中華蜜蜂促分裂原活化蛋白激酶ERK-A基因的克隆與低溫表達分析
2018-03-22 03:43:20陳文鳳張衛星王紅芳胥保華
中國蜂業 2018年2期
陳文鳳 張衛星 王紅芳 胥保華
(山東農業大學動物科技學院,泰安 271018)
細胞對環境變化的反應部分是由一系列胞內信號途徑來誘導的,信號通路接替、放大并整合來自胞外刺激的信號,最終導致基因和生理的改變。昆蟲為適應極端環境,體內會啟動一系列代謝和調控反應。絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信號通路參與細胞的生長、發育、分化、凋亡等多種生理過程,不僅應答生物脅迫中的病原體感染,也對鹽、高溫、低溫、干旱等非生物脅迫有響應[1,2]。MAPK信號通路主要由ERK(extracellular signalregulated kinases)、JNK(The c-Jun amino-terminal kinases)和p38等并行的幾條信號通路組成。JNK主要參與細胞的轉錄調控,p38主要參與細胞對炎癥因子和環境脅迫(UV、溫度、缺血缺氧)的響應,而ERK信號通路主要參與調控細胞分裂過程。ERK信號通路在MAPK信號通路中最早被發現且研究得最為深入。ERK包括ERK-1和ERK-2兩種激酶,該途徑主要介導有絲分裂信號向胞內和核內傳遞,調控細胞的生長過程[3]。
應激反應是受細胞內信號系統調節的復雜過程,MAPK、G蛋白耦聯受體以及TGF-β等信號通路都參與應激反應[4,5]。當動物處于冷應激條件下時,動物機體作為一個整體發生反應,其神經內分泌系統、心血管系統、消化系統以及免疫系統等均發生一系列多系統、多層次的協調效應,以提高機體的適應能力和維持內環境的相對穩定。動物應激及動物對應激原的適應具有分子基礎,可以認為是一種基因行為[6,7]。任何機體酶和蛋白質的差異,其根本原因是基因表達的差異。在冷應激過程中主要表現為蛋白合成率的總體下降,但是某些蛋白的合成反而上升或嚴重下降,說明他們的表達受到了冷應激的特異性調節,并可能在冷應激過程中扮演重要角色[8]。ERK-A信號通路可將胞外刺激與胞內功能聯系起來,通過研究ERK信號通路調節機制可以了解冷應激對細胞活動的影響。研究發現,ERK通路通過調節類固醇和山梨醇的合成來終止家蠶幼蟲的滯育過程,表明ERK通路參與了昆蟲在低溫條件下的代謝調節[9,10]。
雖然對蜜蜂抵抗低溫的生理生化機制已有很多研究[11],但信號通路在中華蜜蜂低溫響應機制中的作用還鮮有報道,本研究通過將中華蜜蜂成蟲置于4℃環境下4 h,并在處理過程中取樣,取樣在3 min內完成,取樣后立即將蜜蜂放回原處理條件下。利用實時定量PCR檢測ERK-A基因的表達量,探討ERK-A基因是否在中華蜜蜂適應低溫環境中發揮作用。
1 材料與方法
1.1 供試蜂種
中華蜜蜂(簡稱中蜂,A.cerana cerana),飼養于山東農業大學實驗研究基地。
1.2 主要試劑
