不同水分處理下喀斯特土層厚度異質性對兩種草本葉片解剖結構和光合特性的影響
2018-03-08 07:43:42趙雅潔宋海燕陶建平劉錦春
生態學報 2018年2期
關鍵詞:植物
李 周,趙雅潔,宋海燕,張 靜,陶建平,劉錦春
西南大學生命科學學院,三峽庫區生態環境教育部重點實驗室,重慶市三峽庫區植物生態與資源重點實驗室, 重慶 400715
喀斯特是自然環境中一種脆弱的生態環境系統,敏感度高,環境容納量低,災變承受閾值彈性小。緩慢的成土速率造成了喀斯特地區土壤資源缺乏的“先天性”缺陷[1],而近年來人為活動使得水土大量流失,導致“土荒”現象愈演愈烈。土壤資源的嚴重缺乏是造成喀斯特生態系統脆弱的基本原因。在土壤貧瘠的喀斯特地區,底層基石在結構上也極不規則,導致了土層分布在微觀尺度上具有較大波動[2],造成土層分布在空間上的強烈異質性[3]。
橡木桶貯藏后樣品感官得分為 97.68±2.35,4 ℃玻璃瓶密封保存酒樣感官得分為 79.84±1.98。貯藏工藝能夠明顯提高獼猴桃果酒樣品的感官,有助于加速產品成熟。橡木桶貯藏工藝對獼猴桃果酒感官的影響還需進一步進行對照研究。
喀斯特土層分布的空間異質性與其特有的“巖溶干旱”間存在著密不可分的關系[4]。植物的水分供應主要取決于降水和土壤儲水,而土層厚度是土壤水分保蓄的關鍵[5]。朱守謙[6]發現,在貴陽花溪石灰巖山丘上,雨后連續晴4d后,土層較厚的石洞土壤含水量為67.7%,而土層較淺的土面土壤含水量僅為31.3%。可見,喀斯特土層分布的空間異質性導致該地區的水分分布也具有高度空間異質性[7]。隨著全球氣候變化,降雨格局變化的問題逐漸凸顯,我國喀斯特所在的亞熱帶地區的降雨量出現下降趨勢[8-10]。在氣候變化過程中,土層厚度對土壤水分和養分在短期內的有效性具有重要的緩沖作用。有研究認為,在一些淺土區域,由于自身水分涵養能力低,其土壤水分狀況直接受到降雨變化的影響,而在一些深土區域,較為充足的土壤資源有利于其土壤水分狀況在降雨和蒸發過程中得到有效緩沖,在氣候變化過程中具有相對穩定的微環境[11]。
在植物生長發育的早期,土壤水分狀況直接影響到植物的形態建成[12],并且這種影響是不可逆的。對于大部分植物而言,在中度干旱脅迫下,氣孔密度增加[13-14],而在重度干旱脅迫下,氣孔密度減少[15]。Zhao等[16]認為水分虧缺刺激了氣孔生成,導致氣孔密度增加,但氣孔大小和孔徑卻降低,但Sam等[17]卻覺得干旱脅迫下植物較高的氣孔密度與葉面積減小有關。此外,在干旱環境中,植物通常由于水分脅迫而引起木質部栓塞,導致木質部導水率下降,較高的葉脈密度被認為是一種有效地抗旱策略[18],因為葉脈密度越高,植物的水分運輸途徑越多,有利于避開栓塞化的木質部運輸水分到蒸騰部位的細胞。
葉脈和氣孔性狀與植物的光合能力間存在著一定的協同變異關系[19]。較高的葉脈密度和較低的葉脈間距被認為有利于支持較高的凈光合速率[20-21],植物葉片氣孔密度越大,有利于短時間內充分利用有效水分進行光合作用[22]。因此,植物的氣孔、葉脈性狀和光合能力對土壤水分狀況都有著較為直接的響應表現。另一方面,植物的光合作用還與土壤資源密切相關。土壤資源匱乏造成植物生長所需的養分不足,影響植物的水分運輸效率[23],同時也進一步加重干旱脅迫對植物的影響,導致植物光合作用降低。
喬木和灌木具有發達的木質根系,能夠深入到深層巖石裂隙中吸取水分和養分,而草本植物通常具有較淺的肉質根系,主要利用地表有土壤覆蓋的斑塊生長[7],所以在喀斯特地區,草本植物的生長更容易受到土層厚度的影響。黑麥草(LoliumperenneL.)和葦狀羊茅(FestucaarundinaceaSchreb.)均為禾本科多年生草本植物,須根發達,但二者具有不同的根深,黑麥草的根深大致在25 cm左右,而葦狀羊茅的最大根深能達到40 cm[24-25]。此外,黑麥草和葦狀羊茅都具有很強的適應能力,在喀斯特地區的引種實驗中綜合性狀排名靠前,廣泛應用于喀斯特生態恢復工作中[26-27]。
三因素方差分析結果表明,兩種植物的胞間CO2濃度、蒸騰速率和氣孔限制值均存在顯著差異,物種、水分和土層厚度三因素的交互作用對植物的胞間CO2濃度、蒸騰速率和氣孔限制值未產生顯著影響,但水分和土層厚度兩因素的交互作用對以上3個光合參數產生了顯著影響(表4)。
1 材料與方法
1.1 試驗材料
本試驗選擇的供試材料為多年生黑麥草(L.perenne)和葦狀羊茅(F.arundinacea),供試土壤是來自重慶市中梁山的黃色石灰土,基本理化性狀:pH為7.4±0.14,有機質為0.34%±0.02%,全氮為(0.28±0.03) g/kg,全磷為(0.39±0.02) g/kg,全鉀為(23.7±3.22) g/kg。
1.2 研究方法
試驗設置了3個土層厚度水平,通過3種底面積相同但高度不同的長方體栽培容器實現,容器底部設有5個小孔,以便多余水分流走。本試驗所采用的黑麥草和葦狀羊茅兩種植物均為須根系植物,根系主要分布在15 cm以內的表土層[28],因此設置15 cm厚的土層為對照組(SCK),設置5 cm和30 cm分別為淺土組(SS)和深土組(SD), 3種容器的底面積均為0.01 m2,3個組依次裝入500 g、1500 g和3000 g重的干土。2015年1月14日,在西南大學生態園內進行播種,于2015年4月4日,選取長勢一致的黑麥草和葦狀羊茅幼苗移植于容器內,每盆2株。將所有盆栽試驗用苗置于西南大學生態實驗園地中(海拔高度249 m)進行相同光照管理適應。
