結(jié)核分枝桿菌耐丙硫異煙胺相關(guān)基因研究進(jìn)展
2018-02-15 07:44:32譚旎張泓
實用心腦肺血管病雜志 2018年11期
譚旎,張泓
結(jié)核病是結(jié)核分枝桿菌感染引起的一種“古老”傳染病,而目前結(jié)核病仍是全球范圍內(nèi)最常見的傳染病[1]。盡管過去20年間人們?yōu)槎糁平Y(jié)核病流行做出了巨大努力,但耐藥結(jié)核病尤其是耐多藥結(jié)核病(MDR-TB)、廣泛耐藥結(jié)核病(XDR-TB)的出現(xiàn)為全球范圍內(nèi)結(jié)核病防控帶來了巨大挑戰(zhàn)。2018年世界衛(wèi)生組織(WHO)全球結(jié)核病調(diào)查報告顯示,2017年全球范圍內(nèi)新發(fā)結(jié)核病患者數(shù)量約為1 010萬,其中我國新發(fā)結(jié)核病患者數(shù)量約占8.9%;2017年全球估算新發(fā)利福平耐藥結(jié)核病(RR-TB)患者數(shù)量約為55.8萬例,其中82%為MDR-TB,而印度(24%)、中國(13%)和俄羅斯(10%)3個國家新發(fā)MDR-TB/RR-TB患者數(shù)量約占全球新發(fā)MDR-TB/RR-TB患者總數(shù)的47%;耐藥結(jié)核病治療成功率較低,全球平均耐藥結(jié)核病治療成功率約為55%[2]。
《2010年全國第五次結(jié)核病流行病學(xué)抽樣調(diào)查報告》[3]顯示,我國結(jié)核病患者對異煙肼(INH)耐藥率最高,為28.6%,對丙硫異煙胺(Pto)耐藥率為12.9%;肺結(jié)核患者耐藥率前5位藥物依次為INH、鏈霉素(SM)、對氨基水楊酸(PAS)、Pto、氧氟沙星(Ofx),其中初治肺結(jié)核患者耐藥率前5位藥物依次為INH、SM、PAS、Pto、Ofx,復(fù)治肺結(jié)核患者耐藥率前5位藥物依次為INH、利福平(RFP)、SM、PAS、Pto;由于初治肺結(jié)核患者耐藥率前5位藥物中包括3種二線抗結(jié)核藥物,因此我國肺結(jié)核患者耐藥形勢較嚴(yán)峻,需引起重視。
Pto是二線抗結(jié)核藥物之一,口服進(jìn)入人體后會被迅速吸收并廣泛分布于全身組織、體液,且各組織、腦脊液、血液藥物濃度較為接近。近年來,隨著結(jié)核分枝桿菌對Pto耐藥率升高,Pto耐藥機(jī)制研究尤其是耐藥基因研究備受關(guān)注。本文通過檢索國內(nèi)外參考文獻(xiàn),綜述了結(jié)核分枝桿菌耐Pto相關(guān)基因,旨在為結(jié)核分枝桿菌對Pto的耐藥機(jī)制研究提供參考。
1 作用機(jī)制
MDR-TB指同時對INH和RFP耐藥的結(jié)核病,Pto、乙硫異煙胺(Eto)結(jié)構(gòu)與INH類似,均屬INH硫代酰胺類似物,可導(dǎo)致結(jié)核分枝桿菌失去耐酸性及合成霉菌酸的能力,因此治療MDR-TB時Pto與Eto可以互換[4];此外,由于Pto與Eto具有良好的腦脊液滲透作用,因此二者常用于治療藥物敏感性結(jié)核性腦膜炎、粟粒狀結(jié)核病等[4-5]。
