乙醇對(duì)法夫酵母發(fā)酵合成蝦青素的影響
2018-04-12 09:04:47劉春利沈?qū)幯?/span>倪輝李利君陳艷紅肖安風(fēng)
食品與發(fā)酵工業(yè) 2018年3期
關(guān)鍵詞:質(zhì)量
劉春利,沈?qū)幯啵咻x, 2, 3, 4,李利君, 2, 3, 4,陳艷紅, 2, 3, 4,肖安風(fēng), 2, 3, 4*
1(集美大學(xué) 食品與生物工程學(xué)院,福建 廈門(mén),361021)2(福建省食品微生物與酶工程重點(diǎn)實(shí)驗(yàn)室,福建 廈門(mén),361021)3(福建省海洋功能食品工程技術(shù)研究中心,福建 廈門(mén),361021)4(廈門(mén)市海洋功能食品重點(diǎn)實(shí)驗(yàn)室,福建 廈門(mén),361021)
法夫酵母(Phaffiarhodozyma)中主要的類胡蘿卜素是胞內(nèi)蝦青素,含量可達(dá)到40%~95%,且法夫酵母具有生長(zhǎng)周期短,培養(yǎng)條件簡(jiǎn)單,能代謝多種碳源等優(yōu)勢(shì),成為了天然蝦青素生產(chǎn)的主要研究對(duì)象[14,16]。法夫酵母可以利用乙醇作為碳源進(jìn)行生長(zhǎng)。YAMANE等[17]在細(xì)胞生長(zhǎng)的穩(wěn)定期添加0.3%的乙醇。GU等[18]在延遲期后期或?qū)?shù)生長(zhǎng)期添加0.2%的乙醇,均使色素產(chǎn)量提高了近1倍左右。汪洪濤等[19]針對(duì)不同的法夫酵母菌株,在培養(yǎng)基中添加乙醇,均能提高法夫酵母中蝦青素的含量。
在搖瓶條件下,易于考察各種營(yíng)養(yǎng)因子對(duì)發(fā)酵過(guò)程的影響,從而篩選出適宜蝦青素生產(chǎn)的發(fā)酵條件。本試驗(yàn)對(duì)法夫酵母JMU-MVP14進(jìn)行了搖瓶培養(yǎng),對(duì)比研究不同初始糖濃度與乙醇添加濃度對(duì)法夫酵母細(xì)胞生長(zhǎng)及蝦青素合成的影響,篩選出最利于蝦青素合成的乙醇濃度;然后,利用乙醇傳感器控制整個(gè)發(fā)酵過(guò)程維持在不同恒定乙醇濃度下,考察乙醇濃度對(duì)蝦青素合成的影響;并在此基礎(chǔ)上同時(shí)補(bǔ)加乙醇與葡萄糖,以蝦青素產(chǎn)量為指標(biāo),得到最佳的乙醇補(bǔ)料方式。
1 材料與方法
1.1 材料與試劑
1.1.1菌種
法夫酵母PhaffiarhodozymaJMU-MVP14 菌株:用甲基磺酸乙酯對(duì)法夫酵母JMU-VDL668 菌株誘變,經(jīng)0.5%的雙氧水和紫外照射來(lái)淘汰低產(chǎn)的菌株選育得到[20]。
1.1.2培養(yǎng)基
種子培養(yǎng)基:4°Bx 麥汁,pH 6.0。
無(wú)碳源培養(yǎng)基(g/L):取KH2PO42 g,(NH4)2SO45 g,CaCl20.2 g,MgSO40.5 g,酵母膏3 g,溶于1 L蒸餾水中,調(diào)pH為6.0。
法夫酵母搖瓶發(fā)酵培養(yǎng)基中葡萄糖及乙醇的添加量如表1所示,其他成分組成和無(wú)碳源培養(yǎng)基相同。

表1 培養(yǎng)基中葡萄糖及乙醇的添加量Table 1 Glucose concentration and ethanol additionwithin medium
1.1.3主要試劑
蝦青素(色譜純),Sigma公司;無(wú)水乙醇(分析純),廣東化學(xué)試劑;3,5-二硝基水鹽酸,國(guó)藥集團(tuán);酵母膏(優(yōu)級(jí)純),廣東環(huán)凱;其他試劑均為分析純,購(gòu)于西隴化工。
