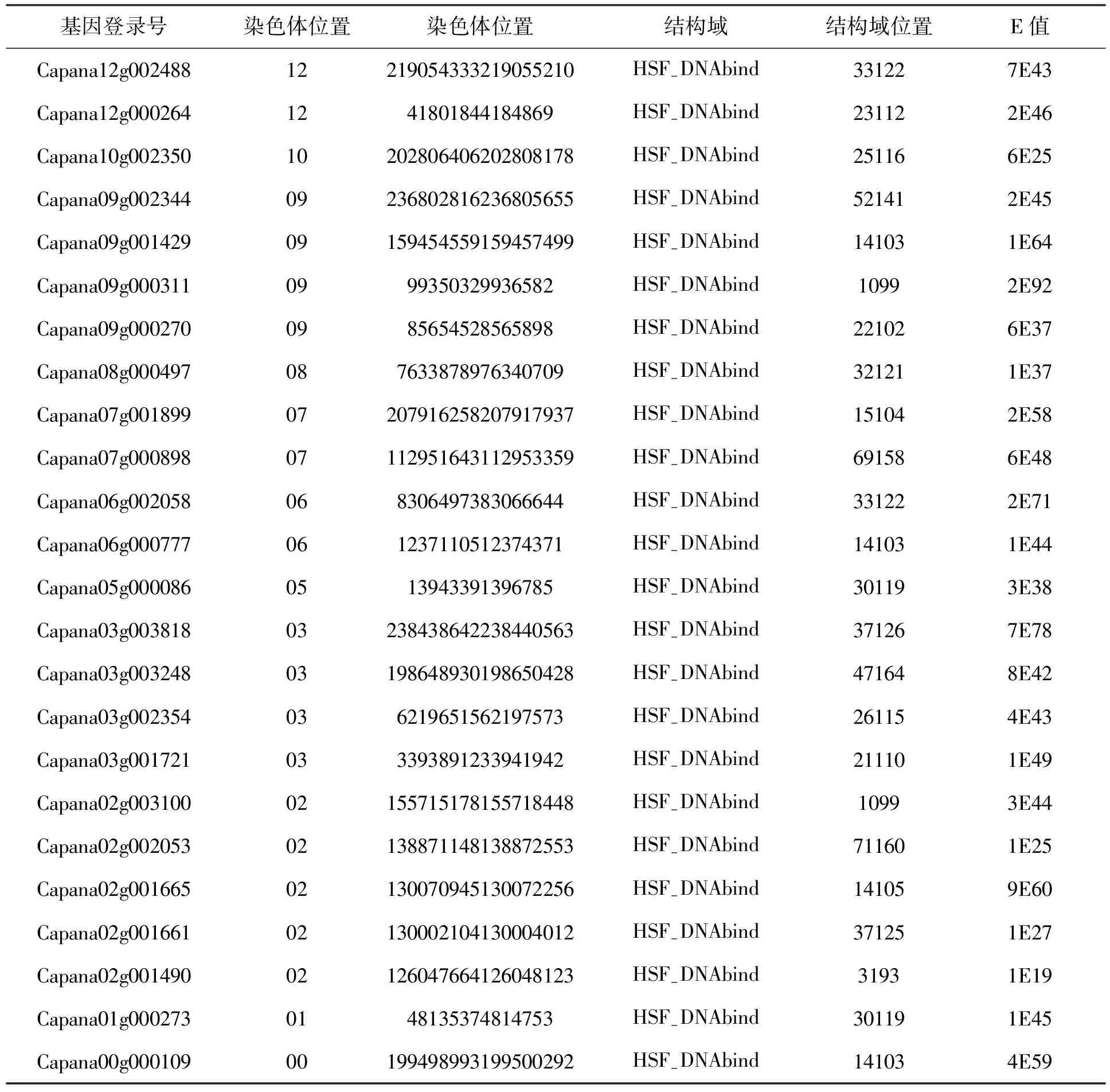
表1 遵辣一號(C. apsicum annuum LZunla1)基因組中HSFs基因信息列表Tab.1 The HSFs genes in the C. apsicum annuum L Zunla1 genome
在栽培辣椒遵辣一號基因組中共鑒定出25個HSFs基因,其野生祖先品系Chiltepin中鑒定出26個HSFs基因。這一HSFs基因家族成員數目與其他植物(擬南芥21,水稻25個,大豆38個)相差不多(Fujimoto and Nakai 2010),但比其他生物,如動物和酵母多。這應該與植物必須靜止應對多種逆境脅迫有關。辣椒基因組經過重復序列的擴增,比其他植物大很多(辣椒3364Mbp;擬南芥125Mbp;水稻466Mbp; 大豆1100Mbp)。但是,基因組中HSFs功能基因的數量,辣椒并沒有比其他植物更多。
2.2 栽培辣椒和野生辣椒品系基因組中HSFs基因的染色體定位
栽培辣椒品種遵辣一號(C.apsicumannuumLZunla1),和其野生祖先品系Chiltepin(C.annuumvar.glabriusculum)的農藝性狀,包括株高、株型、果實大小等有巨大的差異,在抗性方面也有不同的表現。從HSFs基因染色體位置看,與其野生祖先品系Chiltepin比較,遵辣一號HSFs基因家族基本一致。大部分HSFs基因在馴化過程中沒有發生染色體位置變化。在遵辣一號品種一號和七號染色體上,分別丟失了1個HSFs基因(Capang01g003145和Capang07g001960)。遵辣一號基因組中有1個HSF基因,在注釋過程中未能定位到任何一個染色體上(Capana00g000109)。這個基因和Chiltepin野生辣椒品系的Capang04g002355基因在系統發育樹中聚集在一個分支,與Chiltepin基因組中Capang01g003145和Capang07g001960并沒有相近的親緣關系。