Pto是一種前體藥物,在結(jié)核分枝桿菌內(nèi)被單加氧酶ethA活化后與煙酰胺腺嘌呤二核苷酸(NAD)結(jié)合形成Pto-NAD并抑制參與霉菌酸合成途徑的烯酰基載體蛋白(ACP)還原酶、中斷蛋白質(zhì)合成途徑,繼而阻斷霉菌酸生物合成并擾亂結(jié)核分枝桿菌細(xì)胞膜,最終達(dá)到抗結(jié)核分枝桿菌的目的[4]。
研究表明,ethA-ethR基因座編碼單加氧酶,其中ethR屬轉(zhuǎn)錄調(diào)節(jié)因子TetR/CamR家族阻遏物,是一種轉(zhuǎn)錄調(diào)節(jié)因子,ethR的結(jié)合域通過與ethA啟動子區(qū)域結(jié)合而負(fù)調(diào)節(jié)ethA的表達(dá),因此ethA-ethR基因座涉及Pto的活化并調(diào)控Pto的抗酸能力[6-7]。mshA基因編碼的糖基轉(zhuǎn)移酶參與霉菌生物合成及N-乙酰半胱氨酸葡糖胺肌醇(MSH)合成,而MSH可通過ethA促進(jìn)Pto活化并抑制結(jié)核分枝桿菌[4,8]。Ndh基因通過編碼還原型煙酰胺腺嘌呤二核苷酸脫氫酶而調(diào)節(jié)還原型煙酰胺腺嘌呤二核苷酸(NADH)/氧化型煙酰胺腺嘌呤二核苷酸(NAD+)比例,在NAD+與Pto形成Pto-NAD過程中具有重要作用[9-11]。因此,調(diào)節(jié)單加氧酶的ethR基因、mshA基因、ndh基因?qū)Y(jié)核分枝桿菌耐Pto至關(guān)重要。此外,還有研究發(fā)現(xiàn)結(jié)核分枝桿菌中存在Pto生物活化的其他不依賴ethR基因的替代途徑如含黃素腺嘌呤二核苷酸(FAD)的單加氧酶mymA以劑量依賴方式保留結(jié)核分枝桿菌對Pto的敏感性[12]。
2 耐藥機(jī)制
目前,結(jié)核分枝桿菌的耐藥機(jī)制研究主要包括靶基因突變、細(xì)胞壁通透性變化、外排泵機(jī)制、藥物降解及失活酶等,其中靶基因突變是結(jié)核分枝桿菌對Pto耐藥的主要分子作用機(jī)制。由于過去幾十年多數(shù)研究集中于探討Eto而非Pto的耐藥機(jī)制,因此目前報道Pto耐藥相關(guān)的具有硫代酰胺抗性的基因僅有幾個[4],而由于Eto與Pto顯示出高度交叉耐藥性,因此既往關(guān)于Eto的耐藥機(jī)制研究成為分析Pto耐藥機(jī)制研究的主要依據(jù)。目前,多數(shù)學(xué)者認(rèn)為,結(jié)核分枝桿菌對Pto耐藥主要與inhA基因、ethA基因、ethR基因、mshA基因、ndh基因、mymA基因突變有關(guān)[9,12-13],其中ethA基因突變是結(jié)核分枝桿菌對Pto 耐藥的主要基因突變類型[14]。
3 耐藥基因
