組蛋白H3(Ser10) 磷酸化在維持腫瘤細胞惡性表型中的作用
2018-04-09 07:21:09黎青葉柳曉玲吳小嫩陳麗萍
癌變·畸變·突變 2018年2期
黎青葉,柳曉玲,吳小嫩,郭 萍 ,陳 雯,陳麗萍*
(中山大學公共衛生學院預防醫學系,廣東 廣州 510080)
2014年世界癌癥報告顯示,全球每年癌癥新發病例約1 400萬,死亡約800萬,其中肝癌和肺癌的發病率在全球高居前3位;在肝、食道、胃和肺4種臟器的惡性腫瘤中,中國新增病例和死亡人數均居世界首位[1]。腫瘤的形成和發展是多階段、多因素的過程,其中涉及癌基因的激活和抑癌基因的失活,以及遺傳和表觀遺傳機制的異常[2]。表觀遺傳是指不涉及DNA序列改變,而在基因表達層面上發生了可遺傳的改變,包括DNA甲基化、組蛋白修飾、染色質重塑和非編碼RNA[3]。近來腫瘤形成和發展相關的組蛋白修飾日益受到表觀遺傳學研究學者的重視,且越來越多的結果表明組蛋白修飾在腫瘤發生發展過程中發揮重要的作用[4-5]。
組蛋白H3磷酸化作為有絲分裂的中期相標志,在G1期調控轉錄活性,在G2/M期可影響染色質凝集[6],具有調控基因轉錄、DNA損傷修復、細胞分化和凋亡等功能[7-8]。其中,組蛋白H3絲氨酸10位的磷酸化修飾[H3(Ser10)]與細胞周期及基因轉錄調控等密切相關,以往研究發現組蛋白H3(Ser10)磷酸化作為Aurora B激酶高表達的結果可能參與致癌過程[9];也可通過調節14-3-3ζ蛋白、絲裂素活化蛋白激酶磷酸酶-1(MKP1)、絲裂原和應激激活蛋白激酶1(MSK1)的表達,在染色質結構的調節中發揮作用[10]。多種腫瘤促進因子如表皮生長因子(epidermal growth factor,EGF)誘導組蛋白H3(Ser10)的磷酸化進而激活轉錄因子連接蛋白1(adaptor protein 1,AP-1)[11]、鎘通過下調載脂蛋白E(apolipoprotein E,ApoE)[12]或 改變細胞周期[13]等,在細胞惡性轉化中起重要作用。然而,目前對于組蛋白H3(Ser10)磷酸化參與惡性腫瘤細胞表型維持或腫瘤形成的機制仍不明確。
本實驗室前期研究發現,在化學致癌物誘導轉化細胞模型和腫瘤中,蛋白磷酸酶2A(PP2A)的調控蛋白α4呈高表達[14]。基于以上有關組蛋白H3(Ser10)磷酸化的研究,本文擬探討組蛋白H3(Ser10)磷酸化在維持腫瘤細胞惡性表型中的作用,并通過α4的表達調控來闡明H3(Ser10)磷酸化在其中的可能機制。
1 材料與方法
1.1 主要試劑
DMEM和RPMI-1640基礎培養基、100×青/鏈霉素、胎牛血清均購于美國Gibco公司;胰蛋白酶購于美國Amresco公司。兔抗人組蛋白H3及Ser10磷酸化多克隆抗體購于美國Cell Signaling Technology公司,兔抗人IGBP1多克隆抗體購于美國Novus Biologicals公司,熒光標記抗兔二抗購于美國Thermo Fisher Scientific公司,鼠抗人β-actin單克隆抗體、辣根過氧化物酶標記的山羊抗小鼠及兔IgG(二抗)購于美國Santa Cruz公司。胸苷(Thymidine)、諾卡達唑(Nocodazole)均購自美國Sigma公司;碘化丙啶(PI)、DAPI購于上海碧云天公司;甲醛、乙醇、氯化鎘(CdCl2)均購于廣州化學試劑公司;雙熒光素酶報告檢測試劑盒購于美國Promega公司。