相對本科的通識教育,大部分高職院校的通識教育并沒有得到普遍重視,無論是理論建設還是實踐落實都存在不少問題。
此外,每種水分處理和土層厚度均單獨設置3個無植物生長的空白處理,進行和上述試驗同步的水分處理,用于土壤含水量的測定。土樣采集是在每次水分處理之前,通過五點取樣法在容器的4個角和中間位置的不同土層厚度采集土壤樣本混合均勻,放入鋁盒中,帶回實驗室用稱重法進行土壤含水量測定,并根據公式:土壤貯水量=土壤絕對含水量×土壤容重×土壤體積,計算不同水分處理下不同厚度土層的貯水量。
1.3 指標測量
本研究的數據處理與統計分析均采用SPSS 17.0和Microsoft Office Excel 2007軟件完成。采用Three-way ANOVA分析物種、水分和土層厚度3種因素對植物的各葉片解剖結構特征和光合參數的影響;利用One-way ANOVA分析相同水分條件下同一物種各葉片解剖結構特征和光合指標在不同土層厚度間的差異,P<0.05表示差異顯著。利用Origin 8.6軟件作圖。
(1)氣孔密度和大小:采用印跡法獲取氣孔[29],在葉片近軸面和遠軸面的中央位置(避開主脈)均勻地涂上無色透明指甲油,待其干透后撕下,制成臨時裝片。于光學顯微(Nikon edipse 80i)下放大40倍后拍照,然后在Photoshop CS4軟件中統計每張照片中的氣孔數目,并測量氣孔面積(氣孔器面積,包括保衛細胞和副衛細胞)。每個處理選擇5株植物,每株植物在相同部位取成熟健康葉1片制作裝片,每張裝片隨機選擇3個視野拍照,對每張照片中的氣孔數目進行統計,并隨機選擇3個氣孔進行氣孔大小測量。氣孔密度(Stomatal density)用單位面積(mm2)的氣孔數量表示[30];
(2)葉脈密度:用5%的NaOH溶液浸泡葉片4—5 d,葉片會逐漸褪色直至透明(每天更換浸泡液)。再用去離子水沖洗,經番紅-固綠染色后制成臨時裝片,在光學顯微鏡下放大10倍后拍照。在Photoshop CS4軟件中測量每張照片中的葉脈總長度。每個處理選取5片葉子,每片葉子拍攝3個視野。葉脈密度(Vein density)用單位面積(mm2)的葉脈總長度(mm)表示[31];
(3)光合參數:每個處理選擇3株植株,每株選取相同部位的成熟健康葉1片,使用便攜式光合儀LI- 6400(Li-Cor 6400,Li-Cor Inc,USA),于上午9:30—12:00進行光合測定。測定參數包括:凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)、蒸騰速率(Tr),此外,氣孔限制值(Ls)根據公式(Ls=1-Ci/Ca)計算得到[32]。公式中的Ca為大氣中CO2濃度。研究表明,黑麥草和葦狀羊茅的光飽和點大約在800—1200 μmol m-2s-1的范圍內[33- 35],在固定光照為1000 μmol m-2s-1時,兩種植物均表現較強的光合能力,因此本試驗光合測定的光照強度設定為1000 μmol m-2s-1。
1.4 數據處理
水分處理69 d(22次)后,進行以下指標測定:
2 結果
2.1 土壤水分狀況
在正常水分處理下(Wck),淺土組和對照組的土壤含水量分別為21.3%和20.28%,均屬于偏濕生境,深土組的土壤含水量(17.03%)在適宜范圍內[36],淺土組和對照組的土壤含水量無顯著差異,但二者顯著高于深土組(表1);在D1水分處理中,3種厚度土層的土壤含水量在12.82%—14.46%之間,均為輕旱生境,但淺土組的土壤含水量顯著高于對照和深土組;在D2水分下,3種厚度土層的土壤含水量在7.29%—9.05%之間,均屬于中旱水平,深土組和對照組的土壤含水量顯著高于淺土組。此外,在3種水分處理下,淺土組的土壤貯水量均顯著低于對照組,而深土組均顯著高于對照組。兩因素方差分析結果表明,土壤含水量和貯水量受水分和土層厚度兩種因素交互作用的影響而存在顯著差異(表2)。
狗娃是小珍的哥哥。他的兩只眼睛賊亮,繞著一棵棵樹伸長了脖子,不一會兒便找到了一棵野山楂。瘦長的身子猴兒一般縱身一躍就抓住結滿簇簇果實的枝,一邊摘,一邊笑我們笨。
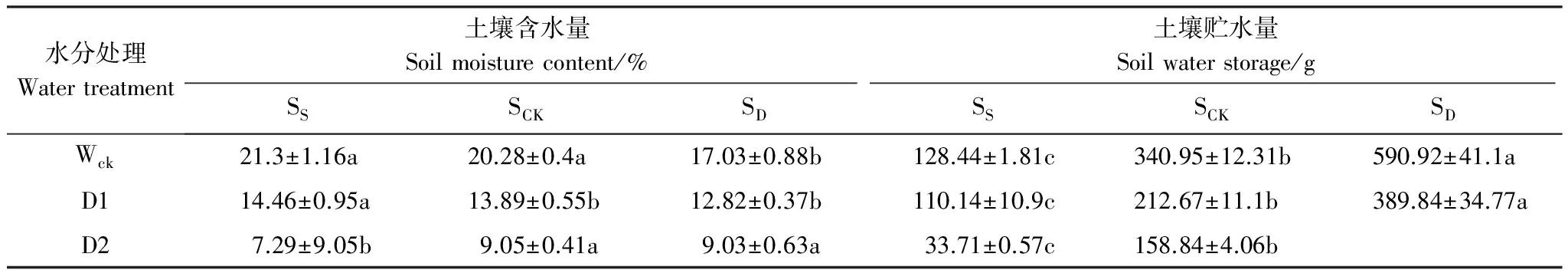
表1 不同水分條件下不同厚度土層的土壤水分狀況(均值±標準誤)
SS:淺土組,shallow soil;SCK:土層厚度對照組,soil control;SD:深土組,deep soil;Wck:正常供水組,water control;D1:減水1組, water reduction group 1;D2:減水2組,water reduction group 2;不同小寫字母表示相同水分處理下不同土層厚度間差異顯著(P<0.05)
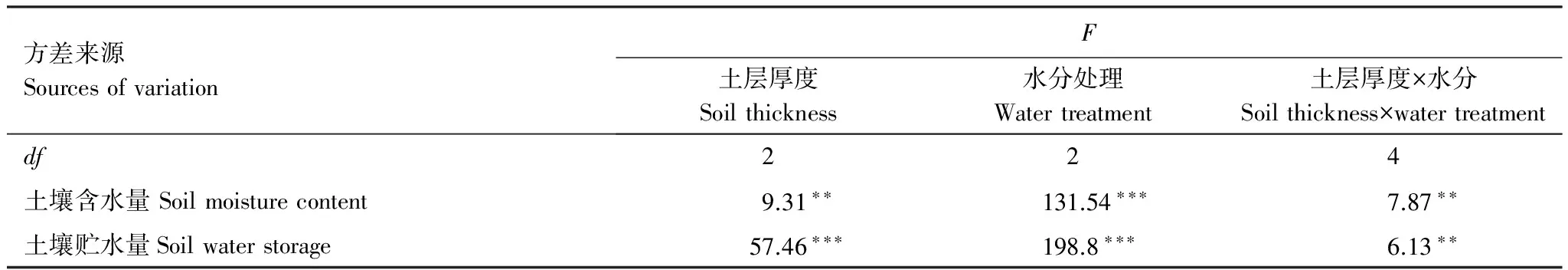
表2 土壤水分狀況的兩因素方差分析