3.1 inhA基因 既往研究發(fā)現(xiàn),Pto對INH高度耐藥的結(jié)核分枝桿菌有效,但對INH輕度耐藥的結(jié)核分枝桿菌的效果則較差。INH是治療結(jié)核病的有效藥物之一,而Pto是治療MDR-TB的二線抗結(jié)核藥物,INH與Pto均屬前體藥物,需不同的酶進(jìn)行活化,由于存在共同作用機(jī)制而導(dǎo)致二者交叉耐藥。MACHADO等[15]研究表明,inhA基因調(diào)控區(qū)域的突變組合效應(yīng)導(dǎo)致MDR-TB患者結(jié)核分枝桿菌對INH、Pto 均產(chǎn)生耐藥性;近期研究表明,INH耐藥通常是由katG基因、inhA基因、inhA基因啟動子區(qū)域突變導(dǎo)致的[16]。katG基因突變常與INH高度耐藥有關(guān),katG基因編碼的過氧化氫酶可將INH轉(zhuǎn)化為其活性形式,但由于katG基因不參與Pto的活化,因此Pto的作用不受katG基因突變影響,而由于Pto與INH具有相同的最終靶點,因此inhA基因突變可導(dǎo)致inhA基因編碼產(chǎn)物過表達(dá)或修飾而造成INH與Pto交叉耐藥[15,17]。inhA基因突變所致INH耐藥通常為低水平耐藥并可通過加大INH劑量而減輕INH耐藥,因此臨床常經(jīng)驗性使用大劑量INH聯(lián)合Pto治療MDR-TB,而對大劑量INH耐藥且無inhA基因突變患者可選擇Pto代替INH[18-20]。除inhA基因突變外,Pto還與inhA基因啟動子區(qū)域、inhA基因開放閱讀框突變[21](見表1)有關(guān),其可通過降低Pto與NAD的親和力而導(dǎo)致結(jié)核分枝桿菌對Pto耐藥[15]。
3.2 ethA基因與ethR基因 ethA基因也稱etaA基因(Rv3854c),其編碼的單加氧酶ethA是一種含F(xiàn)AD的NADPH特異性酶,能夠進(jìn)行Baeyer-Villiger氧化反應(yīng)[4,22-24]。Pto經(jīng)單加氧酶ethA激活后作用方式與INH非常相似,其活性形式與NAD+反應(yīng)產(chǎn)生Pto-NAD,進(jìn)而抑制inhA基因及霉菌酸生物合成[22-23,25-26]。TAN等[14]研究發(fā)現(xiàn),對Pto耐藥的結(jié)核分枝桿菌ethA基因突變頻率為51.4%(19/37),其中ethA基因第798位AGCAGG突變(絲氨酸266精氨酸)突變頻率為18.9%(7/37),被認(rèn)為是與結(jié)核分枝桿菌對Pto耐藥有關(guān)的最普遍的基因突變類型(見表1)。ethR基因是ethA基因表達(dá)的負(fù)轉(zhuǎn)錄調(diào)節(jié)因子,ethR基因突變導(dǎo)致ethA基因過度表達(dá)時可導(dǎo)致結(jié)核分枝桿菌對Pto耐藥[6]。此外,ethA基因啟動子區(qū)域或其負(fù)轉(zhuǎn)錄調(diào)節(jié)因子ethR基因結(jié)合域內(nèi)出現(xiàn)突變時可通過以下方式導(dǎo)致ethA基因表達(dá)下調(diào):(1)獨(dú)立于ethR基因調(diào)節(jié)而直接降低ethA基因轉(zhuǎn)錄;(2)增加ethR基因轉(zhuǎn)錄而導(dǎo)致ethA基因表達(dá)受抑;(3)影響ethA基因與ethA基因啟動子區(qū)域的結(jié)合[27]。