1.2 主要儀器
電泳儀、電泳槽和轉膜儀均購于美國Life公司;倒置顯微鏡購于日本Nikon公司;流式細胞儀購于美國Beckman Coulter公司。
1.3 質粒構建
利用質粒擴增突變試劑盒(Stratagene,美國)構建組蛋白H3(Ser10)位點突變質粒(pBABE-HAH3S10A)。另外以人cDNA為模板,用高保真PCR酶(Thermo Scientific,美國)擴增得到HA-AP1產物后進行凝膠回收,經酶切后裝載至pBABE-puro質粒中,構建AP-1 過表達質粒(pBABE-HA-AP1)。α4啟動子質粒(p G L 3-α 4)人 α 4啟 動 子 片 段 從 h t t p://switchgeargenomics.com獲取,設計引物擴增后酶切插入pGL3-MCS-control質粒(中山大學生命科學院莊詩美教授饋贈),所有質粒經測序確認。
1.4 細胞培養
人肝正常細胞株L02來自深圳疾病預防控制中心,人肝癌細胞株Bel7402、HepG2、SMMC-7721和人肺癌細胞株A549來自廣州市疾病預防控制中心毒理科。L02、Bel7402、HepG2、SMMC-7721和A549細胞培養于RPMI-1640完全培養基(含10%胎牛血清),置于CO2體積分數為5%、37 ℃恒溫箱中培養,待細胞生長至匯合度約為80%時,用0.25%胰蛋白酶消化傳代。
1.5 構建組蛋白H3(Ser10)位點突變和AP-1過表達的細胞株
pBABE-HA-H3S10A或pBABE-HA-AP1質粒、包裝質粒pCL-ampho按1∶1比例經磷酸鈣法轉染293FT細胞,產生的病毒經過濾后感染靶細胞Bel7402和A549,經嘌呤霉素(1 μg/mL)篩選獲得陽性細胞,并經免疫印跡驗證目的基因或標簽蛋白HA的表達。
1.6 流式細胞術檢測細胞周期
將細胞按每孔1×105~5×105個細胞接種到6 cm培養皿,貼壁24 h后,待細胞生長至匯合度70%~80%時,采用胸苷嘧啶核苷雙阻斷法的處理組,在細胞培養基中加入2 mmol/L胸苷處理24 h,使細胞處于G1/S期;采用諾考達唑阻抑法的處理組,用100 ng/mL諾考達唑處理24 h使細胞處于G2/M期;對照組不做任何處理。周期阻滯后,消化收集細胞,離心收集的細胞用預冷PBS洗1次,再重懸于500 μL PBS,輕柔震蕩,并逐滴添加1.5 mL預冷的純乙醇,置-20 ℃過夜(至少12 h);上機前在4 ℃,600 g離心10 min處理樣品,然后用冰PBS洗1次,靜置5 min后再次離心,重懸于300~500 μL的PI/Triton X-100染色液中,37 ℃孵育15 min,后經200目濾網過濾到流式管中,置冰上避光,24 h后用流式細胞儀進行檢測;檢測結果用Flowjo 7.6軟件擬合分析。實驗重復3次。