2.3 栽培辣椒和野生辣椒品系基因組中HSFs基因的系統發育分析
對于HSFs基因家族的功能,研究的最透徹的還是擬南芥。將擬南芥、栽培辣椒遵辣一號和野生辣椒Chiltepin的全部HSFs基因的氨基酸序列進行系統進化分析,如圖2。
與擬南芥一樣,辣椒HSFs基因家族分成了A/B/C3個亞家族。從系統發育樹中,可以清晰地看到辣椒HSFs基因在擬南芥中的同源基因。遵辣一號的HSFs基因,一般與Chiltepin的HSFs基因一一對應,位于同一分支。少數幾個例外包括:Capana00g000109和Chiltepin野生辣椒品系的Capang04g002355在系統發育樹中聚集在一個分支;Chiltepin野生辣椒品系的Capang07g001960基因在遵辣一號中沒有對應基因;遵辣一號的Capana08g000497 與Chiltepin野生辣椒品系的Capang01g003145基因聚集在一個分支。
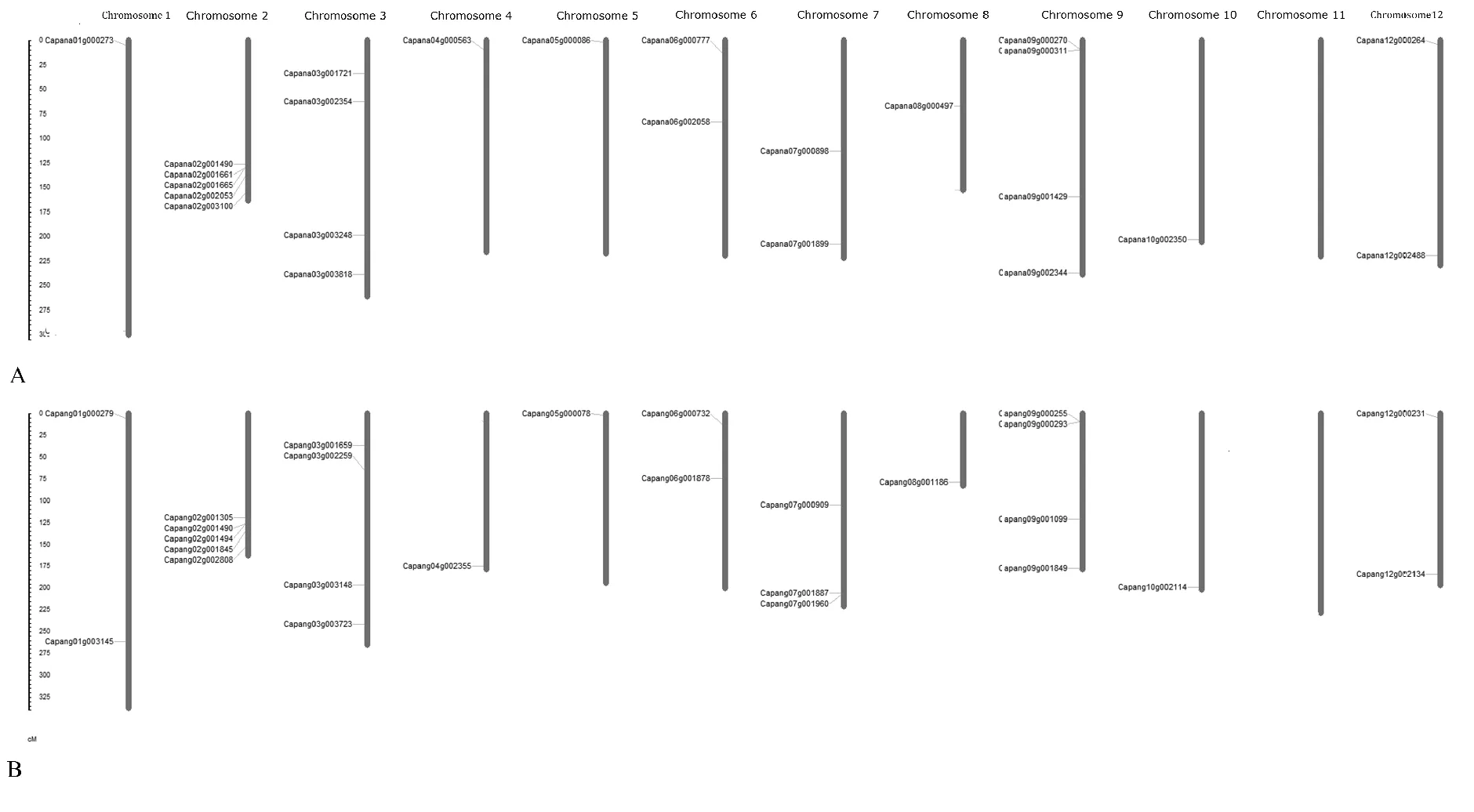
圖1 栽培辣椒遵辣一號” Zunla1”和野生辣椒品系” Chiltepin”基因組中HSFs基因的染色體定位圖譜Fig.1 Chromosomal location map of HSFs gene in the genome of the cultivated pepper "Zunla1" and its wild ancestor "Chiltepin"
2.4 栽培辣椒和野生辣椒品系基因組中HSFs基因的可能功能
HSFs基因家族成員,尤其是HSFAs亞家族的成員,功能研究的報道已經有很多。系統發育分析揭示了與擬南芥HSFAs基因同源的辣椒基因。這為對辣椒HSFs基因進行功能研究提供了很好的線索。
