江淮地區花馬鹿遺傳多態性分析
2018-01-30 02:01:28冀德君張威樊永亮魏定國黃必忠
畜牧獸醫科學 2018年1期
冀德君,張威,樊永亮,魏定國,黃必忠
(1.揚州大學動科學院, 江蘇 揚州 225009;2.江蘇揚州市威威鹿場, 江蘇 揚州 225000;3.邗江區科技局, 江蘇 揚州225000)
梅花鹿(Cervus nippon)是一種具很高經濟價值的食草動物[1],其鹿茸的藥用價值首屈一指[2,3]。我國東北地區有圈養梅花鹿的傳統[4]。我國的人工養殖梅花鹿屬東北亞種[5],茸產量中等[6]。北方養殖的梅花鹿遷移到南方地區養殖,適應性下降,生產性能也下降,并且容易出現猝死的問題[1]。不過,梅花鹿始終具有在我國所有氣候條件下生存的潛能,可能由于長期的馴化過程,僅能適應北方氣候環境條件。我國歷史上在中、東部地區的長期農業開發,使得梅花鹿的棲息地受到破壞,導致我國野生梅花鹿瀕臨滅絕,局限分布于東北大興安嶺等地區[5]。揭示梅花鹿及其近緣物種的遺傳特性及對特定區域、氣候的適應性狀況將為茸鹿的廣泛推廣提供寶貴的基礎依據。
早在20 世紀 70 年代,我國開展了鹿種間和亞種間雜交生產研究[7-10],花-馬雜交可能會大大提高鹿的生產性能。雜交茸鹿適應性強、抗病力強、耐粗飼、生長快。不過雜交實踐中仍會出現失敗的情況。為揭示花馬雜交遺傳學的特性變化,本研究依托位于揚州地區某鹿場,采用東北雙陽梅花鹿和新疆天山馬鹿雜交,采用簡化基因組測序的方法,同時利用生物信息學工具,分析了具優良適應性特點的花-馬雜交后代群體的遺傳多樣性,為花馬雜交的推廣提供有用的信息[11,12]。
1 材料與方法
1.1 動物群體
試驗動物選自我國東部江淮地區一個花馬雜交鹿養殖場(揚州市威威鹿業有限公司)。將健康成年種公馬鹿與母鹿(梅花鹿或花馬雜交鹿)混養。按1∶10的比例配置種公鹿和母鹿,混群飼養,45d左右觀測母鹿的受孕情況。幼鹿斷奶后另組群,跟蹤測定其生產和適應性能。
1.2 簡化基因組測序分析
1.2.1 樣本
采用無菌方法分別自5個雜交公鹿的鹿茸上取1~2g帶血鹿茸組織樣,冷凍保存,提取總DNA并進行簡化基因組測序。
1.2.2 步驟
將樣本DNA采用限制性內切酶進行酶切,RAD建庫,即構建300~500bp插入片段的文庫;Illumina HiSeq2000測序。
1.2.3 生物信息學分析
將樣品RAD-tag比對歸類,按每類tag的深度信息進行由大到小排序,獲得樣本的RAD-tag頻數。 接著對樣品RAD-tag兩兩比對,獲得樣品的雜合位點信息。 進一步分析樣品間的RAD-tag差異信息。 通過綜合每個個體RAD-tag的頻數和比對差異信息,濾掉重復區域的數據,最終得到可信度高的群體SNP分型結果。
1.3 遺傳多樣性分析與BLAST功能分析