ns未達到顯著水平(P>0.05),*,**和***分別代表P<0.05,P<0.01和P<0.001
2.2 葉片解剖結構特征
三因素方差分析結果表明,兩種植物的近軸面和遠軸面氣孔密度之間均存在顯著差異,且植物的近軸面和遠軸面氣孔密度受物種、水分和土層厚度三因素交互作用的影響存在顯著差異(表3)。
在淺土組,黑麥草近軸面氣孔密度在正常水分條件下(Wck)較對照組顯著增加34.81%,在D1水分下也顯著高于對照組,增加幅度為41.49%,但在D2水分條件下,卻較對照組顯著降低45.79%。在深土組,黑麥草近軸面氣孔密度在3種水分條件下均與對照組無顯著差異。黑麥草在淺土組的遠軸面氣孔密度在Wck、D1和D2這3種水分條件下均與對照組無顯著差異,而黑麥草在深土組的遠軸面氣孔密度在Wck和D1水分下與對照組無顯著差異,但在D2水分條件下,較對照組顯著降低32.56%(圖1)。
在淺土組,葦狀羊茅近軸面氣孔密度在正常水分條件下(Wck)較對照組顯著增加22.89%,在D1水分下顯著降低12.74%,在D2水分處理中與對照組無顯著差異。在深土組,葦狀羊茅近軸面氣孔密度在Wck、D1和D2這3種水分處理中均與對照組無顯著差異。葦狀羊茅在淺土組的遠軸面氣孔密度在Wck水分處理中較對照組顯著增加24.65%,在D1水分下與對照組無顯著差異,在D2水分處理中顯著降低14.66%。在深土組中,葦狀羊茅遠軸面氣孔密度在Wck水分下顯著降低12.33%,在D1水分下顯著降低14.47%,在D2水分下與對照組無顯著差異(圖1)。
2.2.1 氣孔密度
2015年4月14日開始進行水分處理。重慶地區1981年至2011年這30年間4—6月每月平均降雨量119.58 mm/m2,按照本試驗具體容器規格計算出每0.01 m2的面積每日降雨量為40 mL(高度為4 mm)。因此,設置正常供水組(Wck)的供水量為40 mL/天,減水1組(D1)和減水2組(D2)在正常供水組水平上分別減少50%(20 mL)和70%(12 mL)。每3天澆一次水(Wck組120 mL/次;D1組60 mL/次;D2組36 mL/次)。
2.2.2 葉脈密度
1981年12月,中國常駐聯合國代表凌青在聯合國第五委員會上發言表示中國政府準備對今后聯合國維和行動采取區別對待的靈活立場。1982年,中國開始承擔聯合國維和費用。這個時期中國對聯合國的態度可從當時英文版的《北京周報》窺見一斑,“國際局勢越動蕩,世界和平和國際安全所面臨的問題就越嚴重,加強聯合國作用的任務就越重要、越迫切”。①1984年,中國全面闡述了對維和行動的原則立場,積極支持聯合國在國際社會發揮作用。
在淺土組,黑麥草的葉脈密度在Wck和D1水分條件下與對照組無顯著差異,在D2水分下較對照組顯著增加30%。在深土組,黑麥草葉脈密度在Wck水分條件下較對照組顯著降低7.14%,在D1水分處理下顯著增加12.6%,在D2水分下與對照組無顯著差異(圖2)。
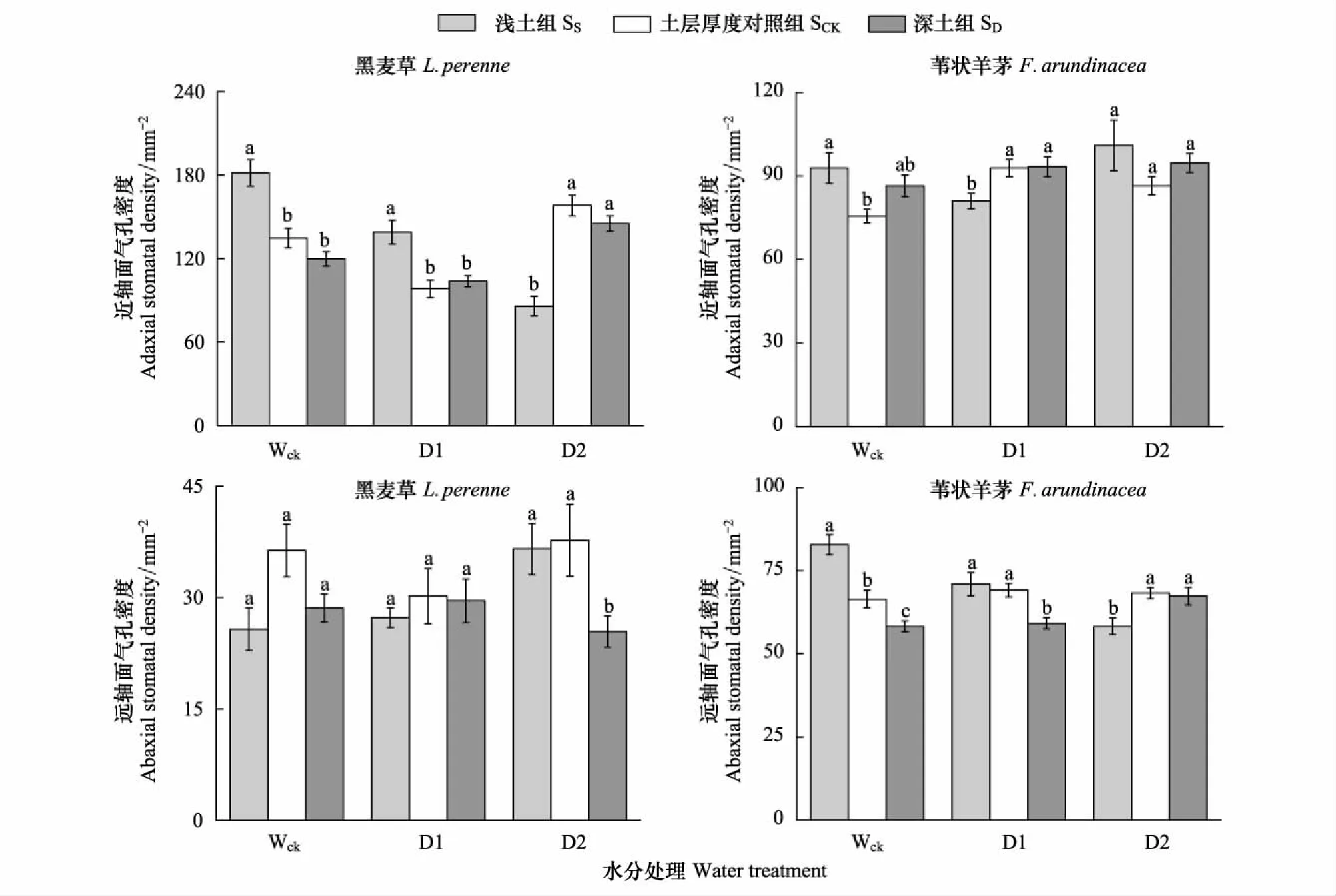
圖1 不同水分處理、不同土層厚度下黑麥草和葦狀羊茅的近軸面和遠軸面氣孔密度(均值±標準誤)Fig.1 The adaxial and abaxial stomatal density of L. perenne and F. arundinacea under different soil thickness and water treatments (M±SD)SS:淺土組,shallow soil;SCK:土層厚度對照組,soil control;SD:深土組,deep soil;Wck:正常供水組,water control;D1:減水1組, water reduction group 1;D2:減水2組,water reduction group 2;柱上的小寫字母表示相同水分處理下不同土層厚度間差異顯著(P<0.05)
Table3ResultsofThree-wayANOVAtestfortheeffectsofspecies,soilthicknessandwatertreatmentonleafanatomicalstructureparameters
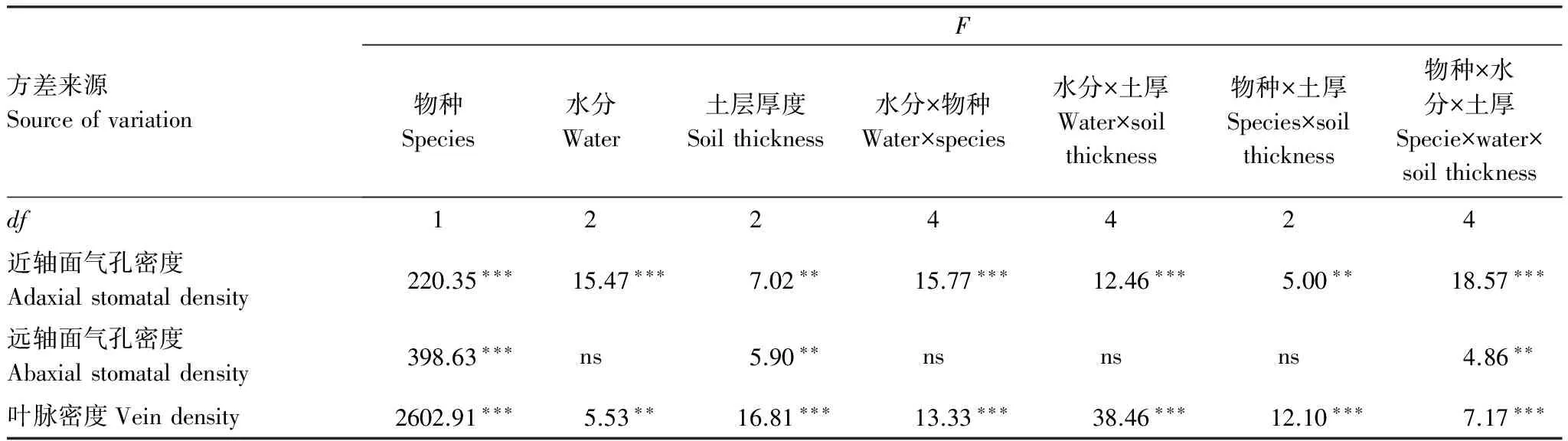
方差來源SourceofvariationF物種Species水分Water土層厚度Soilthickness水分×物種Water×species水分×土厚Water×soilthickness物種×土厚Species×soilthickness物種×水分×土厚Specie×water×soilthicknessdf1224424近軸面氣孔密度Adaxialstomataldensity220.35???15.47???7.02??15.77???12.46???5.00??18.57???遠軸面氣孔密度Abaxialstomataldensity398.63???ns5.90??nsnsns4.86??葉脈密度Veindensity2602.91???5.53??16.81???13.33???38.46???12.10???7.17???