在擬南芥中,熱激轉錄因子HSFA1abd和e組成型表達,負責觸發熱應激反應(HSR,heat stress response)[11]。這些轉錄因子與熱激蛋白啟動子區域的熱應激因子(HSE heat stress elements)元件結合以激活熱激蛋白HSP的轉錄。此外,這些基因也可激活其他熱激轉錄因子,包括HSFA2、HSFA3和HSFA7a的表達,賦予植株長時間的脅迫抗性[15]。這些信號通路中,HSFA2處于重要的中心位置。已經充分證明,HSFA2的過表達可增強植株對熱、缺氧、鹽、滲透脅迫、強光和氧化脅迫的抵抗力[16]。
與擬南芥HSFA2基因同源的是遵辣一號的Capana08g000497 與Chiltepin野生辣椒品系的Capang01g003145基因。遵辣一號的Capana08g000497 與Chiltepin的Capang01g003145兩個基因相似度為100%。與擬南芥HSFA2基因相似度為89%。鑒于HSFA2基因對于擬南芥多種抗性的重要作用,研究遵辣一號的Capana08g000497 與Chiltepin的Capang01g003145基因在辣椒中的功能,具有重要的意義。
這一重要的抗逆基因,在馴化過程中,由Chiltepin一號染色體移動到遵辣一號的八號染色體。這一基因移位發生的時間和機制、基因移位對該基因表達的影響,以及對栽培辣椒抗逆性的影響,是研究栽培辣椒抗逆性的關鍵問題。
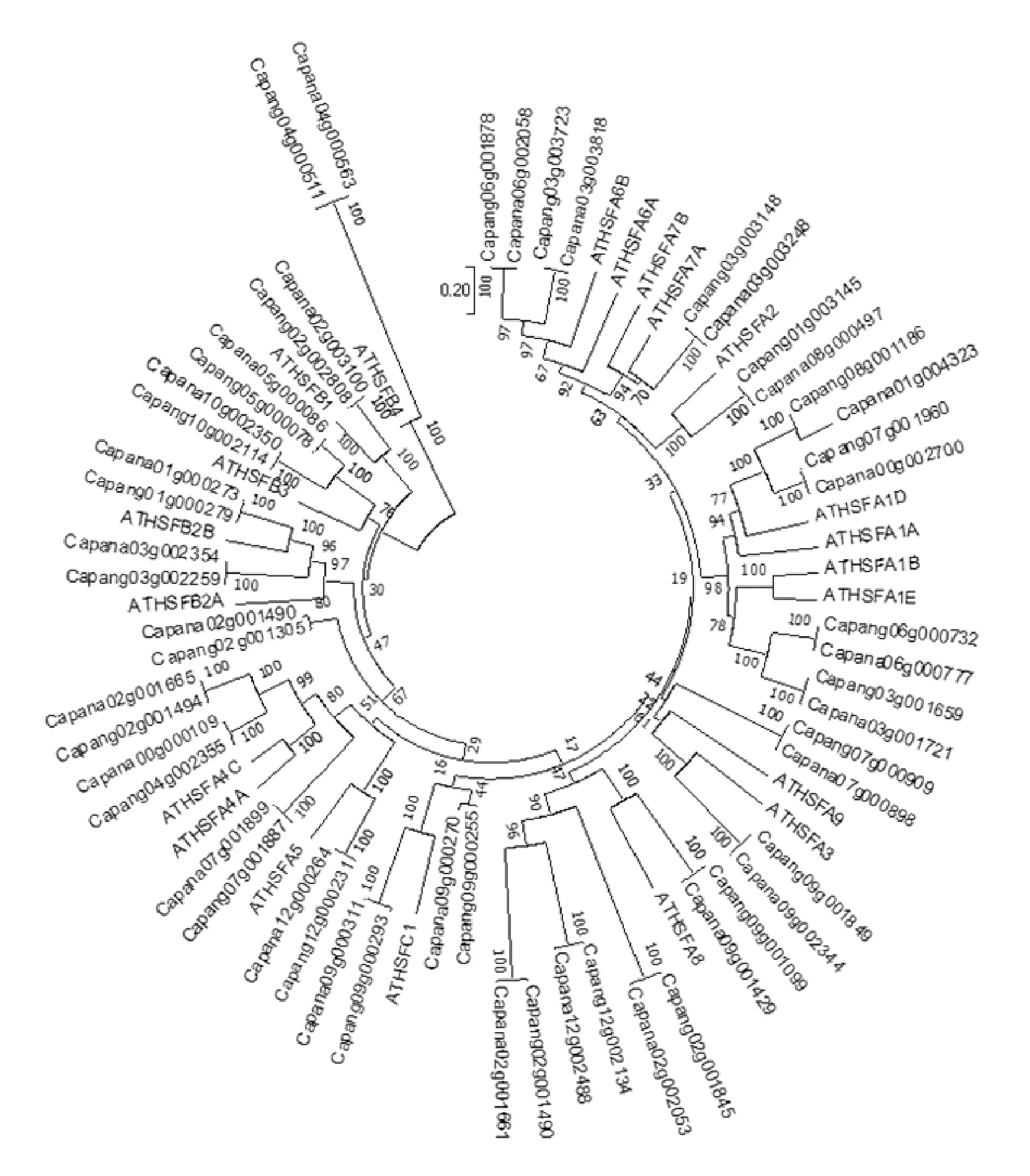
圖2 栽培辣椒遵辣一號” Zunla1”和野生辣椒品系” Chiltepin”基因組中HSFs基因與擬南芥HSFs基因家族的系統進化分析Fig.2 Phylogenetic analysis of HSFs gene in the genome of Arabidopsis, the cultivated pepper "Zunla1" and its wild ancestor "Chiltepin"
3 結 論