1.2 實(shí)驗(yàn)儀器和設(shè)備
ZHWY-2102雙層全溫培養(yǎng)搖床,上海智誠(chéng)分析儀器制造有限公司;1260高效液相色譜儀,安捷倫(中國(guó))科技有限公司;SW-CJ-ZFD單面凈化工作臺(tái),蘇州凈化設(shè)備有限公司;S20K pH酸度計(jì),梅特勒-托利多儀器有限公司;BS223S電子天平,賽多利斯科學(xué)儀器(北京)有限公司;101-3B電熱鼓風(fēng)干熱箱,上海市實(shí)驗(yàn)儀器總廠;TD5M-WS臺(tái)式離心機(jī),湖南湘儀實(shí)驗(yàn)室儀器開(kāi)發(fā)有限公司;LDZX-40KAS立式高壓蒸汽滅菌鍋,上海申安醫(yī)療器械廠;2600A紫外可見(jiàn)分光光度計(jì),尤尼柯(上海)儀器有限公司;乙醇感受器,參考相關(guān)專利[21],在某些結(jié)構(gòu)上進(jìn)行改良,自制了乙醇傳感器。
1.3 實(shí)驗(yàn)方法
1.3.1培養(yǎng)方法
將PDA斜面活化的菌種接入種子培養(yǎng)基中,22 ℃培養(yǎng)2~3 d得1代種子,轉(zhuǎn)接1代種子到新配制的種子培養(yǎng)基為2代種子,轉(zhuǎn)接2代種子到搖瓶中,于22 ℃連續(xù)培養(yǎng)120 h,每24 h取樣檢測(cè)生物量、蝦青素產(chǎn)量及殘?zhí)呛浚靡掖紓鞲衅鳈z測(cè)乙醇質(zhì)量濃度。
1.3.2生物量的測(cè)定

1.3.3蝦青素含量的測(cè)定
二甲基亞砜(DMSO)法破壁[23-24]:取5 mL發(fā)酵液離心2次后去上清液得到菌體。將菌體于55 ℃下預(yù)熱5 min后,加入2 mL已預(yù)熱至75 ℃的二甲基亞砜,充分振蕩后加入5 mL乙醇提取色素,離心得到色素提取液,乙醇定容至10 mL。
紫外分光光度計(jì)法檢測(cè)[25]:配制不同濃度梯度的蝦青素標(biāo)準(zhǔn)品,在紫外光波長(zhǎng)為474 nm下檢測(cè),繪制出蝦青素標(biāo)準(zhǔn)曲線。相同條件下,測(cè)定發(fā)酵樣品吸光度,根據(jù)標(biāo)準(zhǔn)曲線求樣品中的蝦青素含量。
1.3.4發(fā)酵液中殘?zhí)堑臏y(cè)定
3,5-二硝基水楊酸法(DNS)[26]:1 mL的發(fā)酵上清液稀釋至適宜濃度,加入0.6 mL的DNS,沸水浴10 min后立即冷卻,用蒸餾水定容至5 mL,使用紫外分光光度計(jì)在波長(zhǎng)為520 nm下測(cè)定。
1.3.5發(fā)酵液中乙醇質(zhì)量濃度的測(cè)定
將配制好的2 g/L的乙醇標(biāo)準(zhǔn)溶液,稀釋后分裝于相同規(guī)格的50 mL離心管中,每管5 mL,用于測(cè)定乙醇標(biāo)準(zhǔn)曲線。于室溫22 ℃下,乙醇傳感器1提前預(yù)熱30 min,待讀數(shù)為0,將傳感器感應(yīng)端置于離心管樣品液面上0.5 cm處,秒表計(jì)時(shí)50 s后讀數(shù)。取出傳感器感應(yīng)端置于空氣中,待讀數(shù)為0后測(cè)定下一個(gè)樣品。
1.3.6二次參數(shù)的計(jì)算

2 結(jié)果與討論
2.1 不同初始糖濃度時(shí)添加乙醇對(duì)發(fā)酵的影響
分別在10 g/L和20 g/L兩種葡萄糖濃度發(fā)酵培養(yǎng)基中添加不同濃度乙醇,考察乙醇對(duì)搖瓶培養(yǎng)法夫酵母JMU-MVP14的細(xì)胞生長(zhǎng)和蝦青素合成的影響。乙醇會(huì)對(duì)法夫酵母和蝦青素合成產(chǎn)生影響。
