鹽脅迫環境下接種根際促生細菌對白蠟樹根際生物學特征及其生長的影響
2018-03-29 03:14:50井大煒馬海林劉方春杜振宇馬丙堯昝林生賈海慧郭英華丁洪星
水土保持通報 2018年1期
井大煒, 馬海林, 劉方春, 杜振宇, 馬丙堯, 昝林生, 賈海慧, 郭英華, 丁洪星
(1.德州學院 資源環境與規劃學院, 山東 德州 253023; 2.山東省林業科學研究院, 山東 濟南 250014; 3.德州學院生態與園林建筑學院, 山東 德州 253023; 4.濟南市國有苗圃, 山東 濟南 250100; 5.濟南花木聯合開發公司, 山東 濟南 250100)
土壤鹽漬化是造成當今世界土地退化的重要原因之一,鹽漬化土壤在全球分布的面積非常廣泛,已有超過1.00×109hm2的鹽漬化土地急需有效、可持續地管理[1-2]。由于灌溉方式不當、濫砍亂伐、破壞草原及盲目施肥等原因,全球每年的鹽漬化土地面積仍在快速增長[3]。中國現有鹽漬化土地面積約3.63×107hm2,占全國可利用土地面積的4.88%[3]。解決這一問題的根本途徑在于恢復植被,改善土壤結構,這是關系到國民經濟可持續發展與和諧社會構建的重大課題。多年來,人們圍繞鹽堿地改良開展了大量的探索研究工作,也積累了許多鹽堿地改良技術,比如土工排水體排水技術、隔鹽層技術、集雨洗鹽技術與表面覆蓋技術等[4]。
植物根際促生細菌(plant growth-promoting rhizobacteria, PGPR)是指生存在植物根圈范圍內,對植物生長有促進或對病原菌有拮抗作用的有益細菌的統稱,對植物生長與病害防治有極其重要的作用[5]。PGPR對土壤中非寄生性根際有害微生物與有害病原微生物能起到生防作用,并產生利于植物生長的代謝產物,進而促進植物的生長發育[6]。PGPR還具有降解土壤中污染物的作用,并減少生產中使用化肥、農藥帶來的環境污染與土壤板結等問題[7]。有研究表明[8],接種產ACC脫氨酶或產生長素的PGPR,可顯著促進作物生長。也有研究認為[5],接種PGPR對植物根際生態環境的改善,尤其是植物根系生長有顯著促進作用。同時,也有關于在干旱生境下接種產細胞分裂素的PGPR,并能顯著增強林木干旱耐受性的研究報道[9]。可見,PGPR的開發研究正逐漸成為現代林農領域的研究熱點,但有關PGPR在鹽脅迫環境中對根際生物學特征影響的研究甚少。
根際是距離根系表面0—4 mm的土壤區域[10]。林木根際是由樹木細根—土壤—微生物—酶組成的一個特殊微生態系統,也是各種養分、水分及各種物質進入根系參與物質循環和能量轉化的重要場所之一,是與原土體差異較大的特殊土壤微區[5]。土壤微生物作為土壤生態系統的重要組成部分,推動著土壤的碳、氮循環[11]。土壤微生物種類、數量在很大程度上影響著微生物的活性。土壤鹽漬化不僅可以直接影響微生物的活性,還能通過改變土壤的理化性狀影響微生物的生存環境,進而影響微生物的種類與數量[3]。土壤酶也是土壤有機成分中極其活躍的部分,參與了土壤中大部分的生物化學過程,對物質循環與養分形成、積累有抑制或促進作用[10,12]。因此,近年來許多學者認為,積極改善土壤微生態環境,充分協調根際中微生物與酶,有利于提高作物在逆境下的耐受能力。白蠟樹(Fraxinuschinensis)具有較強的抗逆性能,在鹽堿地區土壤改良與生態體系構建中發揮著重要作用[3]。為此,本文選取具有促生功能的陰溝腸桿菌為供試菌株,以白蠟樹為試材,開展不同鹽脅迫梯度下接種PGPR對白蠟樹根際生物學特征的影響研究,旨在明確PGPR對白蠟樹耐鹽性的改善效果,為鹽脅迫環境下PGPR的推廣應用提供理論依據。
