大黃魚人工感染殺香魚假單胞菌的病理形態(tài)學及病原分布的初步研究
2018-04-11 09:00:48謝建軍王庚申許文軍
浙江海洋大學學報(自然科學版) 2018年6期
陳 卓,施 慧,謝建軍,汪 瑋,王庚申,何 杰,許文軍
(浙江海洋大學海洋與漁業(yè)研究所,浙江省海洋水產(chǎn)研究所,浙江省海水增養(yǎng)殖重點實驗室,浙江舟山 316021)
殺香魚假單胞菌Pseudomonas plecoglossicida是引起大黃魚Pseudosciaena crocea內(nèi)臟白點病的病原,大黃魚一旦感染發(fā)病,在自然狀態(tài)下死亡率較高,是危害大黃魚養(yǎng)殖業(yè)的重要病原菌。本實驗室在養(yǎng)殖生產(chǎn)中發(fā)現(xiàn)殺香魚假單胞菌還會感染黑鯛Acanthopagrus schlegelii、小黃魚Larimichthys polyactis等其它海水養(yǎng)殖魚類。研究表明殺香魚假單胞菌對多種水產(chǎn)常用抗生素類藥物有抵抗性,通過依賴藥物以控制該菌效果較差,也可能導致水產(chǎn)品中的藥物殘留進而引起食品質(zhì)量安全問題[1-4]。但是,近年隨著養(yǎng)殖環(huán)境的日益惡化,大黃魚內(nèi)臟白點病在各大黃魚養(yǎng)殖區(qū)域廣泛爆發(fā),給大黃魚養(yǎng)殖業(yè)帶來了巨大的經(jīng)濟損失。
目前,已有不少學者對殺香魚假單胞菌的致病機理及相關的毒力因子展開研究,張丹楓等[5]對大黃魚進行了人工感染實驗與超微電鏡的觀察,從病理學角度展示了病原菌對魚內(nèi)臟組織的入侵與危害;張杰等[6]采用雙交換同源重組法構建了殺香魚假單胞菌Ⅲ型分泌系統(tǒng)轉錄調(diào)控蛋白ExsA突變株,從分子生物學角度闡述了該菌的致病機制,但殺香魚假單胞菌的致病機理至今尚不清楚。本文擬通過人工感染試驗分析殺香魚假單胞菌在大黃魚各器官組織中的分布情況及其對組織產(chǎn)生的病理學變化來闡述病理損傷與病原分布的相互關系,為更深入的探討殺香魚假單胞菌致病機理以及疾病的防治提供理論基礎和依據(jù)。
1 材料與方法
1.1 材料
殺香魚假單胞菌ZS161211為本實驗室分離和保存;健康大黃魚體重約40±3 g,購自舟山臺門星鑫水產(chǎn)養(yǎng)殖場,對試驗魚進行細菌分離及寄生蟲觀察確認無病后實驗室暫養(yǎng)1周備用;兔抗殺香魚假單胞菌多克隆抗體由本實驗室制備保存。