1.7 免疫熒光檢測磷酸化H3(Ser10)的表達
將L02細胞按每孔1×104個接種于24孔板,培養24 h后用周期藥物阻滯(如1.6中所述)。分別收集對照組、胸苷處理組和諾考達唑處理組的細胞,棄培養基,用預溫的PBS洗3次,然后用3.7%的甲醛(用37 ℃預熱PBS配制)室溫固定15 min;去除固定液,加入PBS 輕柔搖動3 min,重復3次,然后加入含有0.1% Triton X-100的通透液(PBS配制)室溫通透5 min。吸走通透液后再用PBS洗3次,加入封閉液(含3% FBS的PBS),在37℃封閉30 min。封閉結束后,加入用封閉液稀釋好的一抗p-H3(Ser10)(1∶1 000),4 ℃孵育過夜。次日用PBS洗3次,每次5 min,加入熒光標記Rabbit二抗(1∶1 000),37 ℃孵育1 h。二抗孵育結束后,用PBS洗3次,再用DAPI(1∶1 000)襯染細胞核5 min,PBS洗1次,在免疫熒光顯微鏡下觀察,并拍照記錄。實驗重復3次。
1.8 蛋白印跡檢測H3(Ser10)磷酸化水平改變對α4表達的影響
按上述方法用100 ng/mL諾考達唑將人正常肝細胞株L02和肝癌細胞株HepG2、Bel7402、SMMC-7721阻滯在M2期。收集細胞后使用SDS上樣緩沖液直接裂解法獲得細胞總蛋白,并使用細胞破碎儀(功率30%,超聲5 s,停頓1 s)處理樣品,8%~16%梯度膠分離蛋白,濕轉法進行轉膜。5%的脫脂牛奶封閉1 h,然后分別加入兔抗人抗體α4(1∶5 000)、p-H3(Ser10)(1∶1 000)4℃孵育過夜,次日用PBS/T洗3次,每次5 min,再置于含山羊抗兔IgG HRP(1∶10 000),室溫孵育1 h。加上ECL發光劑進入暗室使用X光膠片曝光成像。實驗重復3次。
1.9 軟瓊脂試驗檢測腫瘤細胞克隆形成能力
構建p-H3(Ser10)低表達的Bel-H3S10A(S10A)細胞株,轉染空載質粒的Bel-Vector(AP-1)細胞株為對照。使用DMEM培養基配置底層瓊脂(含0.6%瓊脂、10%胎牛血清、1%青/鏈霉素、1%兩性霉素B)鋪于6孔板冷卻備用。消化細胞并計數,按每孔2×104個細胞的密度混懸于頂層瓊脂(含0.4%瓊脂、10%胎牛血清、1%青/鏈霉素、1%兩性霉素B)中,鋪于底層瓊脂上,冷卻凝固后置于細胞培養箱培養。連續培養21 d后,顯微鏡下觀察克隆并計數。實驗重復3次。
1.1 0 雙熒光素酶報告試驗檢測p-H3(Ser10)對α4的轉錄調控作用
將A549細胞按每孔1×104個的密度接種于96 孔板,貼壁24 h后,按說明書使用Lipofectamine?2000 轉染試劑進行pGL3-α4及對照質粒的轉染,48 h后用20 μmol/L氯化鎘染毒,12 h后使用熒光素酶報告試劑盒上機檢測并記錄數據。每個樣品做3個重復孔,取均值;以質粒pRL-TK海腎光的讀取值為內參,計算得到結果。實驗重復3次。