氣候災害環境給農作物生產帶來嚴重的風險,極大的危害農業經濟效益的穩定性。目前對植物抗逆的研究,主要集中在對單個逆境的研究。這很難解決實際生產中遇到的多種逆境同時發生的情況。植物基因組中的熱激轉錄因子——熱激蛋白信號傳導和基因表達調控途徑,是植物在各種脅迫環境下,均會激活以便維持細胞內蛋白質正常生理功能的重要抗逆機制。本文以研究相對透徹的擬南芥HSFs基因家族已知信息為基礎,挖掘辣椒栽培品種遵辣一號(CapsicumannuumLZunla1),及其野生祖先品系Chiltepin(C.annuumvar.glabriusculum)基因組中HSFs基因,并對這些基因進行系統的比對和功能分析。
盡管辣椒基因組遠大于擬南芥,其基因組中HSFs基因的數目并沒有更多。對辣椒和其他茄科植物,如番茄、茄子等的基因組進行比較,發現辣椒基因組中重復序列發生了大規模的擴增,但是基因數目并沒有增加[17]。對辣椒基因組中HSFs基因家族的分析,為此論斷提供一個新的例證。
在栽培辣椒和其野生祖先辣椒品系基因組中,HSFs基因家族比較保守。栽培辣椒遵辣一號(C.apsicumannuumLZunla1)基因組中,HSFs基因家族成員的數目、染色體位置和序列,與其野生祖先大體上保持了一致。但是,可提高擬南芥對多種逆境抗性的重要基因,HSFA2,在遵辣一號及其野生祖先Chiltepin中的同源基因發生了移位。這一基因移位對于栽培辣椒與其野生祖先抗逆性的差異之間的關聯,是值得研究的問題。本研究為分析辣椒HSFs基因家族成員的功能提供了有價值的信息,為辣椒品種多種抗逆能力的改善,積累了基因資源。
[1] RASMUSSEN S, BARAHarah P, SUAREZRODRIGUEZ M C, et al. Transcriptome responses to combinations of stresses in arabidopsis[J]. Plant Physiology, 2013, 161(4):1783-1794.