BL21(DE)和感受態細胞DH5α購自北京全式金生物技術有限公司,克隆載體pEasy-T3購于TransGen公司,原核表達載體Pet-30a(+)由本實驗室提供。TaqDNA聚合酶、T4 DNA連接酶、dNTP、DNA Marker、SYBR PrimeScriptTM RT-PCR Kit、各種限制性內切酶購于寶生物工程(大連)有限公司(TaKaRa);瓊脂糖凝膠DNA回收試劑盒購于Solarbio公司;質粒提取試劑盒Plasmid Mini Kit購于OMEGA公司;引物合成和測序由上海生工生物工程股份有限公司完成。
1.3 PCR引物(表1)

表1 PCR引物
1.4 中華蜜蜂ERK-A基因的生物信息學分析
利用DNAMAN 6.0軟件將ERK-A基因的開放讀碼框翻譯成氨基酸序列,得到蛋白,在NCBI網站(http://www.ncbi.nlm.nih.gov/)上將ERK-A同其它物種進行Blast同源比對。在亞細胞定位預測網站(http://psort.hgc.jp/form.html)上預測該蛋白質在細胞中的位置,利用MEGA 5軟件對不同物種ERK-A基因序列進行多重比對,通過NJ(neighbor joining)方法構建系統進化樹。
1.5 中華蜜蜂的低溫處理
選取正常蜂群中的中華蜜蜂320只,分成4組,每組80只,隨機選取三組在4℃冰箱中放置15 min、30 min、1 h、2 h、2.5 h、3 h、4 h,另一組放置于25℃恒溫恒濕培養箱中,處理后直接將蜜蜂凍存在液氮中,將25℃飼養的蜜蜂成蟲作為對照,不作低溫處理直接凍存在液氮中,以單只蜜蜂為一個樣品,每個處理共3只蜜蜂(即3次重復)。
1.6 中華蜜蜂總RNA的提取、cDNA的合成以及實時熒光定量PCR
每群選取2只蜜蜂液氮研磨,Trizol法提取總mRNA,然后使用反轉錄試劑盒(TaKaRa:DRR037A)立即反轉錄為cDNA,調節樣品cDNA濃度于相同水平后,-20℃保存備用。qRT-PCR取1000 ng cDNA加入到20 μl熒光定量體系中,按照熒光定量試劑盒(TaKaRa)操作指南,用7500 Real-Time PCR儀(ABI 7500,USA)檢測目的基因相對表達量。反應程序:預變性95℃,10 s;變性95℃,5 s;退火60℃40 s,40個循環,熔解曲線添加,1個循環。目的基因引物設計參考序列來自于NCBI數據庫,以action為內參,采用Primer 5.0進行引物設計,委托生工生物科技有限公司合成引物。
2 結果與分析
2.1 中華蜜蜂ERK-A基因cDNA的克隆(圖1)
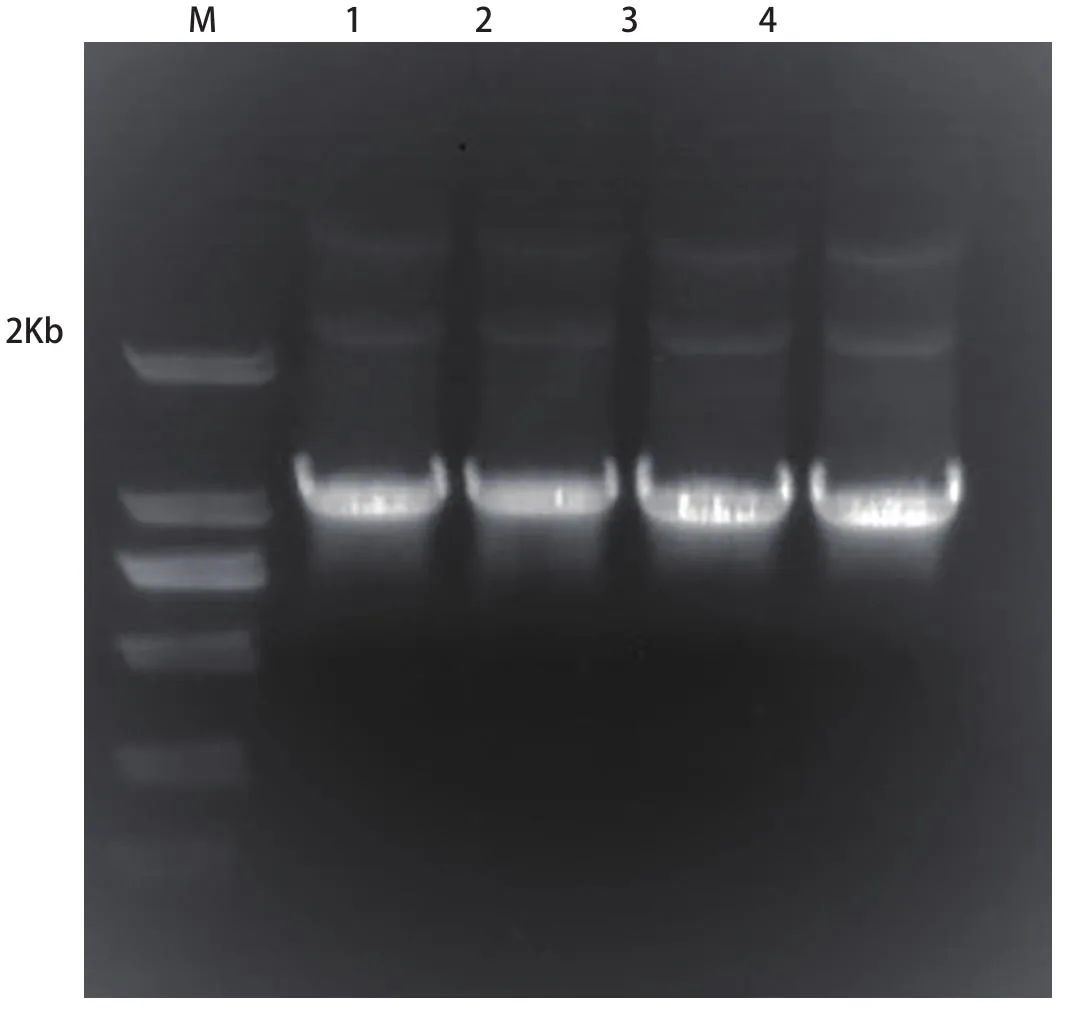
圖1 ERK-A基因克隆
2.2 中華蜜蜂ERK-A基因cDNA的序列分析