在淺土組,葦狀羊茅葉脈密度在Wck和D1水分處理中均與對照組無顯著差異,在D2水分條件下較對照組顯著增加16.9%。在深土組,葦狀羊茅葉脈密度在Wck水分條件下較對照組顯著增加9.9%,在D1水分處理中顯著增加20.8%,在D2水分條件下與對照組無顯著差異(圖2)。
然而,北方的人們卻很難體會這樣的年味兒。北方的冬天氣溫低,寒冷的室外就是天然冰柜,無須腌漬臘味來保存食物,因此也就沒有冬天吃臘味的習慣。而南方冬天溫度較高,宰殺豬或雞鴨之后,如果不盡快吃掉,肉很快就會變質。于是,南方人便將肉類用鹽和其他香料一同浸漬,再將鹽漬的肉曬干或陰干。
三因素方差分析結果表明,兩種植物的葉脈密度間存在顯著差異,并且物種、水分和土層厚度三因素的交互作用對植物葉脈密度產生了顯著影響(表3)。
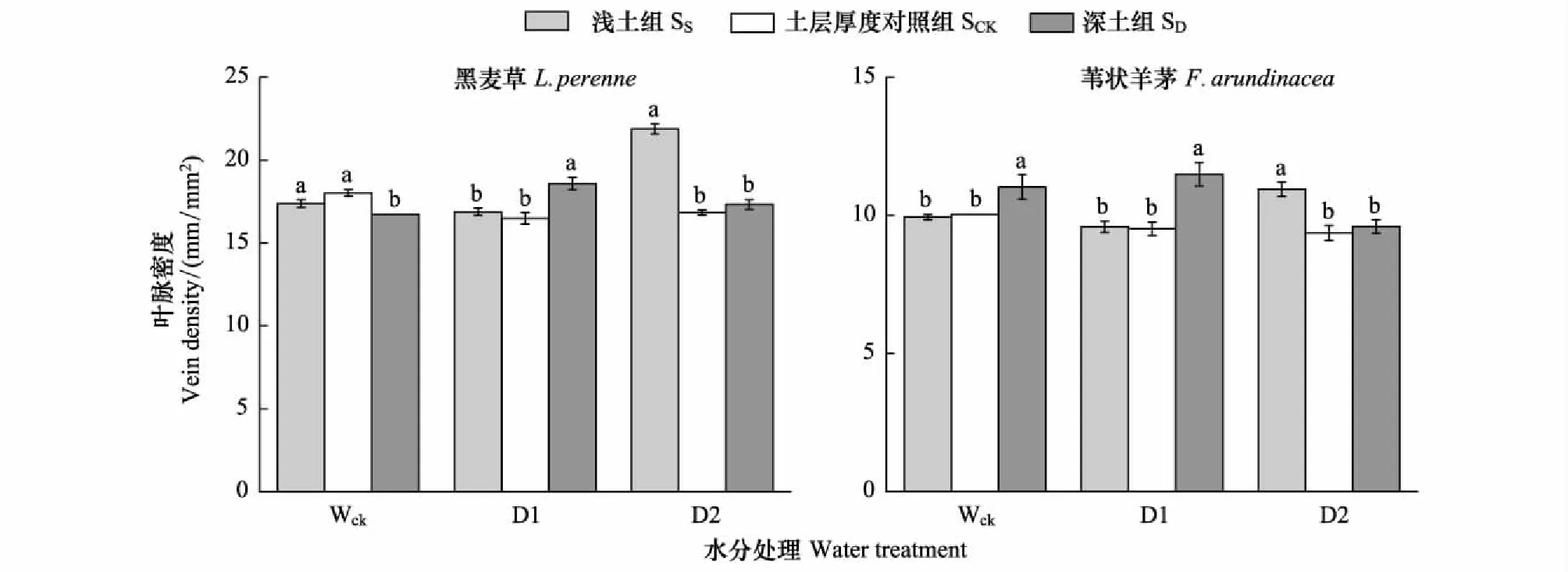
圖2 不同水分處理、不同土層厚度下黑麥草和葦狀羊茅的葉脈密度(均值±標準誤)Fig.2 The vein density of L. perenne and F. arundinacea under different soil thickness and water treatments (M±SD)
2.3 光合特性
2.3.1 凈光合速率Pn
黑麥草在淺土組的凈光合速率Pn在3種水分條件下均低于對照組,在正常水分條件下(Wck)顯著降低27.3%,在D1水分下顯著降低30.7%,在D2水分下,黑麥草在淺土組的Pn較對照組降低17.7%,但二者間的差異不顯著。在深土組,黑麥草的Pn在Wck水分下較對照組顯著降低22.1%,在D1水分下顯著降低30.6%,在D2水分下,黑麥草在深土組的Pn較對照組增加了27.9%,二者間差異不顯著,但淺土組和深土組間差異顯著(圖3)。
在淺土組,葦狀羊茅的凈光合速率Pn在Wck水分條件下較對照組降低了26.4%,但二者間差異不顯著,在D1水分下與對照組無顯著差異,在D2水分下顯著下降51.9%。在深土組,葦狀羊茅的Pn在Wck水分條件下增加了58.5%,但與對照組的差異不顯著,在D1水分處理中顯著增加了127.9%,在D2水分條件下與對照組無顯著差異(圖3)。
三因素方差分析結果表明,兩種植物的凈光合速率存在顯著差異,并且物種、水分和土層厚度三因素的交互作用對植物凈光合速率產生了顯著影響(表4)。
水生態文明建設是我國當前一項長期而又艱巨的歷史任務[7],應堅持規劃先行。而城市人口資源更密集,資源環境矛盾更突出,對美好生活的需求更迫切,因此更加需要強化頂層設計對城市水生態文明建設的指導作用。建議在摸清家底、理清思路的情況下,做好水生態文明建設頂層設計,統籌水、路、岸、產、城,統領水利、生態環境、農業、林業、城建、國土、產業等各類規劃,繪制一張藍圖,解決現有各類規劃自成體系、內容交叉重疊、空間管控分區相互沖突等問題。針對目前部分城市河湖生態空間受擠壓的現象,應在頂層設計中突出河湖生態空間保護的重要性,守住城市水生態空間,實現城市功能與水域空間的有機融合與滲透[8]。
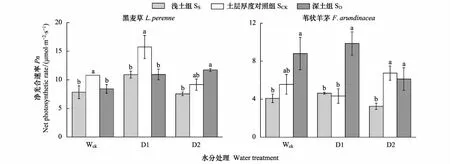
圖3 不同水分處理、不同土層厚度下黑麥草和葦狀羊茅的凈光合速率Pn(均值±標準誤)Fig.3 The net photosynthetic rate(Pn) of L. perenne and F. arundinacea under different soil thickness and water treatments (M±SD)
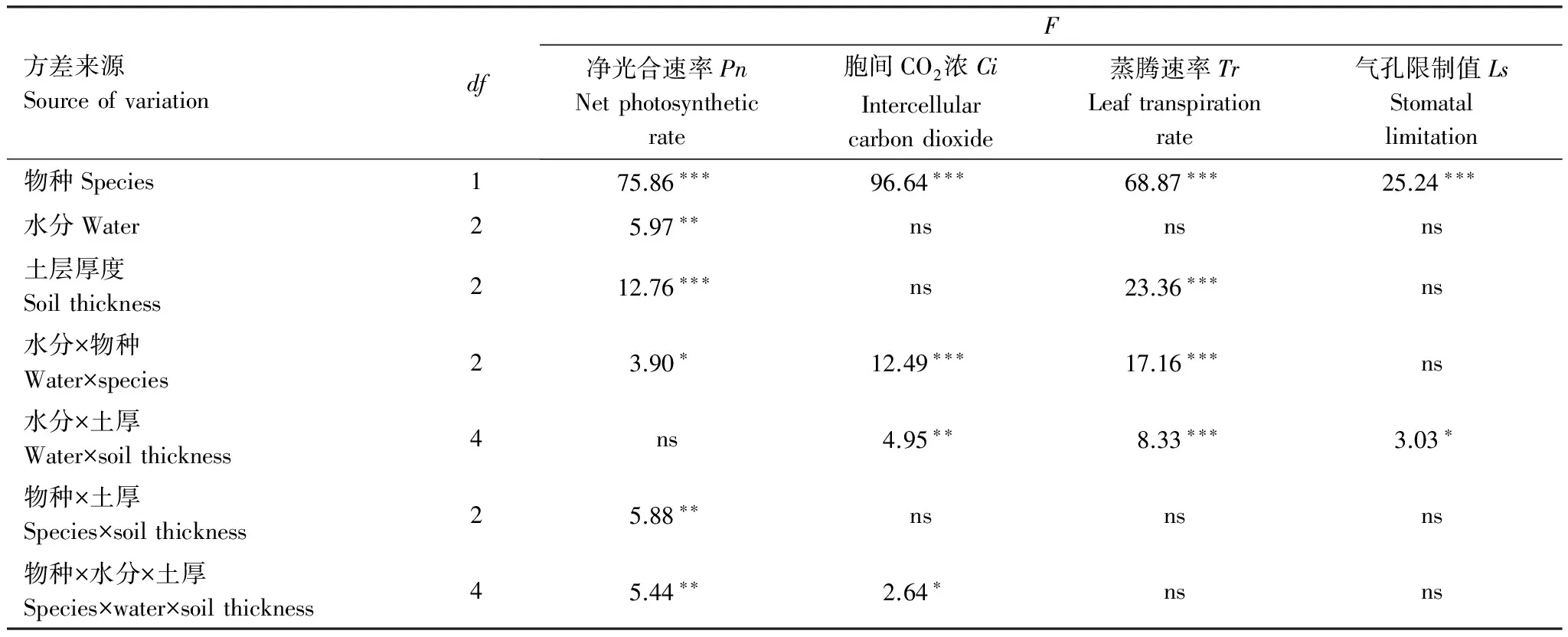
方差來源SourceofvariationdfF凈光合速率PnNetphotosyntheticrate胞間CO2濃CiIntercellularcarbondioxide蒸騰速率TrLeaftranspirationrate氣孔限制值LsStomatallimitation物種Species175.86???96.64???68.87???25.24???水分Water25.97??nsnsns土層厚度Soilthickness212.76???ns23.36???ns水分×物種Water×species23.90?12.49???17.16???ns水分×土厚Water×soilthickness4ns4.95??8.33???3.03?物種×土厚Species×soilthickness25.88??nsnsns物種×水分×土厚Species×water×soilthickness45.44??2.64?nsns
2.3.2胞間CO2濃度Ci、蒸騰速率Tr和氣孔限制值Ls