另有研究表明,ethA基因還可能通過不穩(wěn)定的氧化亞磺酸中間體活化硫代苯甲酰胺、硫代卡巴肽、硫代乙酰胺、硫代乙酮及其他硫代酰胺藥物,這可能是INH與Pto交叉耐藥的重要原因[28-29]。
3.3 mshA基因(Rv0486) 研究表明,對INH和Pto耐藥的結(jié)核分枝桿菌突變體均存在mshA基因突變,mshA基因編碼產(chǎn)物糖基轉(zhuǎn)移酶參與MSH的生物合成[8]。VILCHèZE等[30]研究發(fā)現(xiàn),含有單個堿基對修飾的mshA基因突變體并導(dǎo)致氨基酸改變的8株結(jié)核分枝桿菌MSH含量從99.9%下降至83.0%,而對Pto的耐藥率升高4~8倍;此外,筆者還發(fā)現(xiàn)mshA基因的7個獨(dú)立的錯義及移碼突變,其中4個mshA基因突變體具有單個氨基酸突變,即精氨酸273半胱氨酸、甘氨酸299半胱氨酸、甘氨酸356天冬氨酸、谷氨酸361丙氨酸[8](見表1)。因此,mshA基因缺失、錯義及移碼將導(dǎo)致結(jié)核分枝桿菌產(chǎn)生MSH減少及MSH通過ethA基因促進(jìn)Pto的活化減少,繼而造成結(jié)核分枝桿菌對Pto 耐藥。
ANG等[12]研究發(fā)現(xiàn),單獨(dú)的mshA基因缺失所致Pto對結(jié)核分枝桿菌的半數(shù)最低抑菌濃度(MIC50)較ethA/ethR基因突變高,提示對于Pto殺滅作用的影響,mshA基因突變至少與ethA/ethR基因突變一樣。此外,mshA基因突變可導(dǎo)致敲除ethA/ethR基因的結(jié)核分枝桿菌對Pto完全耐藥,提示Pto殺滅作用中的MSH作用可以是獨(dú)立的,或者至少不限于其與ethA基因的相互作用,可能參與Pto生物激活的替代途徑,但仍需進(jìn)一步研究證實[12]。
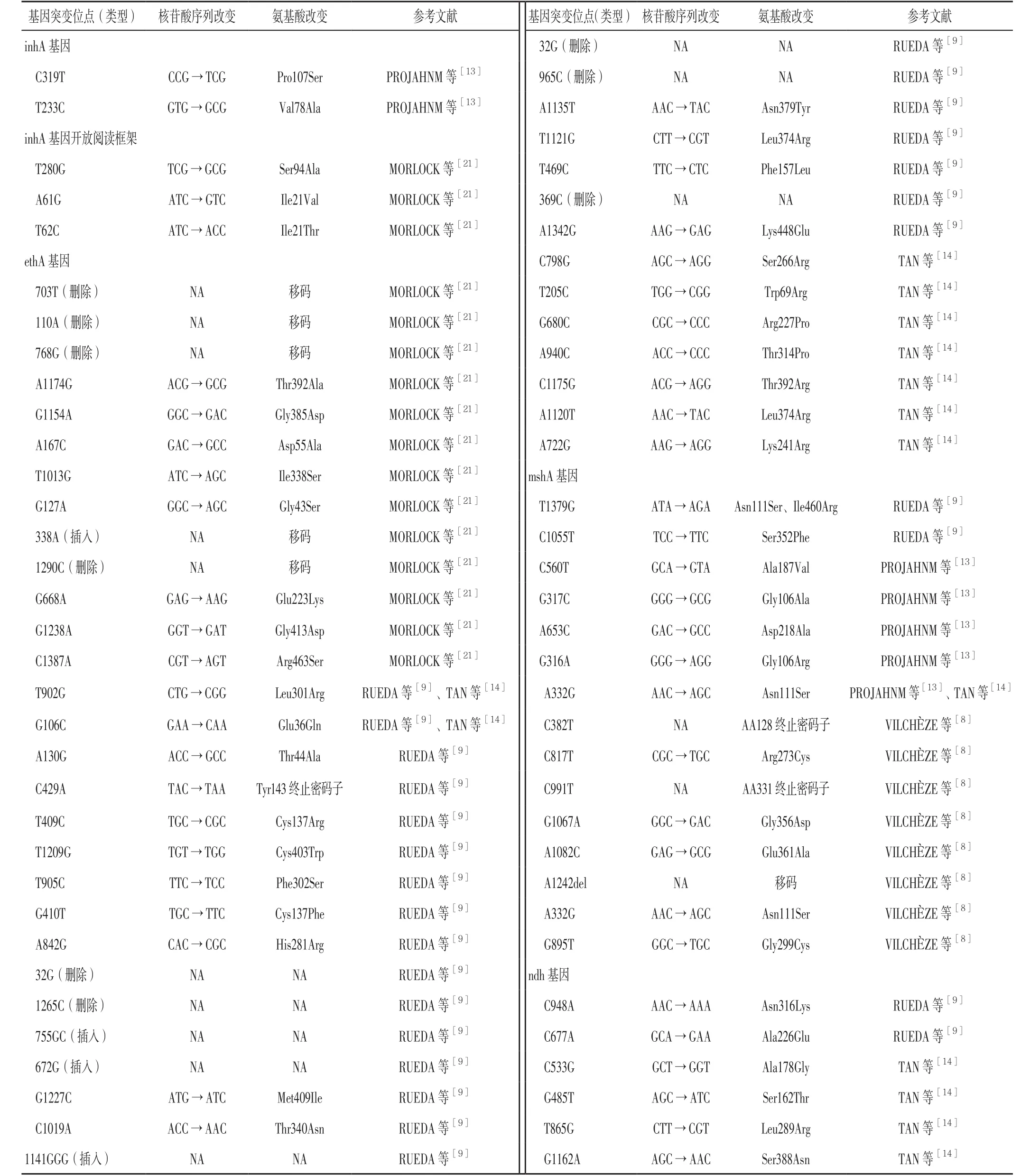
表1 已知的Pto耐藥基因突變位點(類型)、核苷酸序列改變及氨基酸改變Table 1 Known drug resistance related gene mutation site(types)to prothionamide,changes of nucleotide sequences and amino acids
3.4 ndh基因(Rv1854c) ndh基因編碼產(chǎn)物為Ⅱ型還原型煙酰胺腺嘌呤二核苷酸脫氫酶,而Ⅱ型還原型煙酰胺腺嘌呤二核苷酸脫氫酶可通過將NADH氧化成NAD+而調(diào)節(jié)NADH/NAD比值。研究表明,ndh基因突變可導(dǎo)致細(xì)胞內(nèi)NADH含量增加并競爭性地抑制INH-NAD或Pto-NAD及INH或Pto的結(jié)合,繼而導(dǎo)致INH與Pto交叉耐藥[10-11]。TAN等[14]在46株對Pto耐藥的結(jié)核分枝桿菌中發(fā)現(xiàn)37株(80.4%)存在基因突變,基因突變類別包括19種,其中27株為單核苷酸突變,10株為雙核苷酸突變,而27株單核苷酸突變菌株中4株為ndh基因非同義突變,分別為S162T、L289R、G339A、S388N(見表1),證實ndh基因突變可導(dǎo)致結(jié)核分枝桿菌對Pto耐藥。