1.1 1 統計學方法
應用SPSS 21.0及Graph-pad Prism- 5.0統計學軟件處理和分析實驗數據。計量資料采用x±s表示,多組間比較采用方差分析,組內兩兩比較采用LSD-t檢驗,以p<0.05為差異有統計學意義。
2 結 果
2.1 磷酸化修飾的組蛋白H3(Ser10)在細胞中呈周期性表達
組蛋白H3磷酸化修飾具有周期性,且是細胞有絲分裂中期標志[15],因此首先對L02細胞進行了細胞周期阻滯來檢測p-H3(Ser10)的表達。如圖1A所示,L02細胞經2 mmol/L胸苷處理24 h(T)后,大多數細胞處于G1和S期,而經100 ng/mL諾考達唑處理24 h(N)后則處于G2/M期。免疫熒光和免疫印跡結果顯示(1B和1C),與未處理組相比,p-H3(Ser10)表達水平在胸苷處理后明顯下降達71%(p<0.05),幾乎無表達,而在諾考達唑處理后表達明顯增加,為對照組的3.3倍(p<0.05)。表明,p-H3(Ser10)在細胞中呈周期性表達,且在細胞分裂中期達到最高值。
2.2 磷酸化組蛋白H3(Ser10)在腫瘤細胞中的表達及功能
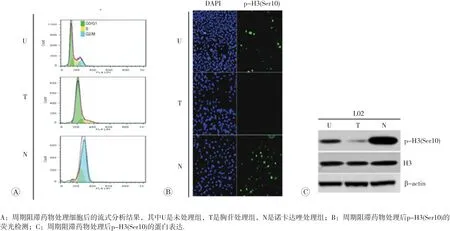
圖1 p-H3(Ser10)的周期性表達
將細胞經諾考達唑阻滯在M2期后,采用免疫印跡法檢測人正常肝細胞株及肝癌細胞株中p-H3(Ser10)的蛋白表達。結果如圖2A所示,與對照L02細胞相比,肝癌細胞株HepG2、Bel7402、SMMC-7721中的p-H3(Ser10)水平升高了(2.61±0.45)倍(p<0.05),但總的組蛋白水平未見明顯改變,提示p-H3(Ser10)高表達在腫瘤發生過程中起作用。為驗證p-H3(Ser10)在腫瘤中的作用,我們進一步通過對Ser10位點進行突變(絲氨酸突變為丙氨酸),構建了p-H3(Ser10)低表達的Bel-H3S10A細胞株,并對該細胞株的惡性轉化功能進行檢測。如圖2B所示,Ser10位點突變引起的p-H3(Ser10)水平下降,使Bel7402腫瘤細胞株在軟瓊脂上形成的克隆數目減少30%(p<0.05),表明p-H3(Ser10)在維持腫瘤細胞惡性表型具有重要作用。
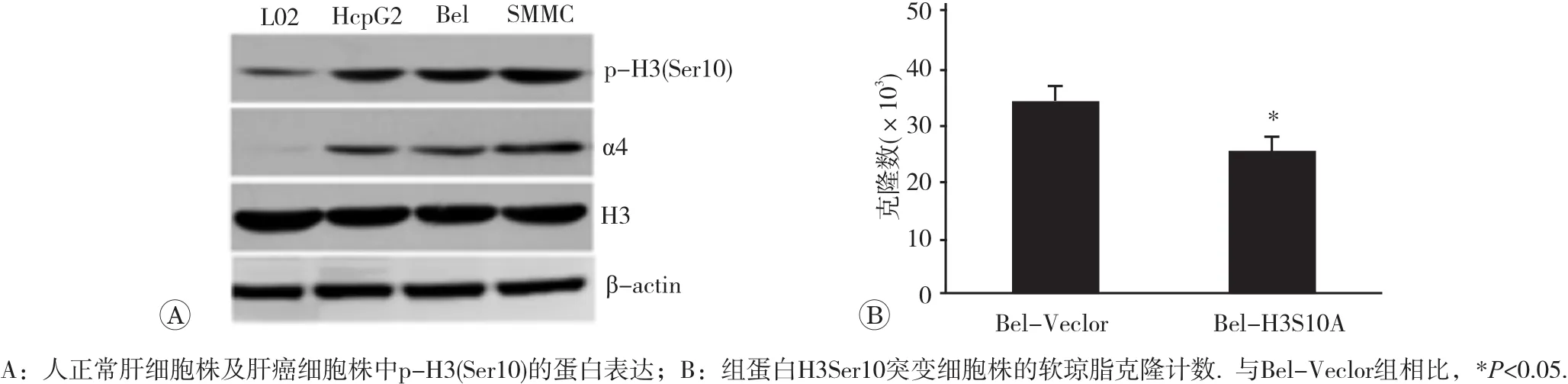
圖2 p-H3(Ser10)在腫瘤細胞中的表達及功能
2.3 鎘誘導組蛋白H3(Ser10)磷酸化修飾增加
進一步探討p-H3S10過表達參與腫瘤發生的可能機制。和前期實驗室結果一致[14],圖2A所示,在癌細胞株中除p-H3(Ser10)高表達外,還觀察到蛋白磷酸酶2A(PP2A)的調控因子α4的高表達,進一步用氯化鎘處理A549細胞。免疫印跡結果如圖3所示,在不同濃度的氯化鎘(0、10、20和40 μmol/L)處理后,隨著氯化鎘濃度的增高,p-H3(Ser10)的表達水平明顯增加,20和40 μmol/L的濃度處理下分別上升了57%和142%(p<0.05),且具有劑量-反應關系,但總的組蛋白表達水平未見明顯改變;同時α4的表達也隨著濃度增高而明顯增加,分別是對照組的1.41、2.80和2.84倍(p<0.05)。在p-H3(Ser10)誘導激活表達的同時也觀察到α4的表達增加,因此我們推測在癌細胞中p-H3(Ser10)可能參與調控α4的表達。