4月26日上午,十一屆全國人大常委會第二十六次會議召開聯組會議,就國務院關于農田水利建設工作情況的報告開展專題詢問。受吳邦國委員長的委托,聯組會議由全國人大常委會副委員長烏云其木格主持。
在淺土組,正常水分條件下(Wck),黑麥草的胞間CO2濃度(Ci)和蒸騰速率(Tr)較對照組分別下降17.7%和25.2%,氣孔限制值(Ls)與對照組相比顯著增加37.5%;在D1水分下,黑麥草的Ci與對照組無顯著差異,Tr和Ls與對照組相比分別顯著降低19.4%和27.8%;在D2水分下,黑麥草的Ci和Tr均與對照組無顯著差異,Ls顯著降低了23.8%。在深土組,Wck水分條件下,黑麥草的Ci和Ls均與對照組無顯著差異,Tr降低10.6%,但與對照組的差異不顯著;在D1水分下,Ci和Ls與對照組無顯著差異,Tr顯著降低了28.6%;在D2水分下,Ci、Tr和Ls均與對照組無顯著差異(圖4)。
在淺土組,正常水分條件下(Wck),葦狀羊茅的胞間CO2濃度(Ci)和蒸騰速率(Tr)較對照組分別顯著下降18%和56.8%,氣孔限制值(Ls)與對照組無顯著差異;在D1水分下,Ci顯著增加21%,Ls顯著降低32.4%,Tr與對照組無顯著差異;在D2水分下,Tr較對照組顯著下降57.7%,Ci和Ls與對照組無顯著差異。在深土組,葦狀羊茅的Ci、Tr和Ls在Wck、D1和D2這3種水分條件下均與對照組無顯著差異(圖4)。
其中αi,βi,δi,σi和ki(i=1,2,3)都是待定常數。令Tan、Tanh和exα1+yβ1+tδ1+σ1等項的系數為零可得
目前普遍認為,水分是喀斯特生態系統恢復中最重要的限制因子,有部分研究探討了喀斯特地區水分分布與土層厚度間的關系,但是很少有研究關注喀斯特土層空間異質性所導致的生境差異直接對植物的生長生理過程所產生的影響。在正常供水條件下,生境土層厚度的差異會對植物的形態建成(葉片解剖結構)和光合能力產生怎樣的影響?在全球降雨變化的大背景下,隨著水分減少,土層厚度對植物的影響是否存在規律性的變化,若存在,是什么樣的規律?對于這些問題,我們目前仍然無從知曉。因此,本試驗選擇黑麥草和葦狀羊茅作為試驗材料,通過盆栽模擬實驗,探究不同水分處理下兩種植物葉片解剖結構和光合生理特性對土層厚度的響應,以驗證以下假設:(1)水分充足條件下,兩種植物在淺土組受水分不足的刺激,具有更高的葉脈密度和氣孔密度,光合作用受到抑制;在深土組中,具有較低的葉脈密度和氣孔密度,光合作用受到促進;(2)隨著水分減少,兩種植物在淺土組受到的抑制作用和在深土組受到的促進作用逐漸增強;(3)與淺根植物黑麥草相比,無論在水分充足還是在水分不足的條件下,深根植物葦狀羊茅對深土組都表現出更強的適應性。
實時熒光定量PCR檢測結果(圖4A)顯示,轉染HSP27-siRNA后U266/BTZ細胞中HSP27 mRNA的表達水平較NC-siRNA組明顯下調(P<0.01),沉默效率為71.43%。蛋白質印跡法檢測結果(圖4B)同樣顯示,HSP27-siRNA組中HSP27蛋白的表達水平較NC-siRNA組明顯下調(P<0.01),沉默效率為65.67%。這一結果表明,轉染HSP27-siRNA后能明顯沉默HSP27基因的表達。
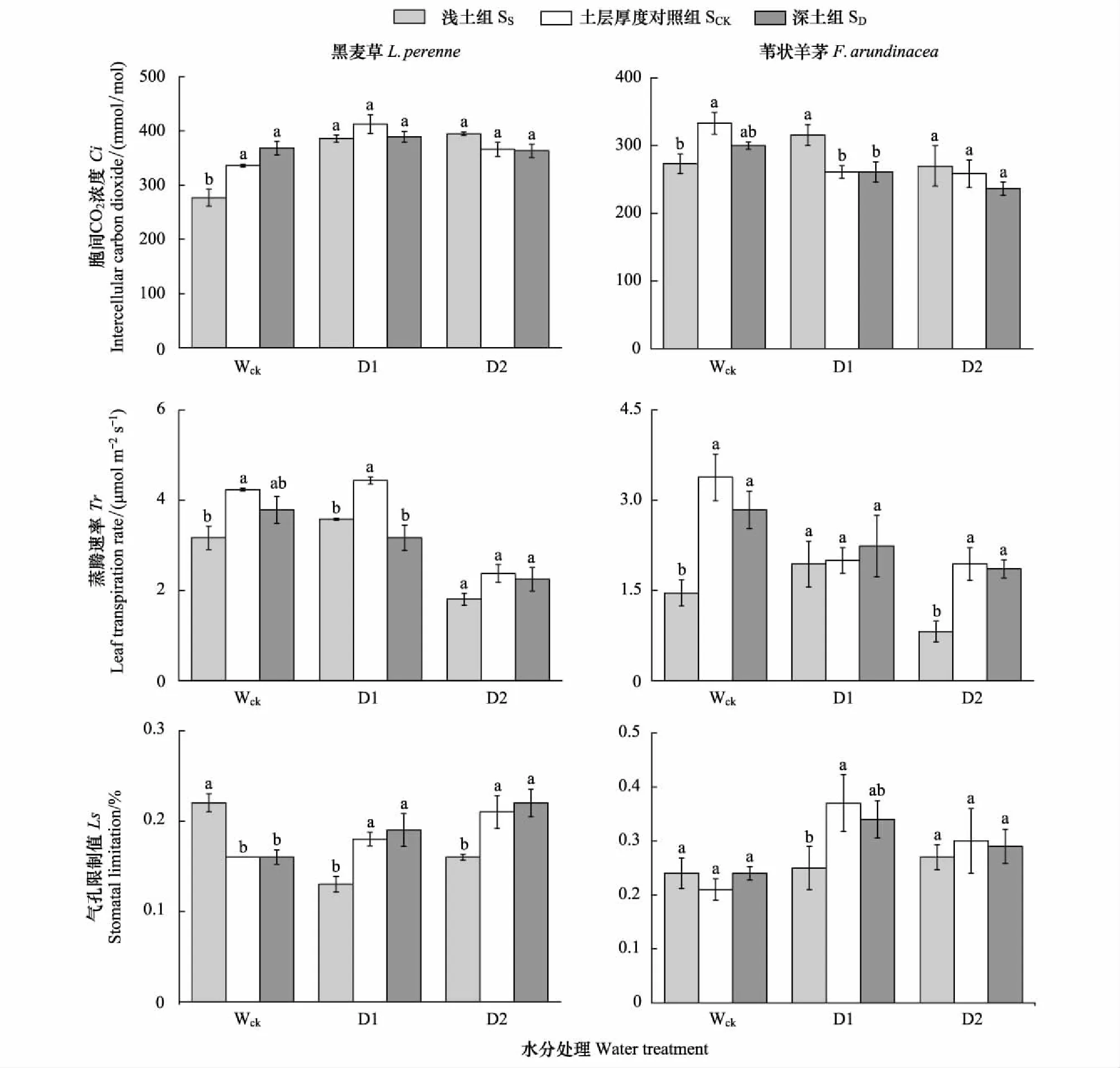
圖4 不同水分處理、不同土層厚度下黑麥草和葦狀羊茅的胞間CO2濃度Ci、蒸騰速率Tr和氣孔限制值Ls(均值±標準誤)Fig.4 The intercellular carbon dioxide (Ci),leaf transpiration rate (Tr) and stomatal limitation (Ls) of L. perenne and F. arundinacea under different soil thickness and water treatments (M±SD)
3 討論
氣候與植被間的相互作用受到土壤過程的調節,土層厚度被認為是一個復合因子,在土壤養分、水分和地下空間等方面扮演重要角色[37- 39]。在喀斯特地區,土壤的不連續分布使得該地區的土壤水分狀況對降雨變化的響應存在著強烈的空間異質性,這種異質性使得生長在不同小生境中的植物具有不同的適應性特征。
3.1 正常供水下(Wck),土層厚度對兩種草本葉片解剖結構和光合作用的影響