1 材料與方法
1.1 試驗地點與供試材料
試驗地點設在山東省德州市農業科學研究院試驗大棚,供試土壤為潮土,土壤速效氮含量32.87 mg/kg,速效磷含量28.06 mg/kg,速效鉀含量86.50 mg/kg,有機質11.93 g/kg,可溶性鹽分為0.069 5%,pH值8.06。供試白蠟樹為1年生實生苗,株高(8.72±3.46)cm,地徑(0.19±0.02)cm。
利用梯度稀釋法與三區劃線法從白蠟樹的根際中分離、純化出細菌分離物,然后參考Liu等[9]的生物法(小麥葉片保綠和蘿卜子葉增重)篩選出1株促生效果較好的植物根際促生細菌,綜合其生理生化特征與16 S rRNA基因序列分析,鑒定為:陰溝腸桿菌(Enterobactercloacae)TIA062。將該菌株接入牛肉膏蛋白胨培養基(牛肉膏0.3%,蛋白胨1%,氯化鈉0.5%,瓊脂2%,pH值7.0~7.2,質量百分比)中,于37 ℃,180 r/min條件下振蕩培養2~3 d。然后,接種到液體培養基中(乳糖2%,蛋白胨2%,K2HPO40.15%, MgSO4·7 H2O 0.02%,CaCl20.01%,pH值6.2~6.8),接種量10%,于30~35 ℃搖床培養,轉速180 r/min,培養12 h,利用平板計數法測定每毫升含菌量,計算菌液濃度,統一稀釋至2.00×108cfu/ml。
1.2 試驗設計
于2016年6月4日選取長勢基本一致的苗木進行盆栽,盆高20 cm,寬30 cm。每盆稱土9.8 kg。
試驗共設7個處理,分別為: ①CK:對照,不加NaCl和PGPR; ②LS:輕度鹽脅迫,鹽分質量分數0.2%; ③MS:中度鹽脅迫,鹽分質量分數0.4%; ④SS:重度鹽脅迫,鹽分質量分數0.6%; ⑤LS+P:在處理2的基礎上加入PGPR菌液; ⑥MS+P:在處理3的基礎上加入PGPR菌液; ⑦SS+P:在處理4的基礎上加入PGPR菌液。重復5次。處理5,6,7是分別取PGPR菌液10 ml,稀釋至150 ml,均勻澆灌于白蠟樹苗根系周圍;然后澆水至田間持水量的75%,每天定時澆水,維持土壤含水量為田間持水量的65%~70%。
1.3 測定項目與方法
2016年9月29日首先用游標卡尺和卷尺測量白蠟樹苗的地徑、株高,然后采用剝落分離法[12]采集根際土,并將所取根際土樣混勻后分為2份:一份鮮樣迅速用于土壤微生物數量與微生物量碳、氮的測定;另一份自然風干,過1 mm篩后用于土壤酶活性的測定。