硫代硫酸鹽檸檬酸鹽膽鹽蔗糖瓊脂培養(yǎng)基(TCBS)、胰蛋白胨大豆肉湯培養(yǎng)基(TSB),購自杭州微生物試劑有限公司;電泳儀、紫外凝膠成像儀(Bio-Rad);離心機(Eppendorf);恒溫培養(yǎng)箱(上海精宏實業(yè)有限公司);恒溫搖床(太倉市科教器材廠);顯微鏡(Nikon)。
1.2 試驗方法
1.2.1 人工回感試驗
將保存的菌株ZS161211在TSA培養(yǎng)基上劃線,18℃下培養(yǎng)24~48 h,用1.5%生理鹽水從平板上洗下菌苔,分光光度計測定菌液濃度,將菌液稀釋至終濃度為5×104 cfu/mL。試驗魚每尾腹腔注射稀釋的菌液100 μL,使感染菌相對魚體重的終濃度為100 cfu/g,對照組魚腹腔注射1.5%生理鹽水100 μL。試驗期間水溫控制在20℃,不投餌,連續(xù)充氣。注射后24 h開始取樣,取樣時對試驗魚的肝脾腎內(nèi)臟組織分別進行細菌分離,同時取適量組織塊使用Bouin氏液進行固定。
1.2.2 人工感染試驗魚病理組織切片制備
無菌取發(fā)病魚的肝、脾、腎等內(nèi)臟組織,加1.5%無菌生理鹽水剪碎,涂布TCBS培養(yǎng)基,27℃培養(yǎng)24~48 h觀察細菌生長情況;取試驗魚的肝、脾、頭腎、中腎及鰓絲分別約4 mm×4 mm×5 mm,經(jīng)Bouin氏液固定,在24 h后換數(shù)次75%乙醇沖洗,最后保存于75%的乙醇中。固定的樣品經(jīng)后續(xù)不同濃度乙醇系列脫水及丙酮脫水過程后,移入二甲苯溶液中透明,經(jīng)浸蠟包埋后切片、貼片,烤干后展片以蘇木精-伊紅染色法(H&E染色)染色,樹膠封片,顯微鏡下觀察拍照。
1.2.3 免疫組化檢測殺香魚假單胞菌在組織內(nèi)分布
將已制作好的石蠟切片脫蠟水化,滴加3%H2O2于載玻片上進行內(nèi)源性酶滅活,然后將切片浸入枸櫞酸鹽緩沖液中,直接煮至92~98℃進行抗原暴露,冷卻后滴加5%BSA封閉,滴加兔抗殺香魚假單胞菌抗?jié)窈蟹跤渭由锼鼗窖蚩雇枚節(jié)窈蟹跤螅尤肜备^氧化物酶進行SP反應,滴加新鮮配制的試劑DAB顯色,清洗后蘇木素復染,最后滴加中性樹脂膠封片。用PBS代替一抗作為陰性對照,顯微鏡下觀察拍照,顯微鏡下中呈棕褐色反應產(chǎn)物代表抗原的定位,未有抗原定位的則被蘇木精染色成藍色。
2 結果
2.1 臨床癥狀與剖解變化