在正常供水條件下(Wck),不同厚度土層間的土壤含水量在很大程度上由其貯水能力的大小和蒸發作用的強弱決定。在試驗中,淺土組雖然貯水能力弱,但由于土量少,單位土壤體積獲得的水分多,且外界水分供給連續,使得其具有較高的土壤含水量,但總的貯水量仍然顯著低于對照組。在淺土組,兩種植物的葉脈密度與對照組沒有明顯差異,但黑麥草的近軸面氣孔密度和葦狀羊茅的兩面氣孔密度顯著增加。王學臣等[40]發現植物氣孔對水分虧缺非常敏感,水分脅迫時,在植物地上部尚未表現出任何可檢測的水分狀況變化時,氣孔性狀就已表現出受抑制的現象。淺土組水分的不足可能會刺激兩種植物氣孔的發育和生成[16],從而使其具有較高的氣孔密度,而兩種植物在淺土組葉面積的減少也可能是導致二者在淺土組表現出氣孔密度增加的原因。本研究進行植物光合參數的測定時,已經進行了69天的水分處理,植物生長已經進入到旺盛時期,對水分的需求量較大,淺土組的土壤水分貯藏顯然不能滿足植物正常的生長和蒸騰需求,所以兩種植物在淺土組的氣孔限制值Ls均有不同程度地升高,在減少水分散失的同時增加了CO2進入葉片細胞的阻力,導致胞間CO2濃度Ci和蒸騰速率Tr的下降,使得凈光合速率Pn降低,與假設一致。
在正常供水條件下(Wck),深土組較大的土壤體積(較多的土量)導致其單位土壤體積獲得的水分相對較少,土壤含水量顯著低于對照組,但仍在適宜植物生長的范圍之內,并且深土組總的土壤貯水量顯著高與對照組。在深土組,黑麥草的近軸面和遠軸面氣孔密度和葉脈密度均不同程度地低于對照組,這可能與其葉面積和葉片寬度的增加有關。而氣孔密度和葉脈密度的降低直接影響到植物水分供應和散失,所以黑麥草在深土組的蒸騰速率表現出低于對照組的情況。此外,在垂直方向上,隨著土層深度的增加,土壤含水量越大[23]。因此在深土組中,水分主要分布在較深層的土壤中,所以在一定程度上增加了黑麥草對水分利用的難度,也可能導致蒸騰速率的下降。并且有學者認為氣孔密度越大,越有利于植物在短時間內充分利用有效水分進行光合作用[41]。宋麗清等[20]發現較高的葉脈密度有利于支持較高的凈光合速率。因此,盡管黑麥草在深土組的氣孔限制值和胞間CO2濃度與對照組并無顯著差異,但葉脈密度和氣孔密度下降而引起植物水分運輸和利用的改變,最終使得黑麥草在深土組的凈光合速率低于對照組。
在深土組,葦狀羊茅的近軸面氣孔密度與對照組無顯著差異,而遠軸面的氣孔密度顯著降低,相比于近軸面,遠軸面的光強較弱[42],植物遠軸面的蒸騰速率也較低[20],而植物的氣孔密度與氣孔導度間具有負相關關系,因此葦狀羊茅遠軸面氣孔密度的降低在促進其遠軸面對CO2吸收的同時又避免了過多的水分散失。之前的研究普遍認為植物在水分條件較好的生境中具有較低的葉脈密度,在干旱生境中通過發展比較密集的維管束,增強植物的疏導能力和支撐能力[41]。而在本研究中,兩種植物在D2水分的淺土組具有較高的葉脈密度,并且伴隨著葉片寬度的顯著降低,因此,植物在干旱生境中表現出的較高的葉脈密度可能與葉片寬度的減小有關。而在正常供水條件下(Wck),葦狀羊茅在深土組的葉脈密度和葉片寬度均顯著地高于對照組,并且在本研究中多次發現兩種植物在水分條件較好的深土組中(Wck和D1水分下)具有更高的葉脈密度和葉片寬度,與之前一些研究的結果不符,造成這種結果的具體原因還有待探究。正常供水條件下,葦狀羊茅在深土組的胞間CO2濃度、蒸騰速率和氣孔限制值均與對照組無顯著差異,說明對照組和深土組良好的土壤水分條件都能很好地滿足葦狀羊茅生長和蒸騰對水分的需求,因此,在兩種土層厚度生境中(SCK和SD)植物自身具有相似的水分條件,因而具有相似的氣體調節表現。而葦狀羊茅在深土組表現出一定程度地凈光合速率增加,可能與深土組更高的養分可利用性有關。
3.2 輕度缺水時(D1),土層厚度對兩種草本葉片解剖結構和光合作用的影響
在D1水分條件下,由于水分減少,各個厚度土層的土壤水分狀況既受到自身水分貯藏能力的影響,也受到外部供水的制約。3種厚度的土層都屬于輕旱生境,淺土組較少的土量使其土壤含水量仍顯著高于對照組,但貯水量顯著低于對照組。相比于對照組,淺土組更為嚴重的干旱脅迫刺激了黑麥草的氣孔發育,其近軸面氣孔密度顯著地高于對照組。并且,淺土組嚴重的干旱脅迫導致黑麥草的光合器官受損,遭到不可逆的破壞,氣孔限制值顯著降低,較低的土壤水勢使得黑麥草在淺土組的蒸騰速率也顯著低于對照組,最終導致凈光合速率顯著下降。葦狀羊茅在淺土組的近軸面氣孔密度顯著降低,李海波[43]也發現,隨著干旱脅迫加重,水稻的氣孔密度先增加后降低。嚴重水分脅迫時,光合作用受到嚴重影響,減少了植株能量供應,抑制氣孔細胞的分裂生長和分化,導致氣孔密度下降;另一方面嚴重干旱脅迫還會使植物體內抗氧化酶的含量急劇下降,而活性氧含量增加較多,無法清除,細胞過早衰老死亡,從而使氣孔絕對數量減少,氣孔密度下降。而近軸面氣孔密度的減少也避免了植物過多的氣孔直接暴露在直射的太陽光下,可以有效地減少水分散失。葉片水勢可能受干旱脅迫的影響而降低[44],導致氣孔對氣體進出的控制力下降,氣孔限制值顯著降低,胞間CO2濃度顯著升高。葦狀羊茅在淺土組的蒸騰速率與凈光合速率均與對照組沒有顯著差異,可能與葦狀羊茅較高的抗旱性有關[45]。