土壤微生物數量采用稀釋平板計數法,其中細菌數、放線菌數與真菌數的測定分別采用牛肉膏蛋白胨瓊脂培養基、改良高氏1號培養基+重鉻酸鉀與馬丁培養基+孟加拉紅+硫酸鏈霉素[5]。微生物量碳、氮采用氯仿熏蒸K2SO4浸提法測定[5]。土壤脲酶、多酚氧化酶、過氧化氫酶和蔗糖酶活性的測定分別采用苯酚鈉—次氯酸鈉比色法、鄰苯三酚比色法、高錳酸鉀滴定法和Na2S2O3滴定法[13]。根系體積采用排水法測定;根系總吸收面積、活躍吸收面積采用甲烯藍吸附法[14]測定。
1.4 數據處理
采用SPSS 19.0軟件進行數據統計分析,采用雙因素方差分析(two-way ANOVA)與最小顯著差異法(LSD)比較不同處理組間的差異,顯著性水平設定為α=0.05。
2 結果與分析
2.1 根際微生物數量與微生物量碳、氮
由表1可見,白蠟樹根際微生物以細菌為主,放線菌次之,真菌最少,其所占微生物總量的比例分別為70.78%~79.17%,20.23%~28.78%和0.44%~0.60%。在鹽脅迫生境中,白蠟樹根際的微生物數量顯著降低,且微生物數量隨土壤鹽分的增加降幅增大。與CK相比,LS,MS和SS處理的細菌數分別下降7.49%,19.81%和38.93%。當接種PGPR后,各處理對應的細菌和放線菌數量顯著增加,而真菌數變化較小。LS+P處理的細菌數、放線菌數和微生物總量均達最高值,并顯著高于其他處理,分別較CK提高14.64%,24.01%和17.04%。土壤微生物總量的變化趨勢與細菌數一致。從表1還可知,不同處理的微生物量碳、氮含量呈現出基本一致的變化規律,加菌液處理均高于對應的不加菌液處理。以上分析可知,土壤鹽脅迫會導致白蠟樹根際微生物數量與微生物量碳、氮含量顯著下降,而接種根際促生細菌可減弱鹽分脅迫的影響。

表1 不同處理對白蠟樹根際微生物數量與微生物量碳、氮含量的影響
注:CK:對照,不加NaCl和PGPR; LS:輕度鹽脅迫,鹽分質量分數0.2%; MS:中度鹽脅迫,鹽分質量分數0.4%; SS:重度鹽脅迫,鹽分質量分數0.6%; LS+P:在SS的基礎上加入PGPR菌液; MS+P:在LS+P的基礎上加入PGPR菌液; SS+P:在MS+P的基礎上加入PGPR菌液。數據為平均值±標準誤; 同列數據后不同小寫字母表示處理間差異顯著(p<0.05)。下同。
2.2 根際酶活性
從表2可見,不同鹽脅迫梯度均顯著降低了白蠟樹根際的脲酶、多酚氧化酶、過氧化氫酶和蔗糖酶活性;當接種PGPR后,對應的脲酶、多酚氧化酶和蔗糖酶活性顯著升高,而過氧化氫酶活性的增幅較小。以LS+P處理的脲酶、多酚氧化酶和蔗糖酶活性為最高,與CK差異不顯著,但顯著高于其他處理,其中脲酶活性分別較LS,MS,SS,MS+P和SS+P處理均提高16%以上。MS+P處理的脲酶、蔗糖酶活性與LS處理無顯著性差異,而多酚氧化酶、過氧化氫酶活性明顯低于LS處理。SS+P處理的蔗糖酶活性與MS處理差異未達顯著水平,而脲酶、多酚氧化酶活性顯著低于MS處理。由此可見,隨土壤鹽分梯度的增加,白蠟樹根際酶活性表現出遞減的變化趨勢,而接種PGPR能使對應的脲酶、多酚氧化酶和蔗糖酶活性顯著升高。
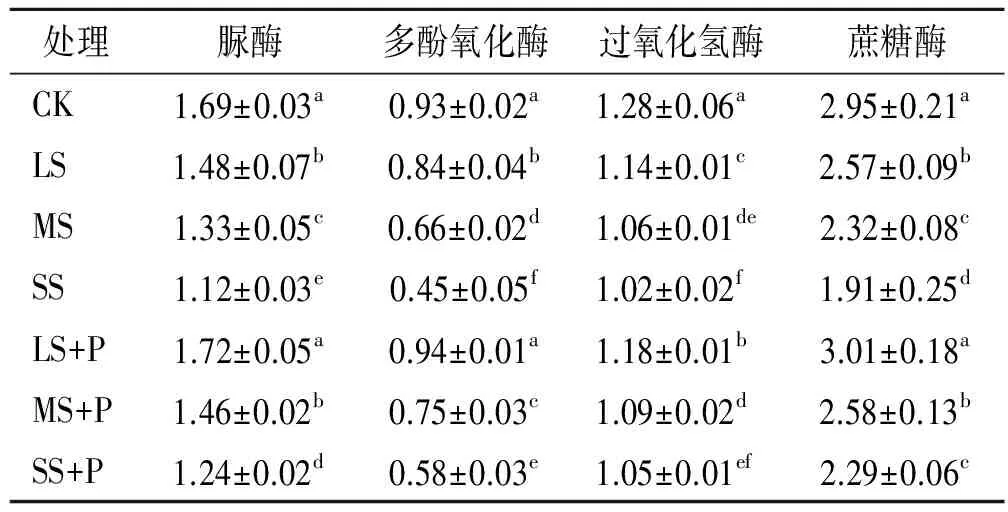
表2 不同處理對白蠟樹根際酶活性的影響 ml/(g·h)
2.3 根系吸收面積