2.4 組蛋白H3(Ser10)磷酸化調控α4的表達

圖3 不同濃度鎘誘導p-H3(Ser10)和α4表達增加
以往研究表明組蛋白磷酸化可激活轉錄因子AP-1對靶基因的轉錄調控作用[16],且通過生物信息學網站預測,α4啟動子區有多個AP-1的結合位點(圖4A)。為驗證p-H3(Ser10)是否通過AP-1調控α4的表達,我們在A549細胞中構建高表達AP-1同時H3(Ser10)位點突變的細胞株A549-AP1-Vector(AP1)和A549-AP1-H3S10A(S10A)。 用20 μmol/L的氯化鎘染毒后,檢測α4的表達水平。如圖4B所示,雙熒光素酶報告試驗結果顯示,AP1細胞經鎘處理后,AP-1的轉錄活性升高了80%(p<0.05);經鎘處理后,p-H3(Ser10)低水平表達的S10A細胞株中AP-1的轉錄活性升高幅度比AP1細胞下降了30%(p<0.05)。一致的是,蛋白印跡結果也顯示(圖4C),在鎘作用下,與高表達AP-1的對照細胞AP1相比,組蛋白H3(Ser10)位點突變株S10A細胞中,p-H3 (Ser10)的表達水平降低51%,同時檢測到S10A細胞中的α4蛋白表達較對照細胞株下降47%(p<0.05),但總的組蛋白表達水平未見明顯改變。以上結果提示p-H3(Ser10)可通過激活轉錄因子AP-1調控α4的表達。
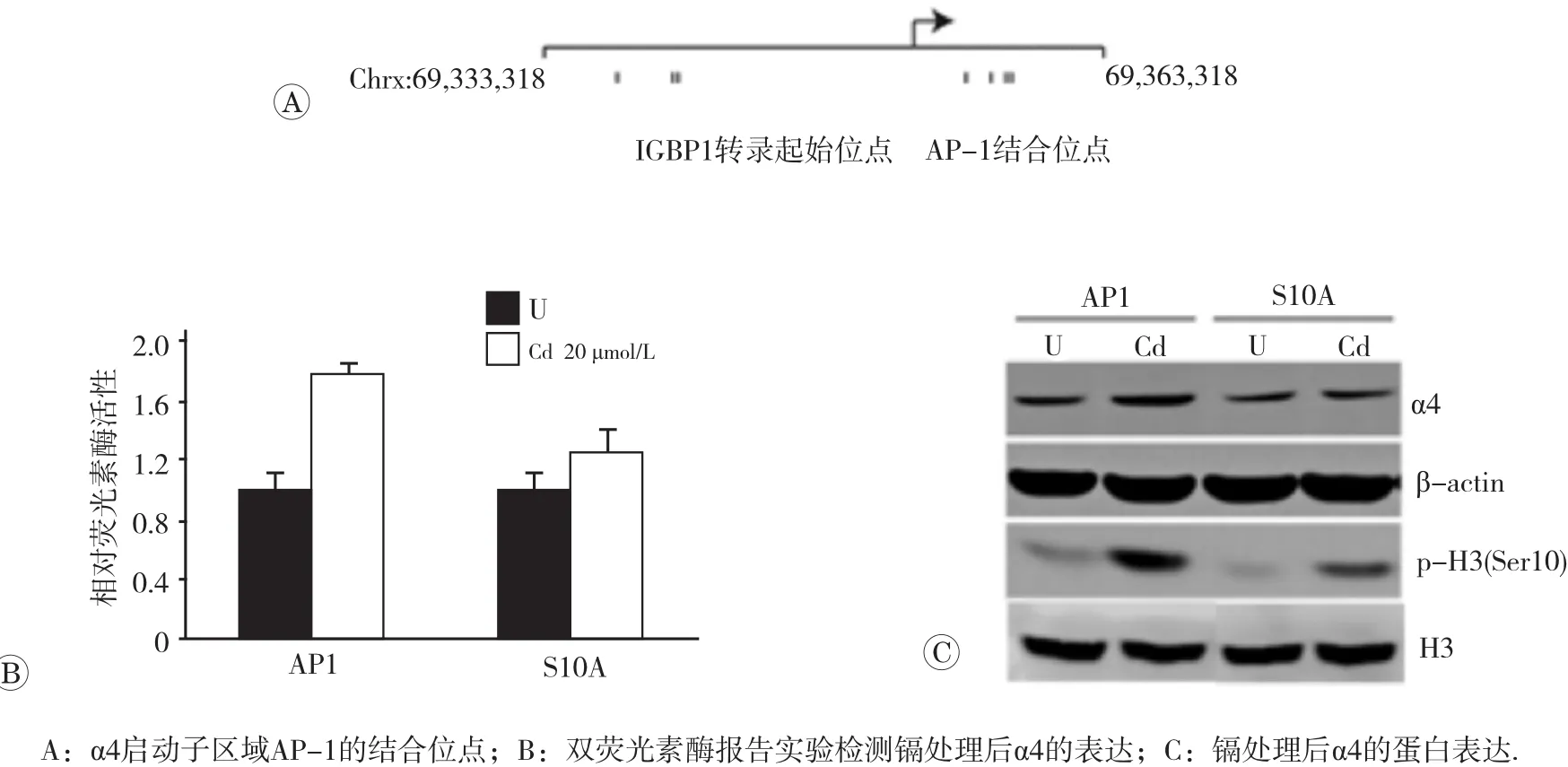
圖4 p-H3(Ser10)調控α4的表達
3 討論
組蛋白修飾在轉錄后水平調節細胞內基因的表達,單個基因位點或全基因組中組蛋白修飾調節模式的紊亂會導致腫瘤的形成[17]。本研究發現H3(Ser10)的磷酸化水平在肝癌細胞株中升高,推測高水平磷酸化修飾的H3(Ser10)具有癌基因功能,而進一步的功能實驗則發現p-H3(Ser10)可通過激活轉錄因子AP-1調控α4的表達來參與腫瘤的發生發展。我們的研究揭示了一條組蛋白異常修飾參與腫瘤發生過程的新信號通路。
以往研究發現H3(Ser10)的磷酸化水平在有絲分裂原和癌基因誘導轉化的小鼠成纖維母細胞中是增加的[18],并參與EB病毒潛在膜蛋白-1(LMP1)誘導的鼻咽癌形成過程[19]。相一致的是,我們也發現全基因組水平的組蛋白H3磷酸化修飾表達升高,這與組蛋白H3(Ser10)位磷酸化水平在以上腫瘤中升高的報道是一致的,證實其很可能參與了腫瘤形成的過程。此外,也有研究證實H3(Ser10)的磷酸化修飾與快速反應基因(immediate-early gene,IE)相關,包括原癌基因c-fos和c-jun[20]。IE基因反應在分化、有絲分裂、疾病如癲癇和癌癥過程中發揮作用[21-22]。