隨機選取20條含SNP的序列片段,使用DNAStar、DNAsp等軟件對這些片段進行多態性分析,運用聚類分析軟件繪制系統發生樹。通過采用blast方法與多種數據庫序列比對,獲得這些序列片段的相關功能注解信息。
2 結果與分析
2.1 花-馬雜交后代的生產與適應性能
花-馬雜交后代整體表現較好,成活率高,體格健康,少見疾病的發生。繁育率較高,總適應能力評價為總體適應性良好。
2.2 花-馬雜交后代的遺傳多樣性
2.2.1 簡化基因組分析的總體評估
將每個個體之間的RAD-tag(80bp)進行內部比對和兩兩之間的比對,整合深度信息和比對結果信息,從而得到高可信度的群體SNP分型結果,最后總共得到489,288個群體SNP標記。
2.2.2 基因座位的遺傳多樣性分析
整合深度信息和比對結果信息,得到高可信度的群體SNP分型結果,最后總共得到489,288個群體SNP標記,說明該群體的種群內的遺傳多樣性非常豐富。對所有SNP位點進行比較分析,發現變異最高的3個基因片段(80 bp),均包含有2個或3個SNP位點,定義了5個等位基因 (圖1)。
用MEGA軟件確定變異位點,以及運用MEGA軟件計算各群間的遺傳距離,并進行構建進化樹。
基于遺傳距離進行親緣關系聚類分析表明,5頭公鹿分為2個類群,表明其具有至少2個血統來源(圖2);
運用BLSAT軟件進行功能性分析,發現這些多態性較豐富的序列區段相對集中分布于染色體上有關適應功能的分布區域。
花馬雜交鹿在基因水平上存在的海量SNP位點數以及較多的高變區段,說明江淮地區的花馬雜交后代群體具備培育不同特性品系的遺傳潛能,為進一步的品系培育提供遺傳上的素材;同時,編碼區上的高多態性初步提供了鹿產品可能存在特殊活性成分的分子生物學證據。
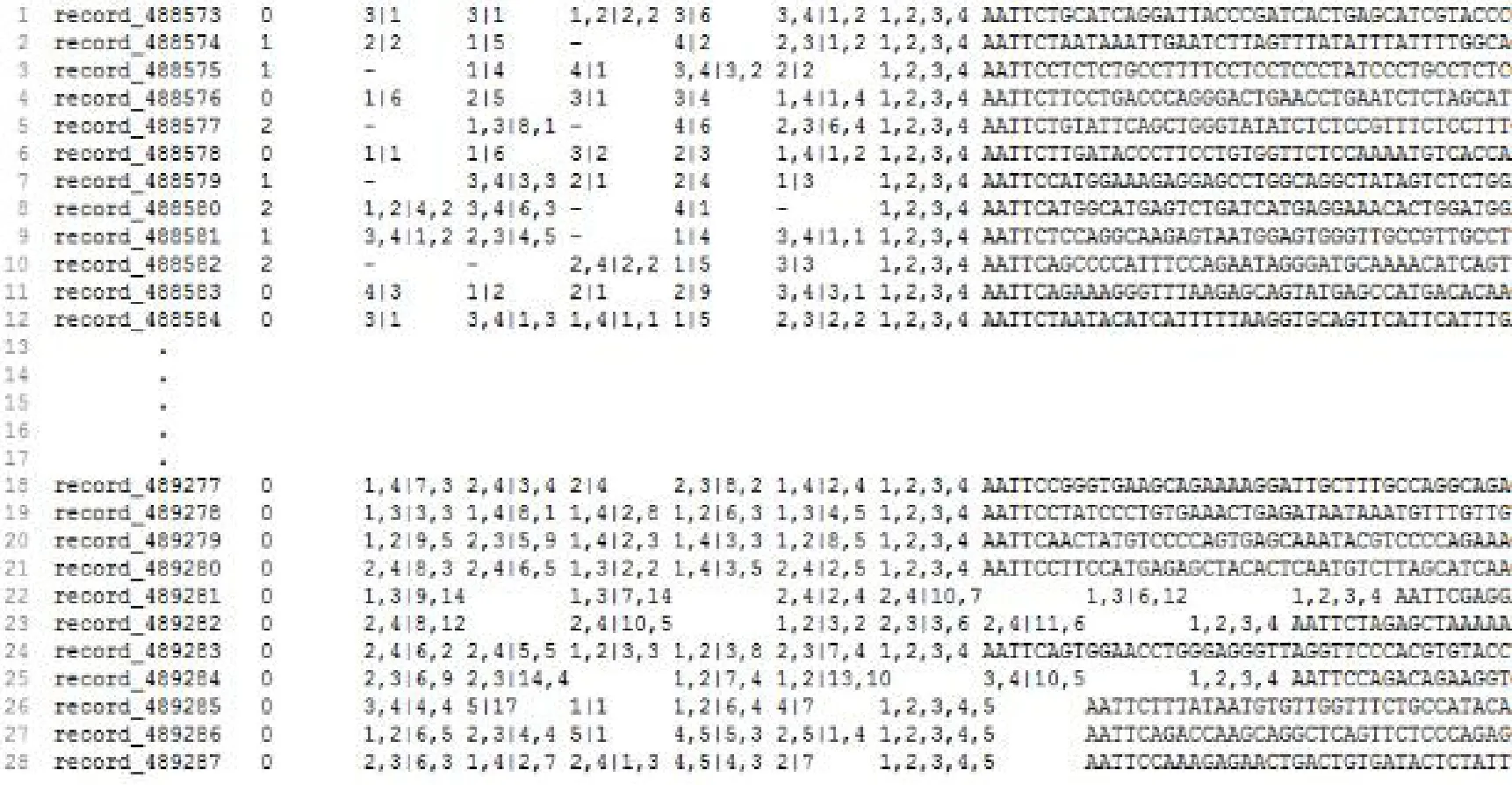
圖1 含四個基因型的80bp序列區段
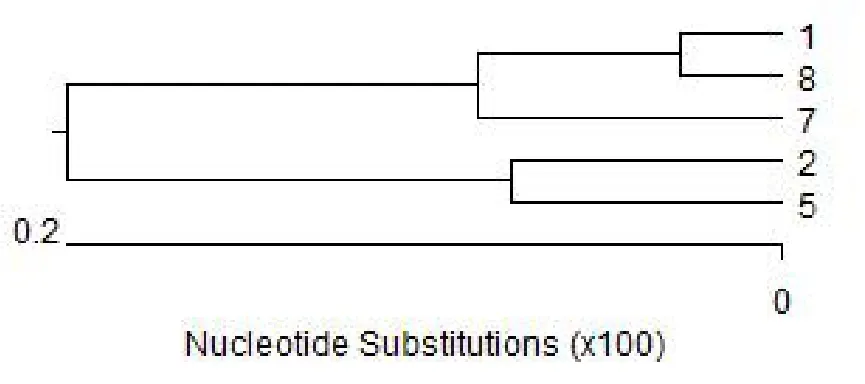
圖2 5頭雜交鹿的系統發生樹
3 討論
3.1 雜交后代有優勢
雜交優勢可大幅度提高動物生產效益,在家畜中被普遍認可。雜交茸鹿具有適應性強、抗病力強、耐粗飼、生長快的特點,我國自70年代初即開始推廣生產應用。
在鹿茸生產上,東北梅花鹿的茸型好、質優,具有品質和價格優勢,可惜茸產量不高,遷移南方地區養殖往往易發病,適應力不夠,且生長速度慢。而新疆的天山馬鹿具有較高的鹿茸產量,適應力好,生長速度快,不過其鹿茸價格低。可以考慮利用雜交的方法,將二者優點相結合,也可能會較快地提高梅花鹿的鹿茸生產水平。
3.2 茸鹿群體的遺傳多樣性