2.1.1初始葡萄糖質(zhì)量濃度為10 g/L時(shí)添加乙醇對(duì)發(fā)酵的影響
以無(wú)碳源培養(yǎng)基為基礎(chǔ),添加10 g/L的葡萄糖,乙醇的添加濃度如表1所示。考察在較低糖濃度下,不同乙醇濃度對(duì)法夫酵母整個(gè)發(fā)酵過(guò)程中葡萄糖的消耗、細(xì)胞生長(zhǎng)、蝦青素的積累及乙醇濃度的變化情況如圖1所示。
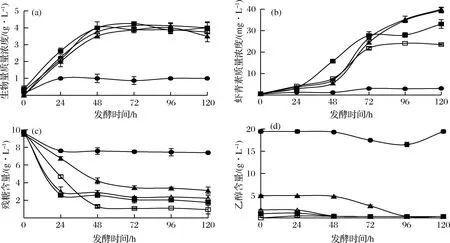
□-不添加乙醇;■-添加1 g/L乙醇;△-添加2 g/L;▲-添加5 g/L乙醇;●-添加20 g/L乙醇圖1 初始葡萄糖質(zhì)量濃度為10 g/L時(shí)添加乙醇對(duì)法夫酵母JMU-MVP14發(fā)酵的影響Fig. 1 The effect of ethanol addtion on astaxanthin production by P. rhodozyma JMU-MVP14 at the initial concentration of10 g/L glucose
由圖1-a可知,添加0~5 g/L的乙醇培養(yǎng)條件下,生物量曲線變化趨勢(shì)基本一致,發(fā)酵至48 h達(dá)到穩(wěn)定期,48 h以后基本不變,且與對(duì)照組(不添加乙醇)相比,添加0~5 g/L的乙醇對(duì)法夫酵母JMU-MVP14生物量影響差別不顯著。從圖1-b可看出,對(duì)照組蝦青素產(chǎn)量在72 h達(dá)到最大,72 h后蝦青素產(chǎn)量保持穩(wěn)定,但添加乙醇后,法夫酵母在72 h后蝦青素產(chǎn)量進(jìn)一步提高。當(dāng)添加乙醇的質(zhì)量濃度為2 g/L和5 g/L時(shí),蝦青素質(zhì)量濃度均達(dá)到最高水平40.0 mg/L,比對(duì)照組顯著提高了70%,且此時(shí)蝦青素細(xì)胞產(chǎn)率分別達(dá)到了10.0 mg/g和11.3 mg/g,與陳鋒等[27]的研究結(jié)果一致。
結(jié)合圖1-c和圖1-d,與對(duì)照組相比,添加1~2 g/L的乙醇時(shí),酵母消耗葡萄糖的速度加快,且發(fā)酵至24 h,酵母不再消耗葡萄糖,開(kāi)始使用乙醇,葡萄糖最終利用率均能達(dá)到78.0%,乙醇最終利用率達(dá)到80.0%;而添加5 g/L的乙醇,酵母發(fā)酵48 h后,才開(kāi)始利用乙醇,乙醇最終利用率為93.4%,葡萄糖最終利用率只有68.2%。由此推測(cè),在葡萄糖和乙醇共同存在的情況下,法夫酵母優(yōu)先利用葡萄糖生長(zhǎng),再利用乙醇,且乙醇能替代葡萄糖作為碳源使酵母產(chǎn)蝦青素,與HU等[28]的文獻(xiàn)報(bào)道一致。而當(dāng)培養(yǎng)基中乙醇的質(zhì)量濃度增至20 g/L時(shí),法夫酵母生長(zhǎng)至24 h就停止生長(zhǎng),對(duì)葡萄糖的利用率降低,發(fā)酵結(jié)束后,生物量質(zhì)量濃度和蝦青素質(zhì)量濃度同對(duì)照組相比差異顯著,分別降低至對(duì)照組的27%和14%。說(shuō)明添加低濃度的乙醇可以促進(jìn)法夫酵母產(chǎn)蝦青素,而添加較高濃度乙醇抑制酵母細(xì)胞的生長(zhǎng)和蝦青素的積累。
2.1.2初始葡萄糖質(zhì)量濃度為20 g/L時(shí)添加乙醇對(duì)發(fā)酵的影響