本研究發現H3(Ser10)的磷酸化水平隨著細胞周期呈現規律性變化,這和以往的研究一致[23]。H3(Ser10)的磷酸化水平在癌細胞株中升高,也在可誘導細胞惡性轉化的化學致癌物氯化鎘誘導下升高,這與α4的表達相似且趨勢一致。我們之前發現PP2A的調節蛋白α4在人惡性細胞轉化過程中起著重要的調控作用,α4在轉化細胞中表達顯著上調并且在肺腺癌組織中呈高表達水平。本研究中α4在肝癌細胞株HepG2、Bel7402、SMMC-7721中的表達水平較L02細胞均顯著升高,且氯化鎘染毒處理也可以誘導α4的表達上調,進一步支持了α4在化學致癌和惡性轉化表型中的重要作用。α4基因啟動子存在轉錄因子AP-1的調控區域,我們構建了高表達AP-1同時H3(Ser10)位點突變的細胞株,發現轉錄因子AP-1對α4的表達調控作用減弱,與高表達AP-1的對照細胞相比,H3(Ser10)位點突變細胞的p-H3 (Ser10)和α4表達水平均下降近50%;且在細胞受到致癌化學物氯化鎘的刺激下,低表達p-H3(Ser10)細胞的α4表達升高幅度比對照細胞下降近50%,說明p-H3(Ser10)可通過激活轉錄因子AP-1調控α4的表達,提示組蛋白H3(Ser10)磷酸化參與了化學物致癌過程中α4的調控通路。
綜上,本研究發現組蛋白H3(Ser10)磷酸化修飾在肝腫瘤細胞中呈過表達水平,且在化學致癌物誘導過程中顯著上調,提示組蛋白H3(Ser10)參與腫瘤細胞維持惡性表型的過程,其機制可能是組蛋白H3(Ser10)通過轉錄因子AP-1調控促癌因子α4的表達。因此,我們認為組蛋白H3(Ser10)磷酸化有望作為癌癥治療的一個新靶點。
[1] STEWART B W,WILD C P. World Cancer Report 2014[M].IARC,2014.
[2] JOHNSON C,WARMOES M O,SHEN X,et al. Epigenetics and cancer metabolism[J]. Cancer Lett,2015,2 Sup A(356):309-314.
[3] MOSHE S. Nongenetic inheritance and transgenerational epigenetics[J]. Trends Mol Med,2015,21(2):134-144.
[4] LI D,BI F F,CAO J M,et al. Poly (ADP-ribose) polymerase 1 transcriptional regulation:A novel crosstalk between histone modification H3K9ac and ETS1 motif hypomethylation in BRCA1-mutated ovarian cancer[J]. Oncotarget,2014,1(5):291-297.