試驗組魚注射24 h后出現(xiàn)游動緩慢,應激反應遲鈍等臨床癥狀;注射72 h后試驗魚出現(xiàn)死亡,死亡魚體表無潰瘍等癥狀,主要是鰓充血。剖解病死魚,部分魚有腹水,肝充血嚴重,脾臟布滿白色小結節(jié)和腎臟白色結節(jié)較脾臟大(圖1)。無菌取病死魚的內(nèi)臟組織分別在TCBS和TSA培養(yǎng)基上接種劃線,27℃培養(yǎng)24 h,TCBS上長出特征性的殺香魚假單胞菌針尖大小的綠色小菌落,TSA上為乳白色、邊緣光滑、表面濕潤的菌落。經(jīng)藥敏試驗和生理生化學試驗確認與感染菌株ZS161211一致。
臨床解剖觀察(圖2)顯示試驗組感染魚的肝、脾、腎等內(nèi)臟組織與器官都表現(xiàn)出不同程度的充血、出血、壞死等。肝細胞發(fā)生廣泛性變性,或壞死、溶解,有大量的炎性細胞浸潤;腎小管上皮細胞腫脹變性,腎間質(zhì)巨噬細胞和中性粒細胞浸潤、嚴重空泡變性、壞死崩解;脾臟少量含鐵血黃素的沉積,淋巴細胞壞死,中性粒細胞和大量巨噬細胞浸潤。

圖1 病死魚臨床解剖圖Fig.1 Clinical anatomic map of P.crocea
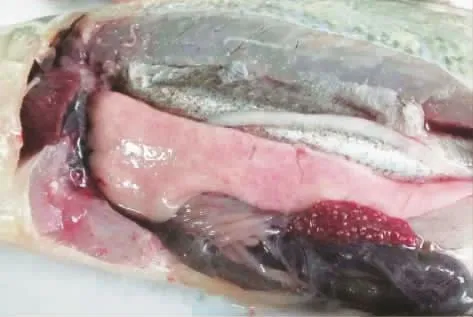
圖2 病魚脾臟充滿白色結節(jié)Fig.2 P.crocea spleen full of the white nodulars
2.2 組織病理學變化
健康大黃魚肝組織結構清晰,細胞形狀規(guī)則;感染魚肝臟組織結構不明晰且伴隨充血,肝血竇與中央靜脈瘀血更明顯,肝血竇、中央靜脈以及一些肝細胞內(nèi)發(fā)現(xiàn)病原菌;肝細胞腫脹明顯、細胞核固縮,病變嚴重的區(qū)域失去原有形態(tài)結構形成壞死灶,壞死灶中心區(qū)內(nèi)充滿解體后的細胞碎片(圖3)。
健康脾臟結構清晰,細胞完整;感染魚脾臟組織壞死嚴重,實質(zhì)結構遭到破壞,在紅髓和白髓均可見散在的壞死灶,壞死灶內(nèi)大量中性粒細胞和巨噬細胞浸潤,脾髓出血較為嚴重,有少量含鐵血黃素的沉積,壞死灶內(nèi)實質(zhì)細胞界限模糊不清,崩解后呈現(xiàn)空泡化。
健康腎臟組織致密;感染魚腎臟充血,結構紊亂;腎小管形狀不規(guī)則,上皮細胞肥大、顆粒變性且界限模糊不清晰、管腔狹窄,部分上皮細胞增生堵塞官腔;腎小球萎縮或崩解;嚴重的腎間質(zhì)組織廣泛性壞死,大量巨噬細胞和中性粒細胞浸潤,間質(zhì)細胞發(fā)生玻璃樣變性,嚴重的壞死,甚至溶解、消失。
健康鰓絲組織緊密,有規(guī)律性;感染魚鰓絲排列不整齊,有的呈現(xiàn)波紋狀扭曲;鰓小片腫大,竇狀隙擴張充血、滲出;鰓小片上皮細胞肥大、變圓,嚴重的上皮細胞崩解壞死脫落。
2.3 菌體在魚體內(nèi)的分布