在較低糖濃度時(shí),當(dāng)添加2 g/L和5 g/L乙醇時(shí),蝦青素產(chǎn)量均達(dá)到最高,且與對(duì)照組相比,添加1~2 g/L質(zhì)量濃度的乙醇,酵母消耗葡萄糖量的速度加快,因此將培養(yǎng)基中初始葡萄糖質(zhì)量濃度提高為20 g/L,其他培養(yǎng)條件不變時(shí),進(jìn)一步考察在較高糖濃度下添加乙醇對(duì)法夫酵母JUM-MVP14發(fā)酵過(guò)程中細(xì)胞生長(zhǎng)、蝦青素合成、乙醇利用等方面的影響。結(jié)果如圖2所示。
如圖2-a所示,添加1~2 g/L的乙醇促進(jìn)了細(xì)胞生長(zhǎng)和蝦青素的積累。且添加1 g/L的乙醇使得生物量較對(duì)照組提高了18%。從蝦青素質(zhì)量濃度曲線圖2-b可看出,添加1 g/L的乙醇,發(fā)酵至120 h,蝦青素質(zhì)量濃度為42.0 mg/L,比對(duì)照組顯著提高了26.9%,此時(shí)乙醇的最終利用率為84.0%,蝦青素細(xì)胞產(chǎn)率為10.5 mg/g,比對(duì)照組提高了7.1%。本試驗(yàn)結(jié)果與肖安風(fēng)等[29]的研究發(fā)現(xiàn)一致,乙醇作為糖代謝途徑中的相關(guān)產(chǎn)物,可為蝦青素合成提供更多的乙酰輔酶A,從而提高了蝦青素的產(chǎn)量。

□-不添加乙醇;■-添加1 g/L乙醇;△-添加2 g/L;▲-添加5 g/L乙醇;○-添加10 g/L乙醇;●-添加20 g/L乙醇圖2 初始葡萄糖質(zhì)量濃度為20 g/L時(shí)添加乙醇對(duì)法夫酵母JMU-MVP14發(fā)酵的影響Fig. 2 The effect of ethanol addtion on astaxanthin production by P. rhodozyma JMU-MVP14 at the initial concentration of20 g/L glucose
從殘?zhí)呛康淖兓闆r發(fā)現(xiàn),在發(fā)酵進(jìn)入對(duì)數(shù)生長(zhǎng)期后,細(xì)胞代謝旺盛,對(duì)碳源的需求增大,從而殘?zhí)呛垦杆傧陆担?2 h之后,菌體生長(zhǎng)趨于穩(wěn)定,糖含量下降速度明顯減緩。其中,添加1 g/L乙醇顯著提高了法夫酵母JUM-MVP14對(duì)葡萄糖的利用率,可達(dá)75.0%(比對(duì)照組顯著提高了36.4%),加快了其對(duì)葡萄糖的代謝,產(chǎn)生更多的乙酰輔酶A[29],為蝦青素的合成提供原料,從而進(jìn)一步闡明低濃度乙醇能促進(jìn)法夫酵母蝦青素合成。但和初始葡萄糖濃度為10 g/L時(shí)相比,酵母對(duì)葡萄糖的利用率明顯降低,且蝦青素的產(chǎn)量卻無(wú)顯著提高。
2.2 控制恒定乙醇的濃度及補(bǔ)加葡萄糖對(duì)發(fā)酵的影響
由2.1的試驗(yàn)結(jié)果可知,乙醇能顯著促進(jìn)法夫酵母JMU-MVP14菌株細(xì)胞生長(zhǎng)及蝦青素的合成,為進(jìn)一步研究乙醇在整個(gè)法夫酵母發(fā)酵周期中對(duì)蝦青素合成的影響,采用乙醇傳感器實(shí)時(shí)在線檢測(cè)乙醇濃度,發(fā)酵過(guò)程補(bǔ)加乙醇,使整個(gè)發(fā)酵周期中乙醇能維持在一個(gè)恒定的濃度。
2.2.1控制恒定乙醇的濃度對(duì)發(fā)酵的影響