窗墻面積比是指窗戶洞口總面積與同朝向建筑立面面積的比值。對于門窗,首先應符合功能要求,如充足的光線可避免白天燈光的使用,減少電能的損耗,同時也要滿足通風的要求;要使用保溫、隔聲,氣密性等級、水密性能好的符合國家標準的中空保溫玻璃,可以積極使用推廣一些先進的材料,如現在示范性項目中用的Low-E玻璃等,以達到節能的目的。
在深土組,黑麥草較淺的根系不能有效地利用深層土壤中的水分,所以相比于對照組,黑麥草在深土組獲取水分的難度增加,而葉脈密度的顯著增加也從側面印證黑麥草在深土組受到一定程度的水分虧缺的影響。黑麥草在淺土組通過增加氣孔限制值和降低蒸騰速率來保存體內有限的水分,但受水分不足的影響,凈光合速率仍出現顯著地降低。在深土組,葦狀羊茅的近軸面氣孔密度無明顯變化,遠軸面氣孔密度顯著下降,葉脈密度與葉片寬度均顯著高于對照組,說明深土組的土壤水分條件比對照組更利于葦狀羊茅的生長。在輕度干旱的深土組中,葦狀羊茅較大的根深有助于其從深層次的土壤中吸取水分供自身生長和蒸騰,較高的葉脈密度加強了其水分運輸的能力,所以葦狀羊茅在深土組的氣孔限制值略微降低,蒸騰速率略微高于對照組,凈光合速率表現出顯著增加。
3.3 重度缺水時(D2),土層厚度對兩種草本葉片解剖結構和光合作用的影響
在D2水分處理中,水分嚴重不足,外部供水的多少成為影響3種厚度土層土壤水分最為重要的因素。3種厚度土層的水分狀況均為中旱水平,但可能由于不同厚度土層的土壤蒸發作用強度不同,使得彼此土壤含水量之間存在一定差異,淺土組的土壤含水量表現出顯著降低,而深土組與對照組無顯著差異。此外,淺土組的土壤貯水量也顯著地低于對照組,所以植物在淺土組遭受的干旱脅迫更為強烈。黑麥草在淺土組受嚴重干旱脅迫的影響,近軸面氣孔密度顯著降低,葉脈密度顯著增加,嚴重的水分脅迫使得黑麥草的光合結構遭到破壞,氣孔限制值顯著降低,最后導致凈光合速率也出現明顯下降。葦狀羊茅在淺土組的近軸面氣孔密度與對照組無明顯差異,遠軸面氣孔密度顯著降低,葉脈密度表現出顯著地增加,說明葦狀羊茅在淺土組遭受到較為嚴重的干旱脅迫,蒸騰速率和凈光合速率表現出顯著地下降。
一對一死譯、硬譯的現象在公示語翻譯中十分普遍,這是一種不符合英語規范的中國式英語。例如,位于市中心的“桃園湖”被譯為“Peach Source Lake”,真是讓人啼笑皆非,應譯為“Taoyuan Lake”;楚雄市內一所KTV名為“大明星”,英譯為“Big Start KTV”,這是典型的死譯,應改為“Superstar”。又如,將“吸煙點”生硬翻譯為“Smoking Point”,事實上,此標識牌是要向吸煙的游客指明公共場所內允許吸煙的地方,譯為“Smoking Area”較為妥當。
在D2水分處理中,深土和對照組間的土壤含水量并沒有明顯差異。在研究中我們發現,由于外部供水太少,在干燥的土壤中,水分很難滲透到深層的土壤中去,而土壤表面的蒸騰作用太強導致水分大量散失,所以在對照和深土組中,水分主要集中分布在5—15 cm的土層中。在深土組,除了黑麥草的遠軸面氣孔密度顯著下降,兩種植物的其它葉片解剖結構和光合生理指標在對照和深土組間均沒有顯著差異。說明在水分極度缺乏的情況下,土層厚度的增加在緩解干旱脅迫對植物的影響方面并沒有明顯的作用。
4 結論
生境土層厚度的差異直接影響著植物的葉片解剖結構和光合特性,并且,隨著水分減少,土層厚度對植物的影響表現出規律性的變化。水分充足條件下,植物光合作用在淺土中受到抑制,在深土中,深根植物(葦狀羊茅)的光合作用受到促進,而淺根植物(黑麥草)受到輕微抑制;隨著水分減少,淺土對植物的抑制作用增強,深土對深根植物的促進作用和對淺根植物的抑制作用先增強后減弱;植物氣孔和葉脈性狀特征隨水分條件的變化在一定程度上與葉面積和葉片寬度的變化有關。因此,喀斯特地區土層厚度的空間異質性對植物的形態和光合特性具有重要作用,并且影響著生境中的植物對外部水分條件的響應表現。
[1] 王世杰, 季宏兵, 歐陽自遠, 周德全, 鄭樂平, 黎廷宇. 碳酸鹽巖風化成土作用的初步研究. 中國科學 D輯: 地球科學, 1999, 29(5): 441- 449.
[2] Fridley J D, Grime J P, Askew A P, Moser B, Stevens C J. Soil heterogeneity buffers community response to climate change in species-rich grassland. Global Change Biology, 2011, 17(5): 2002- 2011.