陽性信號主要分布于肝竇、巨噬細胞、腎小管上皮細胞、腎間質(zhì)細胞和巨噬細胞內(nèi)。肝組織強陽性信號主要集中于肝竇和靜脈,吞噬細胞破裂后部分周邊區(qū)域、肝竇周圍少量肝細胞和竇壁細胞有明顯壞死性病變;脾組織中的陽性信號位于脾臟間質(zhì)細胞和巨噬細胞的白髓區(qū)。這些迅速增殖的感染性病原體在器官和組織中引起明顯的病理變化。腎組織中的陽性信號主要位于腎小管上皮細胞和腎小球淋巴細胞。由于在腎間質(zhì)中大量的細菌復制,組織發(fā)生壞死和崩解,并且也可在崩解組織中檢測到。鰓組織中的陽性信號集中在小的上皮細胞和基底毛細血管中(圖4)。
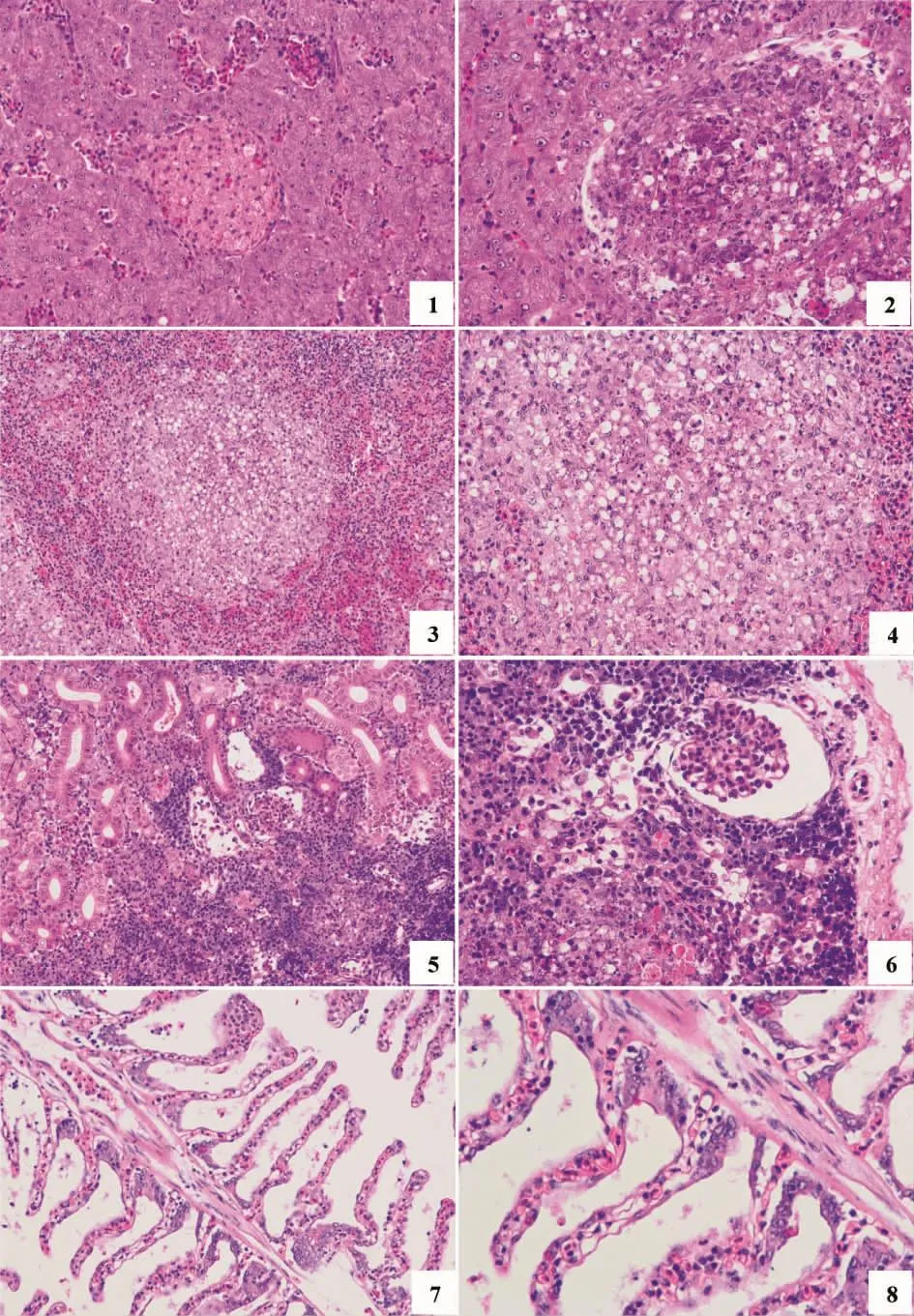
圖3 人工回感魚組織病理變化圖,HEFig.3 Histopathological lesions on P.crocea infected by P.plecoglossicida

圖4 人工回感魚組織免疫組化圖Fig.4 Immunohistochemical staining results of P.crocea infected by P.plecoglossicida
3 討論
本研究在水溫20℃條件下,對養(yǎng)殖大黃魚進行了殺香魚假單胞菌人工感染實驗,運用組織病理與免疫組化技術對發(fā)病魚內(nèi)臟組織進行分析,結果顯示殺香魚假單胞菌對大黃魚組織機體,尤其是內(nèi)臟組織造成了嚴重損傷。
大黃魚內(nèi)臟白點病是近年來在大黃魚養(yǎng)殖中暴發(fā)流行的一種細菌性疾病,因病魚脾、肝臟、腎等內(nèi)臟長滿白色結節(jié)而得名,假單胞菌屬的細菌被確認為該病病原,目前有許多關于大黃魚內(nèi)臟白點病病原的報道[7-9]。由于大黃魚內(nèi)臟白點病臨床發(fā)病急、體表無明顯臨床癥狀、低水溫發(fā)病等特點給該病的防治造成很大困難。關于該病病原假單胞菌種的鑒定一直未能統(tǒng)一,直到2013年張杰等[6]通過全基因組測序確認其為殺香魚假單胞菌。殺香魚假單胞菌由日本學者NISHIMORI,et al[10]從淡水養(yǎng)殖的患病香魚體內(nèi)首次分離得到并且命名。殺香魚假單胞菌不僅存在于淡水環(huán)境中,也廣泛存在于海水環(huán)境中,而且是危害較大的致病菌[4]。