[5] SUN Y,BELL J L,CARTER D,et al. WDR5 supports an NMyc transcriptional complex that drives a protumorigenic gene expression signature in neuroblastoma[J]. Cancer Res,2015,75(23):5143-5154.
[6] NOWAK S J,CORCES V G. Phosphorylation of histone H3:a balancing act between chromosome condensation and transcriptional activation[J]. Trends Genet,2004,4(20):214-220.
[7] METZGER E, YIN N A, WISSMANN M, et al.Phosphorylation of histone H3 at threonine 11 establishes a novel chromatin mark for transcriptional regulation[J]. Nat Cell Biol,2007,10(1):53-60.
[8] YOSHIDA I,IBUKI Y. Formaldehyde-induced histone H3 phosphorylation via JNK and the expression of protooncogenes[J]. Mutat Res,2014(770):9-18.
[9] OTA T,SUTO S,KATAYAMA H,et al. Increased mitotic phosphorylation of histone H3 attributable to AIM-1/Aurora-B overexpression contributes to chromosome number instability[J].Cancer Res,2002,62(18):5168-5177.
[10] SHARMA A K,MANSUKHA,VARMA A,et al. Molecular modeling of differentially phosphorylated serine 10 and acetylated lysine 9/14 of histone H3 regulates their interactions with 14-3-3 zeta, MSK1, and MKP1[J]. Bioinform Biol Insights,2013(7):271-288.
[11] CHOI H S,CHOI B Y,CHO Y Y,et al. Phosphorylation of histone H3 at serine 10 is indispensable for neoplastic cell transformation[J]. Cancer Res,2005,65(13):5818-5827.
[12] TAKEDA S M,TERAOKA-NISHITANI N,YAMAGATA A,et al. Cadmium-induced malignant transformation of rat liver cells:Potential key role and regulatory mechanism of altered apolipoprotein E expression in enhanced invasiveness[J].Toxicology,2017,382:16-23.
[13] NGALAME N N,WAALKES M P,TOKAR E J. Silencing KRAS overexpression in cadmium-transformed prostate epithelial cells mitigates malignant phenotype[J]. Chem Res Toxicol,2016,9(29):1458-1467.
[14] CHEN L P,LAI Y D,LI D C,et al. [alpha]4 is highly expressed in carcinogen-transformed human cells and primary human cancers[J]. Oncogene,2011,30(26):2943.
[15] XU Y M,Du J Y,LAU A T. Posttranslational modifications of human histone H3:an update[J]. Proteomics,2014,17/18(14):2047-2060.
[16] SCHMUCKER A C,WRIGHT J B,COLE M D,et al. Distal interleukin-1 beta (IL-1 beta) response element of human matrix metalloproteinase-13 (MMP-13) binds activator protein 1 (AP-1)transcription factors and regulates gene expression[J]. J Biol Chem,2012,2(287):1189-1197.
[17] VARDABASSO C,HASSON D,RATNAKUMAR K,et al.Histone variants:emerging players in cancer biology[J]. Cell Mol Life Sci,2014,3(71):379-404.
[18] STRELKOV I S,DAVIE J R. Ser-10 phosphorylation of histone H3 and immediate early gene expression in oncogenetransformed mouse fibroblasts[J]. Cancer Res,2002,62(1):75-78.
[19] LI B, HUANG G, ZHANG X, et al. Increased phosphorylation of histone H3 at serine 10 is involved in Epstein-Barr virus latent membrane protein-1-induced carcinogenesis of nasopharyngeal carcinoma[J]. BMC Cancer,2013,13:124.
[20] HAZZALIN C A,BARRATT M J,MAHADEVAN L C,et al.The nucleosomal response associated with immediate-early gene induction is mediated via alternative MAP kinase cascades:MSK1 as a potential histone H3/HMG-14 kinase[J]. EMBO J,1999,18(17):4779-4793.
[21] HERSCHMAN H R. Primary response genes induced by growth factors and tumor promoters[J]. Annu Rev Biochem,1991(60):281-319.
[22] MCMAHON S B,MONROE J G. Role of primary response genes in generating cellular responses to growth factors[J].FASEB,1992,6(9):2707.
[23] FEHRI L F,RECHNER C,JANSSEN S,et al. Helicobacter pylori-induced modification of the histone H3 phosphorylation status in gastric epithelial cells reflects its impact on cell cycle regulation[J]. Epigenetics,2009,8(4):577-586.
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
學苑創造·A版(2015年11期)2016-01-14 09:03:27