梅花鹿與馬鹿親緣關系較近, 兩者雖然是不同的物種,但在自然條件下兩者可雜交,并且可繁殖后代。據文獻報道,馬鹿是梅花鹿由中東向歐洲和北非擴展過程中產生的。原始型的馬鹿在新世中期從歐洲北上然后返轉,通過絲綢之路的天山北路返回中國。天山馬鹿的遺傳特性對鹿的選育是不可或缺的。另一方面, 梅花鹿可分為2類:亞洲大陸類群,從越南梅花鹿到臺灣梅花鹿, 最后是東北梅花鹿,和日本類群,其很可能由東北梅花鹿直接演化為日本梅花鹿, 同時也向馬鹿過渡。馬鹿類群包括甘肅馬鹿和阿拉善馬鹿,而天山馬鹿可能是亞洲類群向歐洲類群過渡的亞種。眾所周知,遺傳多樣性的喪失會對物種生存可能帶來不利影響。據推測,保持鹿種內遺傳多樣性可以減慢由于物種在適應和進化中可能導致其滅絕的過程。
3.3 雜交動物的實踐意義
雖然東北梅花鹿具有茸型好、質優,具有品質和價格優勢,但茸產量偏低,生長慢。考慮我國茸鹿養殖業的困難,雖然東北梅花鹿數量大,但其生產性能很難提高。將天山馬鹿和東北梅花鹿的兩者優點結合起來,很可能會大大提高茸鹿生產水平。利用天山馬鹿作為父本來雜交改良梅花鹿,實踐效果顯著,雜種后代的生產性能,包括產茸性能都有了較大的提高。花馬雜交是當前提高養鹿效益的重要途徑。
本研究進一步展示了鹿科動物具有極豐富的遺傳多樣性。基因組水平的多態性特征反映了鹿科動物具有很好的適應潛能。本研究初步表明花馬雜交具有豐富的遺傳多樣性,可能具有較好的適應性潛能。
[1]郭立梅,李和平.我國茸鹿遺傳資源的利用現狀分析[J].東北林業大學學報,2007,35(6):66-68.
[2]徐巖,許佳明,何璐等.鹿茸多肽對人膠質瘤細胞生長抑制率及細胞周期的影響[J].吉林中醫藥,2014, 10: 1019-1021.
[3]朱文赫,張巍,李妍等.鹿茸多肽對過氧化氫所致血管內皮細胞損傷的保護作用[J].中國藥理學與毒理學雜志,2014,5:697-681.
[4]夢夢,尹峰. 全國梅花鹿養殖狀況調查[J].野生動物雜志,2008, 29(1):47- 49.
[5]郭延蜀,鄭惠珍.中國梅花鹿地史分布-種和亞種的劃分及演化歷史[J].獸類學報,2000,20(3):168-179
[6]熊彩云,高璜,黃曉鳳等.梅花鹿生態學研究進展 [J].江西林業科技,2007(3):44-46.
[7]李和平.中國茸鹿品種(品系)間的雜交效果 [J]. 東北林業大學學報,2002,30(2):87-89.
[8]趙列平,韓歡勝,趙廣華.東北梅花鹿與天山馬鹿種間雜交效果[J].經濟動物學報,2011,3:153-156.
[9]羅劍通,鄭興濤,楊志勇.梅花鹿與馬鹿雜交繁育的技術路線及關鍵技術[J].經濟動物,2012(9):7.
[10]柳存, 秦大偉.天山馬鹿與青海馬鹿茸重性狀的雜種優勢研究 [J].安徽農業科學,2014,42( 26) : 9047,9056.
[11]鄧蓉,鄭文堂,胡寶貴等.試論中國休閑農業發展中的鹿文化挖掘 [J].北京農學院學報,2014(1):13-16.
[12]楊鎰峰,陳秀敏,宋興超等.探索我國養鹿業發展新模式 [J].特產研究,2012(4):65-67.