在較高葡萄糖濃度環(huán)境中,法夫酵母對(duì)糖的利用率降低,且蝦青素的產(chǎn)量沒(méi)有明顯提高。因此用初始葡萄糖質(zhì)量濃度為10 g/L的培養(yǎng)基,空白組設(shè)為只添加2 g/L的乙醇,實(shí)驗(yàn)組用乙醇傳感器檢測(cè)乙醇濃度在整個(gè)發(fā)酵過(guò)程中維持2 g/L。考察恒定乙醇補(bǔ)料策略對(duì)法夫酵母整個(gè)發(fā)酵過(guò)程中生物量、殘?zhí)恰⑽r青素產(chǎn)量等發(fā)酵參數(shù)的變化情況。實(shí)驗(yàn)結(jié)果如圖3所示。
由圖3-a可知,乙醇持續(xù)補(bǔ)加,JMU-MVP14菌株的細(xì)胞生長(zhǎng)至后期,其生物量提高了,說(shuō)明此菌株可利用乙醇來(lái)合成自身的物質(zhì),即達(dá)到穩(wěn)定期后仍可利用乙醇繼續(xù)生長(zhǎng),與HU等[28]文獻(xiàn)報(bào)道一致。在酵母培養(yǎng)過(guò)程中控制恒定乙醇濃度,從24 h開(kāi)始蝦青素合成量呈對(duì)數(shù)增長(zhǎng)趨勢(shì),發(fā)酵至120 h蝦青素質(zhì)量濃度為50.1 mg/L,比對(duì)照組提高了20.3%。從圖3-c可知,實(shí)驗(yàn)組和對(duì)照組葡萄糖的利用率均能達(dá)到77.2%以上。結(jié)合圖3-d可以看出,乙醇提高了單位細(xì)胞內(nèi)蝦青素的產(chǎn)量(13.1 mg/g),比對(duì)照組(12.2 mg/g)提高了7.4%,進(jìn)而從總體上提高了JMU-MVP14菌株的蝦青素合成能力。說(shuō)明恒定乙醇控制策略更有利于法夫酵母蝦青素的合成。
2.2.2控制恒定乙醇濃度并補(bǔ)加葡萄糖對(duì)JMU-MVP14發(fā)酵的影響
前期的研究結(jié)果表明,在較低糖質(zhì)量濃度(10 g/L)培養(yǎng)基中,添加乙醇,蝦青素質(zhì)量濃度達(dá)到最高水平40.0 mg/L,在高糖質(zhì)量濃度(20 g/L)培養(yǎng)基中,添加乙醇,蝦青素質(zhì)量濃度為42.0 mg/L,后者比前者多了5%,說(shuō)明乙醇對(duì)法夫酵母產(chǎn)蝦青素的作用與葡萄糖濃度密切相關(guān)。故以無(wú)碳源培養(yǎng)基為基礎(chǔ),添加10 g/L的葡萄糖,設(shè)置3個(gè)實(shí)驗(yàn)組,即只補(bǔ)加葡萄糖、只補(bǔ)加乙醇、同時(shí)補(bǔ)加葡萄糖和乙醇,空白對(duì)照只添加2 g/L的乙醇,同時(shí)不補(bǔ)加乙醇也不補(bǔ)加葡萄糖,考察乙醇及葡萄糖的搖瓶補(bǔ)料策略對(duì)法夫酵母整個(gè)發(fā)酵過(guò)程中生物量、殘?zhí)恰⑽r青素產(chǎn)量等發(fā)酵參數(shù)的變化情況。試驗(yàn)結(jié)果如圖4所示。

□-對(duì)照組;■-添加乙醇的實(shí)驗(yàn)組圖3 控制恒定乙醇濃度對(duì)法夫酵母JMU-MVP14發(fā)酵的影響Fig. 3 Fermentation curves of P. rhodozyma JMU-MVP14 at the constant concentration of 2 g/L ethanol

□-對(duì)照組;■-只補(bǔ)加乙醇的實(shí)驗(yàn)組;△-只補(bǔ)加葡萄糖的實(shí)驗(yàn)組;▲-補(bǔ)加乙醇和葡萄糖的實(shí)驗(yàn)組圖4 乙醇補(bǔ)料策略對(duì)法夫酵母JMU-MVP14發(fā)酵的影響Fig.4 The effect of fermentation parameters in fed-batch process with ethanol by P. rhodozyma JMU-MVP14 inshaking flask fermentation