從表3可知,土壤鹽分對白蠟樹根系產生了較大影響。隨著土壤鹽分的增加,白蠟樹的根體積、總吸收面積和活躍吸收面積呈逐漸降低的趨勢,且下降幅度越來越大。當接種PGPR后,加菌液處理的根體積、總吸收面積和活躍吸收面積均高于對應的不加菌液處理,其中LS+P處理的根體積、總吸收面積與CK差異不顯著,而活躍吸收面積顯著高于CK 15.97%;同時,LS+P,MS+P和SS+P處理的根系指標呈遞減的變化趨勢。從表3還可見,鹽脅迫使活躍吸收面積/總吸收面積的比值較對照有上升趨勢;接種PGPR后,對應的比值繼續升高,以LS+P處理為最高,并與其他處理差異達顯著水平。數據分析表明,在不同梯度的鹽脅迫環境下接種根際促生細菌能不同程度地提高白蠟樹根系的總吸收面積與活躍吸收面積,其中在輕度鹽脅迫下接種的效果最顯著,這能更利于促進根系對水分、養分的吸收。
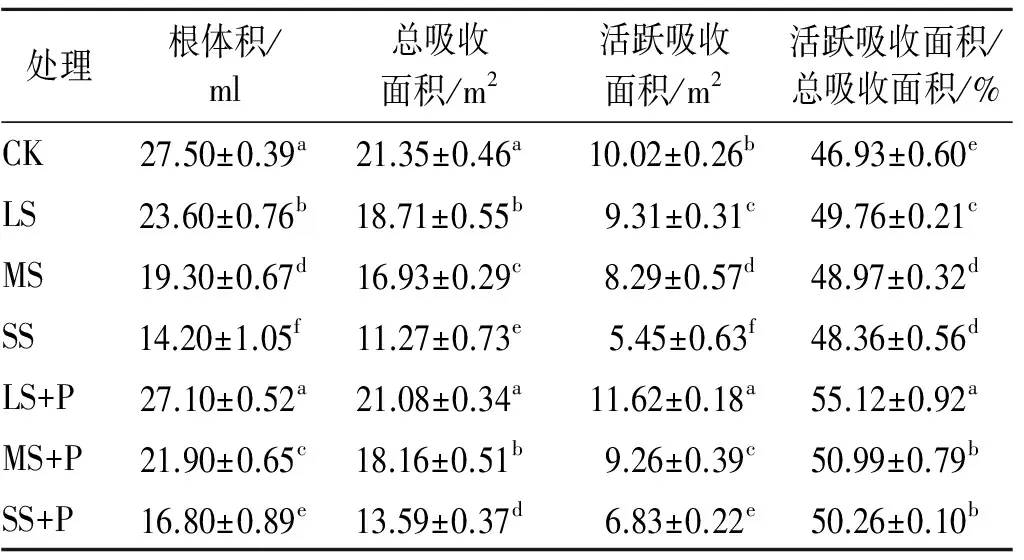
表3 不同處理對白蠟樹根系總吸收面積與活躍吸收面積的影響
2.4 地徑、株高
從圖1可見,與對照相比,輕度、中度和重度鹽脅迫使白蠟樹的地徑、株高顯著降低,且脅迫程度越高,下降幅度也越大。而接種PGPR使白蠟樹的地徑、株高呈明顯升高趨勢,其中LS+P處理的地徑、株高與CK無顯著性差異,但顯著高于其他處理,地徑分別較LS,MS,SS,MS+P和SS+P處理高出13.80%,30.01%,95.64%,15.58%和69.57%,株高分別高出19.08%,41.41%,124.84%,24.11%和71.84%。同時,MS+P處理的地徑、株高顯著高于SS+P處理。綜上所述,土壤鹽脅迫可明顯抑制白蠟樹地徑、株高的生長,而接種根際促生細菌能顯著促進白蠟樹的生長,其中在輕度鹽脅迫生境下接種根際促生細菌的促長效應最明顯。

圖1 不同處理對白蠟樹地徑與株高生長的影響
3 討 論
根際是植物—土壤—微生物—酶相互作用的特殊生態環境,根際微生物數量和種類是表征土壤肥力的主要生物學指標[15]。本試驗研究表明,鹽脅迫環境下白蠟樹根際細菌數占微生物總量的比例為70.78%~79.17%,放線菌為20.23%~28.78%,而真菌僅占0.44%~0.60%。鹽脅迫下接種根際促生細菌并沒有大幅度改變土壤中三大功能微生物的總體比例。鹽脅迫導致根際微生物數量顯著下降,且隨著鹽脅迫程度的加劇,降幅呈增加趨勢,這與莫俊杰等[16]對鹽脅迫下甘蔗根際微生物量的研究結論相似。這說明白蠟樹根際中的微生物種群對鹽分濃度變化十分敏感,土壤鹽脅迫改變了土壤微生物的生長繁殖環境,從而使其種群數量受到了影響[17]。本試驗還得出,接種PGPR能使對應的微生物數量顯著提高,其原因可能是由于接種陰溝腸桿菌可分泌生長素,促進白蠟樹根系的生長,并刺激根系活性,促使其根系分泌物的增多,從而為微生物的生長繁殖提供了營養物質。其中在輕度脅迫下接種的效果最顯著,其次為中度或重度脅迫下接種,表明低鹽環境可能更利于根際促生細菌的定植與繁殖生長,也說明根際促生細菌的影響作用與土壤鹽分梯度有一定內部關聯。