通過DNAMAN 6.0軟件進行分析獲得中華蜜蜂ERK-A基因的開放讀碼框,含有1098 bp的堿基,編碼365個氨基酸,編碼蛋白質命名為ERK-A,其分子量為42 kDa,等電點為6.03。ERK-A1/2 是細胞對外界信號產生應答的上游樞紐,通過亞細胞定位預測(http://psort.hgc.jp/form.html),ERK-A主要存在于細胞質中,且無信號肽編碼序列,無跨膜結構(圖2)。ERK-A具有MAPK超家族的序列特征,即存在激酶的保守區和磷酸化位點(下劃線標的TEY序列)。

圖2 中華蜜蜂ERK-A的核酸與氨基酸序列
2.3 中華蜜蜂ERK-A蛋白的同源性分析

圖3 中華蜜蜂與其他物種的同源比對
將ERK-A的氨基酸序列在NCBI網站進行Blast比對,結果表明ERK-A與同為蜜蜂科Apis mellifera(GenBank登錄號:XP_393029.2)的相似性最高,一致性99.18%,與Harpegnathos saltator (GenBank登錄號:XP_017789097.1)的一致性97.81%。與Eufriesea Mexicana和Melipona quadrifasciata的一致性都在97%以上(圖3),說明該基因在進化上高度保守。
2.4 中華蜜蜂ERK-A基因的三維結構預測及磷酸化位點分析
MAPK的激活都是通過TEY基序中蘇氨酸(T)和酪氨酸(Y)的同時磷酸化實現的。利用SWISSMODEL(http://www.swissmodel.expasy.org/)預測ERK-A激酶的三級結構(圖4),可以看出,N端結構域主要由β折疊組成,C端結構域主要由α螺旋組成,兩個結構域之間的裂痕(即無規則卷曲)為ATP結合的激活環,TEY基序就位于激活環內[12]。
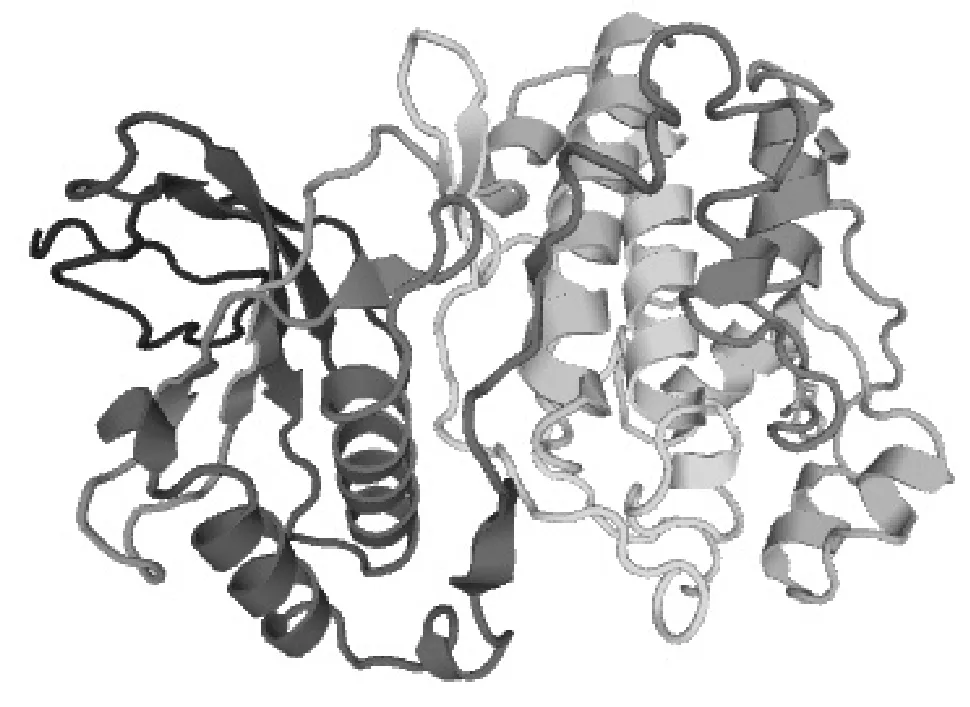
圖4 ERK-A的三維結構剖面圖
2.5 中華蜜蜂ERK-A激酶與其他物種ERK-A激酶的系統樹構建
從NCBI網站下載相近物種的部分ERK-A激酶序列,利用MEGA軟件,以酵母Saccharomyces merevisiae的ERK-A作為外群,通過Test Neighbor Joining Tree方法構建以氨基酸序列為基礎的系統發生樹(圖5),結果表明:中華蜜蜂ERK-A和同為蜜蜂科的意大利蜜蜂Apis mellifera的ERK-A聚為一支,與蟻科的收獲蟻Pogonomyrmex barbatus和擬步甲科的赤擬谷盜Tribolium castaneum親緣關系較近。小鼠ERK-A和人ERK-A聚為一支,昆蟲ERK-A與哺乳動物ERK-A有更近的起源。

圖5 昆蟲ERK-A的系統進化樹
2.6 低溫脅迫下中華蜜蜂ERK-A基因的表達規律
為了研究ERK-A基因的表達是否對低溫脅迫做出響應,進行實時熒光定量PCR檢測。熒光實時定量PCR結果顯示,在低溫脅迫下ERK-A的表達均有顯著上調。在4℃脅迫30 min后,ERK-A的表達量就達到對照的2倍,2 h時達到最高峰是對照的4倍,此后雖有下降,但仍保持較高水平(圖6)。低溫表達分析說明,ERK-A基因的表達受到低溫的誘導,可能在中華蜜蜂應對低溫脅迫時發揮對細胞的調控作用。