本文采用殺香魚假單胞菌單位為100 cfu/g,注射至大黃魚腹腔內(nèi),感染24 h即開始發(fā)病,肝、脾、中腎、頭腎上出現(xiàn)明顯的白色結節(jié),與自然發(fā)病癥狀相同,且從感染魚內(nèi)臟組織中分離培養(yǎng)到了殺香魚假單胞菌。組織病理觀察結果顯示,感染魚肝、脾、腎等內(nèi)臟組織都出現(xiàn)明顯病變損傷,全身組織器官廣泛性表現(xiàn)出不同程度的充血、出血、壞死等,隨著菌體在大黃魚體內(nèi)大量增殖,最終導致其全身多組織細胞壞死溶解,魚因多組織器官的功能衰竭而死亡,這些組織病理變化與張丹楓等[5]對自然患病魚的組織病理觀察結果一致。
免疫組化技術利用標記的抗體對細胞或組織內(nèi)的相應抗原進行定位檢測,經(jīng)過組織化學的顯色反應呈現(xiàn)醒目的陽性色彩,因此免疫組化可清楚直觀的反映病原感染生物體后的分布狀況[11-12]。隨著現(xiàn)代魚類病害學、分子生物學的迅速發(fā)展,使病理形態(tài)和功能的研究更加深入,免疫組化已成為在水產(chǎn)病害學科研究中的新型手段。連林坤[13]運用免疫組化方法開展了螺原體對中華絨螯蟹Eriocheir sinensis H.Milne-Edwards和克氏原螯蝦Procambarus clarkii不同器官和組織的感染病理學研究,證實了不同水生動物上分離到的螺原體對不同寄主存在感染差異。本試驗應用免疫組化法對人工感染大黃魚各組織器官內(nèi)殺香魚假單胞菌進行定位檢測,在試驗組魚的肝、脾、腎和鰓絲各相關組織器官中觀察到了特征性的陽性信號,這些陽性信號主要分布于肝血竇、脾間質(zhì)或巨噬細胞、腎間質(zhì)活巨噬細胞以及腮絲的巨噬細胞內(nèi),表明殺香魚假單胞菌對機體多組織、器官都具有侵染性。試驗結果表明免疫組化能成功反映殺香魚假單胞菌感染大黃魚后在機體內(nèi)的分布情況。通過組織病理學觀察與免疫組化染色的結果相比較,魚體組織病理損傷程度與免疫組化檢測的中陽性信號的有無與強弱表現(xiàn)出較高一致性,其病理損傷與病原菌分布數(shù)量呈明顯的正相關。
魚類的巨噬細胞是魚類的非特異性免疫系統(tǒng)的重要組成部分,當病原微生物表面覆蓋有免疫球蛋白和補體成分時,巨噬細胞可通過這些因子的特異性受體識別并殺傷微生物[14]。但一些病原菌可以適應巨噬細胞內(nèi)的生活環(huán)境,并作為其生活史的一部分,使其自身免受其它細胞介導和體液免疫反應的傷害,巨噬細胞內(nèi)存活能力已成為很多病原菌毒力機制的重要組成部分[15-16]。張杰等[17]通過電鏡超薄切片觀察發(fā)現(xiàn)殺香魚假單胞菌也可以在巨噬細胞內(nèi)存活和增殖,本試驗的免疫組化結果顯示在試驗魚的多個組織器官的巨噬細胞內(nèi)出現(xiàn)了強陽性信號,為以后進一步進行殺香魚假單胞菌對大黃魚的致病機理研究提供了更多參考資料。