在搖瓶發(fā)酵的3種補(bǔ)料方式中,將乙醇質(zhì)量濃度控制在2 g/L左右,對(duì)法夫酵母JMU-MVP14細(xì)胞生長(zhǎng)最有利,生物量質(zhì)量濃度最大為3.2 g/L。而在只補(bǔ)加葡萄糖的情況下對(duì)法夫酵母細(xì)胞生長(zhǎng)有抑制作用。結(jié)合發(fā)酵液殘?zhí)堑淖兓€可知,每24 h補(bǔ)加5 g/L葡萄糖并未被法夫酵母JMU-MVP14菌株所利用,而是在發(fā)酵液中累積,從而造成發(fā)酵液中葡萄糖的濃度升高。且每24 h補(bǔ)加葡萄糖,較對(duì)照組并未使菌體的生物量增加,但促進(jìn)了蝦青素的合成,這與相關(guān)文獻(xiàn)報(bào)道相一致,高C/N導(dǎo)致細(xì)胞生長(zhǎng)減慢,但有利于蝦青素的積累[30]。法夫酵母JMU-MVP14菌株的3種補(bǔ)料方式中,從蝦青素質(zhì)量濃度變化曲線可知,乙醇和葡萄糖的補(bǔ)加能都促進(jìn)蝦青素的合成,以控制恒定2 g/L乙醇質(zhì)量濃度并每24 h補(bǔ)加5 g/L葡萄糖的培養(yǎng)條件下,蝦青素產(chǎn)量達(dá)到最大值55.3 mg/L,與對(duì)照組比顯著提高44.4%,比只補(bǔ)加乙醇顯著提高了17.4%。圖4-d表明:同時(shí)補(bǔ)加乙醇和葡萄糖,法夫酵母在發(fā)酵120 h其蝦青素細(xì)胞產(chǎn)率為19.7 mg/g,比對(duì)照組(14.1 mg/g)顯著提高,達(dá)到39.7%,比只補(bǔ)加葡萄糖(17.7 mg/g)提高了11.3%,比只補(bǔ)加乙醇16.3 mg/g提高了20.9%。由此可知,同時(shí)補(bǔ)加葡萄糖和乙醇有利于法夫酵母JMU-MVP14菌株細(xì)胞內(nèi)蝦青素的合成。
3 結(jié)論
考察乙醇對(duì)高產(chǎn)蝦青素的法夫酵母菌株JMU-MVP14合成蝦青素的影響,在10 g/L和20 g/L兩種葡萄糖質(zhì)量濃度下,向培養(yǎng)基中添加1 g/L和2 g/L的乙醇時(shí),均可提高蝦青素的產(chǎn)量;而添加20 g/L的乙醇時(shí),則會(huì)抑制法夫酵母的細(xì)胞生長(zhǎng)和蝦青素的產(chǎn)生。在法夫酵母培養(yǎng)基中初始葡萄糖質(zhì)量濃度10 g/L的條件下,通過(guò)補(bǔ)加乙醇控制發(fā)酵液中乙醇的質(zhì)量濃度恒定為2 g/L,蝦青素質(zhì)量濃度為50.1 mg/L,比只在發(fā)酵初始階段添加2 g/L的乙醇,蝦青素質(zhì)量濃度提高了20.3%,蝦青素細(xì)胞產(chǎn)率提高了7.4%,說(shuō)明恒定乙醇控制策略更有利于法夫酵母蝦青素的合成。在較低糖質(zhì)量濃度(10 g/L)培養(yǎng)基中,添加乙醇,蝦青素質(zhì)量濃度最高水平達(dá)到40.0 mg/L,在高糖質(zhì)量濃度(20 g/L)培養(yǎng)基中,添加乙醇,蝦青素質(zhì)量濃度達(dá)到42.0 mg/L,后者比前者多了5%,說(shuō)明乙醇對(duì)法夫酵母產(chǎn)蝦青素的作用與葡萄糖質(zhì)量濃度密切相關(guān)。