[3] 凡非得, 王克林, 熊鷹, 宣勇, 張偉, 岳躍民. 西南喀斯特區域水土流失敏感性評價及其空間分異特征. 生態學報, 2011, 31(21): 6353- 6362.
[4] 李周, 高凱敏, 劉錦春, 梁千慧, 陶建平. 西南喀斯特地區兩種草本對干濕交替和N添加的生長響應. 生態學報, 2016, 36(11):3372- 3380.
[5] Committee Report. Soil science in relation to water resources development: III. soil moisture conservation. Soil Sci. Soc. Amer. Proc., 1969, 33:480- 482.
[6] 朱守謙. 喀斯特森林生態研究Ⅰ. 貴陽: 貴州科學技術出版社, 1993.
[7] 郭柯, 劉長成, 董鳴. 我國西南喀斯特植物生態適應性與石漠化治理. 植物生態學報, 2011, 35(10): 991- 999.
[8] Shen W J, Reynolds J F, Hui D F. Responses of dry land soil respiration and soil carbon pool size to abrupt vs. gradual and individual vs. combined changes in soil temperature, precipitation, and atmospheric[CO2]: a simulation analysis. Global Change Biology, 2009, 15(9): 2274- 2294.
[9] Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt M B, Tignor M, Miller H L. Climate Change 2007: The Physical Science Basis. Cambridge: Cambridge University Press, 2007.
[10] 吳紹洪, 趙宗慈. 氣候變化和水的最新科學認知. 氣候變化研究進展, 2009, 5(3): 125- 133.
[11] 陳佳, 史志華, 李璐, 羅璇. 小流域土層厚度對土壤水分時空格局的影響. 應用生態學報, 2009, 20(7): 1565- 1570.
[12] 王勇, 梁宗鎖, 龔春梅, 韓蕊蓮, 于靖. 干旱脅迫對黃土高原4種蒿屬植物葉形態解剖學特征的影響. 生態學報, 2014, 34(16): 4535- 4548.
[13] Dunlap J M, Stettler R F. Variation in leaf epidermal and stomatal traits ofPopulustrichocarpafrom two transects across the Washington Cascades. Canadian Journal of Botany, 2001, 79(5): 528- 536.
[14] Fu Q S, Yang R C, Wang H S, Zhao B, Zhou C L, Ren S X, Guo Y D. Leaf morphological and ultrastructural performance of eggplant (SolanummelongenaL.) in response to water stress. Photosynthetica, 2013, 51(1): 109- 114.
[15] Xu Z Z, Zhou G S. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. Journal of Experimental Botany, 2008, 59(12): 3317- 3325.
[16] Zhao W S, Sun Y L, Kjelgren R, Liu X P. Response of stomatal density and bound gas exchange in leaves of maize to soil water deficit. Acta Physiologiae Plantarum, 2015, 37(1): 1- 9.
[17] Sam O, Jeréz E, Dell′Amico J, Ruiz-Sanchez M C. Water stress induced changes in anatomy of tomato leaf epidermes. Biologia Plantarum, 2000, 43(2): 275- 277.
[18] Scoffoni C, Rawls M, Mckown A, Cochard H, Sack LDecline of leaf hydraulic conductance with dehydration: relationship to leaf size and venation architecture. Plant Physiology, 2011, 156(2): 832- 843.
[19] Brodribb T J, Feild T S. Leaf hydraulic evolution led a surge in leaf photosynthetic capacity during early angiosperm diversification. Ecology Letters, 2010, 13(2): 175- 183.
[20] 宋麗清, 胡春梅, 侯喜林, 石雷, 劉立安, 楊景成, 姜闖道. 高粱、紫蘇葉脈密度與光合特性的關系. 植物學報, 2015, 50(1): 100- 106.
[21] Nardini A, Salleo S. Water stress-induced modifications of leaf hydraulic architecture in sunflower: co-ordination with gas exchange. Journal of Experimental Botany, 2005, 56(422): 3093- 3101.
[22] 曹娟云, 劉世鵬, 白重炎, 陳宗禮. 干旱脅迫條件下狗頭棗葉片解剖學結構比較研究. 安徽農業科學, 2008, 36(8): 3310- 3311.
[23] 張翠萍. 氮、磷對核桃苗木水分代謝的影響[D]. 北京: 中國林業科學研究院, 2014.
[24] 耿玉華. 多年生四倍體黑麥草. 農業知識, 2000(6): 38- 39.
[25] De Araujo M R A, 田有鳳. 葦狀羊茅種質農藝性狀的遺傳變異和相關. 草原與草坪, 1985(1): 31- 33.
[26] 魏寶祥. 葦狀羊茅在昆明地區的引種及種性研究初報. 云南農業大學學報, 2002, 17(3): 303- 306.
[27] 唐成斌, 吳佳海, 蔣明鳳. 貴州喀斯特地區牧草引種試驗評價. 草業科學, 2003, 20(12): 39- 42.
[28] 杜繼瓊. 三種冷季型草坪草抗旱性研究[D]. 楊凌: 西北農林科技大學, 2007.
[29] Weyers J D B, Johansen L G. Accurate estimation of stomatal aperture from silicone rubber impressions. New Phytologist, 1985, 101(1): 109- 115.
[30] Graham J D, Bergmann D C, Berry J A. An integrated model of stomatal development and leaf physiology. New Phytologist, 2014, 201(4): 1218- 1226.
[31] Sack L, Scoffoni C. Leaf venation: structure, function, development, evolution, ecology and applications in the past, present and future. New Phytologist, 2013, 198(4): 983- 1000.
[32] 蔡海霞, 吳福忠, 楊萬勤. 干旱脅迫對高山柳和沙棘幼苗光合生理特征的影響. 生態學報, 2011, 31(9): 2430- 2436.
[33] 曾旭, 陳芳清, 許文年, 王建柱. 向家壩工程擾動區6種先鋒草本植物的光合生理特性比較. 中國水土保持, 2010(12): 19- 22.
[34] 楊志民. 不同光照強度對冷季型草坪草越夏的影響[D]. 北京: 北京林業大學, 2006.
[35] 姚春艷. 果林下十種牧草的適應性比較研究[D]. 北京: 北京林業大學, 2011.
[36] 中華人民共和國水利部. SL 424- 2008 旱情等級標準. 北京: 中國水利水電出版社, 2009.
[37] Mcconnaughay K D M, Bazzaz F A. Is physical space a soil resource? Ecology, 1991, 72(1): 94- 103.
[38] Hess L, De Kroon H. Effects of rooting volume and nutrient availability as an alternative explanation for root self/non-self discrimination. Journal of Ecology, 2007, 95(2): 241- 251.
[39] Von Felten S, Schmid B. Complementarity among species in horizontal versus vertical rooting space. Journal of Plant Ecology, 2008, 1(1): 33- 41.
[40] 王學臣, 任海云, 婁成后. 干旱脅迫下植物根與地上部間的信息傳遞. 植物生理學報, 1992, 28(6): 397- 402.
[41] 劉球, 吳際友, 李志輝. 干旱脅迫對植物葉片解剖結構影響研究進展. 湖南林業科技, 2015, 42(3): 101- 104.
[42] 王曉琳, 李志強, 姜闖道, 石雷, 邢全, 劉立安. 散射光和直射光對高粱葉片光合功能的影響. 作物學報, 2012, 38(8): 1452- 1459.
[43] 李海波. 水分虧缺和鹽脅迫對水稻葉片氣孔及其他生理性狀的影響[D]. 沈陽: 沈陽農業大學, 2004.
[44] 紀文龍, 范意娟, 李辰, 魏靈芝, 姜金鑄, 李冰冰, 賈文鎖. 干旱脅迫下葡萄葉片氣孔導度和水勢動態的變化規律. 中國農業大學學報, 2014, 19(4): 74- 80.
[45] 孫鐵軍, 蘇日古嘎, 馬萬里, 武菊英. 10種禾草苗期抗旱性的比較研究. 草業學報, 2008, 17(4): 42- 49.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