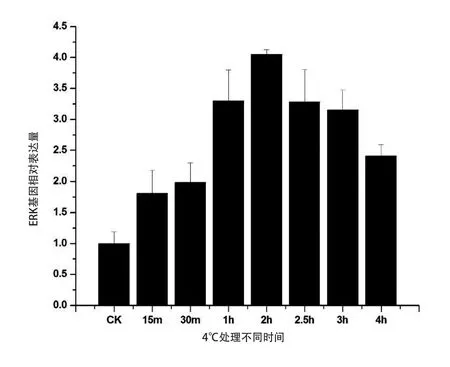
圖6 低溫條件下ERK-A基因表達的變化
我們從蜜蜂轉錄組數據庫中獲得ERK-A基因的cDNA序列的完整開放閱讀框長1098 bp,編碼365個氨基酸,編碼蛋白ERK-A的分子量為42 kDa,等電點為6.03。同源性分析發現,該基因在進化上十分保守,在氨基酸水平上,與相近物種的ERK-A的一致性都達到97%以上,與意大利蜜蜂ERK-A的一致性達99.18%,表明ERK-A在生物體中都具有重要功能。本研究發現ERK-A在4℃呈上調表達,說明ERK-A通路可能參與了中華蜜蜂耐寒機制的調控。
3 結論與討論
真核細胞已經形成了特殊的信號轉導通路來響應細胞外的各種刺激,絲裂原蛋白激酶MAPK組成了一個絲氨酸/蘇氨酸蛋白激酶家族,通過磷酸化作用激活受體,調節細胞的增殖、分化和脅迫響應[13]。在比較生物化學領域,MAPK信號通路在代謝調節和生化適應方面的研究還比較少見,最初關于MAPK信號通路在缺氧狀態、熱應激和冷應激等方面的代謝應答的研究也僅僅觸及其表面[14]。
通常認為,ERK通路主要參與細胞存活和生長繁殖,其有效激活信號分子包括生長因子、激素、神經遞質、鈣離子等[15],而p38和JNK主要參與生長抑制和細胞凋亡,其有效激活因素有溫度、缺血缺氧等各種環境刺激[16]。ERK主要被生長因子、多肽類激素以及神經遞質激活,控制細胞增殖、分化、生存和凋亡。本研究中,ERK-A基因的表達受低溫誘導,可能與ERK、p38和JNK三條通路的相互作用有關[17,18]。
本研究從中華蜜蜂轉錄組數據中獲得ERK-A基因的cDNA,該基因在低溫條件下上調表達,說明在中華蜜蜂中ERK通路可能參與了低溫下的信號轉導。昆蟲的一生會經常性受到來自環境的各種各樣的刺激,昆蟲的誘導表達適應機制提示人們,昆蟲對環境的適應是一個能動的過程。昆蟲通過啟動與抗低溫有關的基因并使之表達,給出逆境保護措施,從而協助昆蟲度過難關或逆境。因此,研究與冷應激密切相關的基因,探討如何迅速誘導昆蟲冷適應機制的高效運作,提高昆蟲在不利環境下的存活率,將是現在乃至將來昆蟲學家應傾力研究的課題。此外,不同類型的應激可以誘導不同的應激蛋白,后者即可用來判斷昆蟲受到何種應激和應激的程度。研究冷應激過程中發生改變的這些蛋白用以衡量應激的程度,作為指導我們工作和生產的一個標準。因此,從分子水平上研究冷應激的分子機制,將具有極大的基礎研究和實踐應用價值,研究MAPK信號通路對低溫脅迫的調控,有助于全面了解昆蟲的耐寒機制。
[1] Atay O, Skotheim J M.Spatial and temporal signal processing and decision making by MAPK pathways [J].Journal of Cell Biology,2017, 216(2):317.