以10 g/L的葡萄糖質(zhì)量濃度為初始培養(yǎng)基,補(bǔ)料控制整個(gè)發(fā)酵過(guò)程中乙醇質(zhì)量濃度為2 g/L,并每24 h補(bǔ)加5 g/L葡萄糖,蝦青素產(chǎn)量顯著提高,最大值為55.3 mg/L,比對(duì)照組提高了44.4%,比只補(bǔ)加乙醇提高了17.4%;并且法夫酵母在發(fā)酵120 h其蝦青素細(xì)胞產(chǎn)率為19.7 mg/g,比對(duì)照組(14.1 mg/g)提高了39.7%,比只補(bǔ)加葡萄糖(17.7 mg/g)提高了11.3%,比只補(bǔ)加乙醇16.3 mg/g提高了20.9%。由此可知,同時(shí)補(bǔ)加葡萄糖和乙醇有利于法夫酵母JMU-MVP14菌株細(xì)胞內(nèi)蝦青素的合成。但是每24 h補(bǔ)加5 g/L葡萄糖并未被法夫酵母JMU-MVP14菌株所利用,而是在發(fā)酵液中累積,從而造成葡萄糖最終利用率降低,因此后續(xù)試驗(yàn),繼續(xù)對(duì)法夫酵母產(chǎn)蝦青素發(fā)酵過(guò)程中葡萄糖補(bǔ)加的量和時(shí)間進(jìn)行條件優(yōu)化。
[1]黃文文, 洪碧紅, 易瑞灶,等. 蝦青素生產(chǎn)方法及生物活性的研究進(jìn)展[J]. 中國(guó)食品添加劑, 2012(6):214-218.
[2]RAO A R, SINDHUJA H N, DHARMESH S M, et al. Effective inhibition of skin cancer, tyrosinase, and antioxidative properties by astaxanthin and astaxanthin esters from the green algaHaematococcuspluvialis[J]. Journal of Agricultural & Food Chemistry, 2013, 61(16):3 842-3 851.
[3]MAOKA T, TOKUDA H, SUZUKI N, et al. Anti-oxidative, anti-tumor-promoting, and anti-carcinogensis activities of nitroastaxanthin and nitrolutein, the reaction products of astaxanthin and lutein with peroxynitrite[J]. Marine Drugs, 2012, 10(6):1 391-1 399.
[4]KOWSHIK J, BABA A B, GIRI H, et al. Astaxanthin inhibits JAK/STAT-3 signaling to abrogate cell proliferation, invasion and angiogenesis in a hamster model of oral cancer[J]. PloS One, 2014, 9(10):e109114.
[5]項(xiàng)榮, 丁棟博, 李杰. 蝦青素抑癌作用機(jī)制研究進(jìn)展[J]. 天然產(chǎn)物研究與開(kāi)發(fā), 2013, 25(7):1 011-1 015.
[6]AUGUSTI P R, QUATRIN A, SOMACAL S, et al. Astaxanthin prevents changes in the activities of thioredoxin reductase and paraoxonase in hypercholesterolemic rabbits[J]. Journal of Clinical Biochemistry & Nutrition, 2012, 51(1):42-49.