[2] MITTLER R, BLUMWALD E. Genetic engineering for modern agriculture: Challenges and perspectives[J]. Annual Review of Plant Biology, 2010, 61(1):443-462.
[3] NOBUHIRO SUZUKI, RIVERO R M, VLADIMIR S, et al. Abiotic and biotic stress combinations[J]. New Phytologist, 2014, 203(1):32-43.
[4] CAREY C C, GORMAN K F, RUTHERFORD S. Modularity and intrinsic evolvability of Hsp90-buffered change[J]. Plos One, 2006, 1(1):76.
[5] SWINDELL W R, HUEBNER M, WEBER A P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways[J]. Bmc Genomics, 2007, 8(1):125.
[6] FUJIMOTO M, NAKAI A. The heat shock factor family and adaptation to proteotoxic stress[J].Febs Journal, 2010, 277(20):4112-4125.
[7] VON K P, SCHARF K D, NOVER L. The diversity of plant heat stress transcription factors[J].Trends in Plant Science, 2007, 12(10):452-457.
[8] DORING P. Arabidopsis and the heat stress transcription factor world: How many heat stress transcription factors do we need?[J]. Cell Stress & Chaperones, 2001, 6(3):177-189.
[9] SCHARF K D, BERBERICH T, EBERSBERGER I, et al. The plant heat stress transcription factor (Hsf) family:Structure, function and evolution.[J]. Biochimica et Biophysica Acta, 2012, 1819(2):104-119.
[10] GUO M, LIU J H, MA X, et al. The plant Heat Stress Transcription factors (HSFs): Structure, regulation, and function in response to abiotic stresses[J]. Frontiers in Plant Science, 2016, 7(273):114.
[11] YOSHIDA T, SAKUMA Y, TODAKA D, et al. Functional analysis of an Arabidopsis, heat-shock transcription factor HsfA3, in the transcriptional cascade downstream of the DREB2A stress-regulatory system[J]. Biochemical & Biophysical Research Communications, 2008, 368(3):515-521.
[12] DEBBIE WINTER, BEN VINEGAR, HARDEEP NAHAL, et al. An "Electronic Fluorescent Pictograph" browser for exploring and analyzing Large-Scale biological data sets[J]. PloS one, 2007, 2(8):e718.
[13] HWANG S M, KIM D W, WOO M S, et al. Functional characterization of Arabidopsis HsfA6a as a heat-shock transcription factor under high salinity and dehydration conditions[J]. Plant Cell & Environment, 2014, 37(5):1202-1222.
[14] MISHRA S K, TRIPP J, WINKELHAUS S, et al. In the complex family of heat stress transcription factors, HsfA1 has a unique role as master regulator of thermotolerance in tomato[J]. Genes & Development, 2002, 16(12):1555-1567.
[15] NISHIZAWAYOKOI A, NOSAKA R, HAYASHI H, et al. HsfA1d and HsfA1e involved in the transcriptional regulation of HsfA2 function as key regulators for the Hsf signaling network in response to environmental stress[J]. Plant & Cell Physiology, 2011, 52(5):933-945.
[16] DAISUKE OGAWA, KAZUO YAMAGUCHI, TAKUMI NISHIUCHI. High-level overexpression of the Arabidopsis HsfA2 gene confers not only increased themotolerance but also salt/osmotic stress tolerance and enhanced callus growth[J]. Journal of Experimental Botany, 2007, 58(12):33-73.
[17] QIN C, YU C, SHEN Y, et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization.[J]. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(14):5135-5140.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