土壤酶主要來自微生物和植物根系的分泌。土壤中鹽分的增加不僅影響到植物的生長發育,而且對土壤自身的物理化學性狀也產生不良效應[18]。土壤酶活性的變化可從一個側面反映土壤性狀的改變[19]。本研究表明,在鹽脅迫生境中,白蠟樹根際的脲酶、多酚氧化酶、過氧化氫酶和蔗糖酶活性顯著降低,且隨著鹽脅迫程度的加劇,下降幅度越來越大。這與周德平等[17]關于鹽脅迫對蔬菜地土壤酶活性的研究結果相一致。其原因可能在于鹽分改變了土壤環境,鹽分產生的滲透脅迫與離子毒害均會抑制土壤酶的活性。當接種PGPR后,對應的脲酶、多酚氧化酶和蔗糖酶活性顯著升高。究其原因在于: ①PGPR能顯著提高白蠟樹根際的微生物數量,這可能會增加微生物向土壤中分泌的土壤酶量[19]; ②該種促生菌可產生長素,能刺激白蠟樹根系分泌物的增多。
當植物受到鹽脅迫時,根系首先感應到根際的變化。植物根系是活躍的吸收器官與合成器官,根系吸收面積可反映根系吸收水分與養分的能力,而根系活躍吸收面積則能在一定程度上反映根系活力的情況[14]。有研究發現[20-21],作物通過某種機制感知根際養分變化,然后通過啟動體內特定基因的表達和相應的生理生態反應,最終引起根系的形態與生理特性變化,來增加養分的吸收。因此,植物對養分的吸收與產量的形成是根系形態特征與生理特性共同影響的結果。本試驗得出,鹽脅迫導致白蠟樹根系的總吸收面積與活躍吸收面積顯著降低,但接種PGPR能使根系總吸收面積與活躍吸收面積明顯升高。這可能是由于該促生細菌能分泌生長素,明顯增強了白蠟樹的根系活性,同時還能顯著改善土壤微生態環境條件,進而為根系創造了優越的生長環境。
本研究還得出,鹽脅迫明顯抑制了白蠟樹地徑、株高的生長,而接種陰溝腸桿菌TIA062則可顯著促進其生長。這可能與PGPR能顯著提高白蠟樹根際微生物數量和酶活性,并增加白蠟樹根系總吸收面積與活躍吸收面積有密切關聯。這進一步驗證了根際促生細菌可通過改善根系生長的微域環境條件,進而促進苗木的生長。
4 結 論
鹽脅迫顯著降低了白蠟樹根際的微生物數量、微生物量碳、氮含量和脲酶、多酚氧化酶、過氧化氫酶、蔗糖酶活性,且其降幅隨著鹽脅迫梯度的增加而增大;當接種PGPR后,對應的指標均呈上升趨勢,其中輕度鹽脅迫接種PGPR處理的細菌數、放線菌數和微生物總量達最高。同時,白蠟樹的根體積、根系總吸收面積與活躍吸收面積隨著鹽脅迫程度的加劇呈遞減趨勢,但接種PGPR后,對應的根系指標均有增加,其中輕度鹽脅迫接種PGPR處理的活躍吸收面積顯著高于對照15.97%。此外,同對照相比,不同程度鹽脅迫均顯著降低了白蠟樹的地徑、株高,而接種PGPR則顯著提高了地徑、株高,其中輕度鹽脅迫接種PGPR處理的地徑、株高與對照差異不顯著,但顯著高于其他處理。綜上,輕度鹽脅迫下(鹽分含量0.2%)接種陰溝腸桿菌能顯著改善白蠟樹根際生物學特征,并促進白蠟樹生長,其作用效果顯著優于中度、重度鹽脅迫下接種陰溝腸桿菌。
[1] Huo Long, Pang Huancheng, Zhao Yonggan, et al. Buried straw layer plus plastic mulching improves soil organic carbon fractions in an arid saline soil from Northwest China[J]. Soil and Tillage Research, 2017,165:286-293.