[2] Dent P, Yacoub A, Fisher PB, et al.MAPK pathways in radiation responses [J].Oncogene, 2003, 22(37):5885-5896.
[3] Lim W, Park S, Bazer F W, et al.Apigenin reduces survival of choriocarcinoma cells by inducing apoptosis via the PI3K/AKT and ERK1/2 MAPK pathways [J].Journal of Cellular Physiology, 2016,231(12):2690.
[4] Sonna L A, Fujita J, Gaffin SL, et al.Invited review: Effects of heat and cold stress on mammalian gene expression [J].Journal of Applied Physiology, 2002, 92(4):1725.
[5] 楊發青,錢令嘉,王萬銀,等.寒冷適應差異表達基因的研究[J].生理學報,2003, 55(3):360-363.
[6] Shinozaki K, Yamaguchishinozaki K, Seki M.Regulatory network of gene expression in the drought and cold stress responses [J].Current Opinion in Plant Biology, 2003, 6(5):410-417.
[7] 柳巨雄,欒新紅,楊煥民,等.冷休克蛋白表達的分子機制[J].中國獸醫學報,2003, 23(6):607-610.
[8] 姜冬梅,李士澤,姜書磊,等.冷應激動物相關基因表達的研究進展[J].環境與健康雜志,2006, 23(5):471-473.
[9] Fujiwara Y, Tanaka Y, Lwata K, et al.ERK/MAPK regulates ecdysteroid and sorbitol metabolism for embryonic diapause termination in the silkworm, Bombyx mori [J].Journal of Insect Physiology, 2006, 52(6):569-575.
[10] Huang, Hao-Jen, Fu, et al.Expression of Oryza sativa MAP kinase gene is developmentally regulated and stress-responsive [J].Physiologia Plantarum, 2002, 114(4):572.
[11] 陳琳,徐新建,王青,等.低溫20℃對意大利蜜蜂未受精卵發育的影響[J].應用昆蟲學報,2016, 53(3):574-580.
[12] Widmann C, Gibson S, Jarpe MB, et al.Mitogen-activated protein kinase: conservation of a three-kinase module from yeast to human [J].Physiological Reviews, 1999, 79(1):143-180.
[13] Xu J, Zhang S.Mitogen-activated protein kinase cascades in signaling plant growth and development [J].Trends in Plant Science,2015, 20(1):56-64.
[14] Cowan K J, Storey K B.Mitogen-activated protein kinases: new signaling pathways functioning in cellular responses to environmental stress [J].Journal of Experimental Biology, 2003, 206(Pt 7):1107.
[15] Johnson G L, Lapadat R.Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases [J].Science, 2002, 298(5600): 1911-1912.
[16] Ono K, Han J.The p38 signal transduction pathway: activation and function [J].Cellular Signalling, 2000, 12(1):1-13.
[17] Wang J Y, Chen S P, Gao Y H, et al.Effect of repeated electroacupuncture intervention on hippocampal ERK and p38 MAPK signaling in neuropathic pain rats [J].Evidence-Based Complementray and Alternative Medicine, 2015, 2015(12):641286.
[18] Sinha A K, Jaggi M, Raghuram B, et al.Mitogen-activated protein kinase signaling in plants under abiotic stress [J].Plant Signaling & Behavior, 2011, 6(2):196-203.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
汽車工程學報(2017年2期)2017-07-05 08:13:02
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