3.5 mymA基因 既往研究表明,結(jié)核分枝桿菌存在6種Baeyer-Villiger單加氧酶,而在這6種單加氧酶中ethA特征最為明顯,而mymA、ethA則具有最大的序列同源[26,31]。GRANT等[24]研究發(fā)現(xiàn),單獨(dú)的mymA基因突變可導(dǎo)致結(jié)核分枝桿菌對Pto低度耐藥,而mymA基因與ethA基因可通過加成方式活化Pto,這可能是Pto活化及抗性的新分子作用機(jī)制。ANG等[12]研究發(fā)現(xiàn),ethA/ethR基因刪除僅導(dǎo)致結(jié)核分枝桿菌對Pto耐藥性輕微升高,這可能與結(jié)核分枝桿菌存在mymA基因而保留其對Pto的易感性、劑量依賴性藥物反應(yīng)有關(guān)。
4 小結(jié)與展望
耐藥結(jié)核病為全球結(jié)核病防控工作帶來重大挑戰(zhàn),鑒于新型抗結(jié)核藥物的研發(fā)及應(yīng)用所需時間較長,因此提高現(xiàn)有藥物療效是一種不應(yīng)忽視的替代策略。目前,基于Pto耐藥機(jī)制研發(fā)的具有抗結(jié)核分枝桿菌作用的單加氧酶inhA、ethR抑制劑等Pto增效劑研究已取得一定成果,并有望為抗結(jié)核治療方案提供新的選擇[32-34];Pto耐藥機(jī)制及相關(guān)基因突變研究已取得一定進(jìn)展,已發(fā)現(xiàn)多種耐藥遺傳機(jī)制,包括靶基因機(jī)構(gòu)改變、激活前體藥物所需酶功能喪失等,證實基因突變及基因轉(zhuǎn)錄改變是結(jié)核分枝桿菌對Pto耐藥的重要機(jī)制,但由于缺乏系統(tǒng)的全基因組轉(zhuǎn)錄對照,因此部分基因轉(zhuǎn)錄異常導(dǎo)致結(jié)核分枝桿菌對Pto耐藥的機(jī)制仍需進(jìn)一步深入研究。此外,目前已知的結(jié)核分枝桿菌對Pto耐藥相關(guān)突變基因及突變位點仍十分有限,可能還存在尚未發(fā)現(xiàn)的基因突變,相信隨著不斷的探索及新技術(shù)的應(yīng)用,必將逐步發(fā)現(xiàn)可能存在的基因突變及基因轉(zhuǎn)錄改變對結(jié)核分枝桿菌耐藥性的影響,繼而指導(dǎo)臨床制定科學(xué)、有效的抗結(jié)核治療方案。
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
昆明醫(yī)科大學(xué)學(xué)報(2022年1期)2022-02-28 07:43:40
中國生殖健康(2020年2期)2021-01-18 02:51:26
文苑(2018年21期)2018-11-09 01:23:06
小學(xué)生導(dǎo)刊(2018年13期)2018-06-29 03:49:00
中國衛(wèi)生(2015年9期)2015-11-10 03:11:12
中國當(dāng)代醫(yī)藥(2015年17期)2015-03-01 02:03:58
中國衛(wèi)生(2014年3期)2014-11-12 13:18:12
中國火炬(2014年4期)2014-07-24 14:22:19