[2] 王佳麗,黃賢金,鐘太洋,等.鹽堿地可持續利用研究綜述[J].地理學報,2011,66(5):673-684.
[3] 馬海林,劉方春,馬丙堯,等.白蠟樹容器苗和裸根苗對鹽分脅迫的生長響應[J].東北林業大學學報,2016,44(6):13-16.
[4] 王彎彎,諸葛玉平,王慧橋,等.外源NO對鹽脅迫下小麥幼苗生長及生理特性的影響[J].土壤學報,2017,54(2):516-524.
[5] 劉方春,邢尚軍,馬海林,等.干旱生境中接種根際促生細菌對核桃根際土壤生物學特征的影響[J].應用生態學報,2014,25(5):1475-1482.
[6] 劉方春,馬海林,杜振宇,等.金銀花容器苗對干旱脅迫下接種根際促生細菌的生理響應[J].生態學報,2015,35(21):7003-7010.
[7] 丁新景,黃雅麗,馬風云,等.根際促生菌對景天科多肉植物雪蓮的促生作用[J].東北林業大學學報,2016,44(12):26-30.
[8] 康貽軍,程潔,梅麗娟,等.植物根際促生菌作用機制研究進展[J].應用生態學報,2010,21(1):232-238.
[9] Liu Fangchun, Xing Shangjun, Ma Hailin, et al. Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings[J]. Applied Microbiology and Biotechnology, 2013,97(20):9155-9164.
[10] 井大煒,邢尚軍,劉方春,等.保水劑施用方式對側柏根際微生態環境的影響[J].農業機械學報,2016,47(5):146-154.
[11] 沈仁芳,趙學強.土壤微生物在植物獲得養分中的作用[J].生態學報,2015,35(20):6584-6591.
[12] 井大煒,王明友,張紅,等.蚯蚓糞配施尿素對豇豆根系特征與根際土腐殖質的影響[J].農業機械學報,2017,48(1):212-219.
[13] 關松蔭.土壤酶及其研究方法[M].北京:中國農業出版社,1986:274-332.
[14] 井大煒,邢尚軍,劉方春,等.保水劑—尿素凝膠對側柏裸根苗細根生長和氮素利用率的影響[J].應用生態學報,2016,27(4):1046-1052.
[15] 劉磊,李彩鳳,郭廣昊,等. NaCl+Na2SO4脅迫對甜菜根際土壤微生物數量及酶活性的影響[J].核農學報,2016,30(10):2033-2040.
[16] 莫俊杰,彭詩春,葉昌輝,等.鹽脅迫下甘蔗根際土壤微生物量及其酶活性的效應分析[J].廣東農業科學,2016,43(6):103-108.
[17] 周德平,吳淑杭,褚長彬,等.鹽脅迫對蔬菜地土壤微生物及土壤酶活的毒害效應[J].農業環境科學學報,2011,30(8):1602-1607.
[18] 張雪,賀康寧,史常青,等.鹽脅迫對檉柳和白刺幼苗生長與生理特性的影響[J].西北農林科技大學學報:自然科學版,2017,45(1):105-111.
[19] 吳林坤,林向民,林文雄.根系分泌物介導下植物—土壤—微生物互作關系研究進展與展望[J].植物生態學報,2014,38(3):298-310.
[20] Bankaji I, Cacador I, Sleimi N. Assessing of tolerance to metallic and saline stresses in the halophyteSuaedafruticosa: The indicator role of antioxidative enzymes [J]. Ecological Indicators, 2016,64:297-308.
[21] Chaum S, Kirdmanee C. Effect of inland salt-affected soil on physiological and growth characters of Eucalypt tree (EucalyptuscamaldulensisDehnh.)[J]. Emirates Journal of Food and Agriculture, 2010,22(6):466-474.