[7]LI Jing-jing, XIA Yu-jing,LIU Tong, et al. Protective effects of astaxanthin on conA-induced autoimmune hepatitis by the JNK/p-JNK pathway-mediated inhibition of autophagy and apoptosis[J]. Plos One, 2015, 10(3):e0120440.
[8]AMBATI R R, MOI P S, RAVI S. Astaxanthin: sources, extraction, stability, biological activities and its commercial applications—areview[J]. Marine Drugs, 2014, 12(1):128-152.
[9]YAMASHITA E. Astaxanthin as a medical food[J]. Functional Foods in Health & Disease, 2013, 3(7):254-258.
[10]WUHai-jian, NIU Huan-jiang,SHAO An-wen, et al. Astaxanthin as a potential neuroprotective agent for neurological diseases[J]. Marine Drugs, 2015, 13(9):5 750-5 766.
[11]BHUVANESWARI S, YOGALAKSHMI B, SREEJA S, et al. Astaxanthin reduces hepatic endoplasmic reticulum stress and nuclear factor-κB-mediated inflammation in high fructose and high fat diet-fed mice[J]. Cell Stress and Chaperones, 2014, 19(2):183-191.
[12]TOMINAGA K, HONGO N, KARATO M, et al. Cosmetic benefits of astaxanthin on humans subjects[J]. Acta Biochimica Polonica, 2012, 59(1):43-47.
[13]陶姝穎, 明建. 蝦青素的功能特性及其在功能食品中的應(yīng)用研究進(jìn)展[J]. 食品工業(yè), 2012(8):110-115.
[14]劉穎. 法夫酵母生物法生產(chǎn)蝦青素的研究進(jìn)展[J]. 發(fā)酵科技通訊, 2012, 41(3):23-27.
[15]蔡俊, 游智能. 發(fā)酵法生產(chǎn)蝦青素的研究進(jìn)展[J]. 食品科學(xué), 2015, 36(23):358-366.
[16]鄭蕾. 法夫酵母產(chǎn)蝦青素發(fā)酵培養(yǎng)基的優(yōu)化[J]. 宿州學(xué)院學(xué)報(bào), 2012, 27(8):60-62.
[17]YAMANE Y, HIGASHIDA K, NAKASHIMADA Y, et al. Astaxanthin production byPhaffiarhodozymaenhanced in fed-batch culture with glucose and ethanol feeding[J]. Biotechnology Letters, 1997,19(11):1 109-1 111.
[18]GUW-L ANG-H, JOHNSONE A. Ethanol increases carotenoid production inPhaffiarhodozyma[J]. Journal of Industrial Microbiology & Biotechnology, 1997,19(2):114-117.
[19]汪洪濤, 徐學(xué)明, 金征宇. 法夫酵母產(chǎn)蝦青素發(fā)酵促進(jìn)劑的研究[J]. 江蘇農(nóng)業(yè)科學(xué), 2006(6):368-370.
[20]肖安風(fēng), 洪清林, 蔡慧農(nóng), 等. 細(xì)胞生長(zhǎng)、葡萄糖代謝及副產(chǎn)物重新利用對(duì)法夫酵母蝦青素合成的影響分析[J]. 高校化學(xué)工程學(xué)報(bào), 2014,28(2):290-297.
[21]張?jiān)d, 周祥山, 陸健,等. 一種用于在線檢測(cè)溶液中甲醇濃度的傳感器: CN 2001.
[22]洪清林.法夫酵母JMU-MVP14菌株產(chǎn)蝦青素發(fā)酵工藝的優(yōu)化[D]. 廈門(mén):集美大學(xué), 2012.
[23]肖冬光, 李賢宇, 郝玥. 高效液相色譜法檢測(cè)紅發(fā)夫酵母胞內(nèi)蝦青素[J]. 天津科技大學(xué)學(xué)報(bào), 2005,20(1):9-12.
[24]JOHNSON E A, LEWIS M J. Astaxanthin formation by the yeastPhaffiarhodozyma[J]. Journal of General Microbiology, 1979,115(1):173-183.
[25]倪輝.法夫酵母蝦青素發(fā)酵條件的優(yōu)化及提取與分析研究[D]. 杭州:浙江大學(xué), 2005.
[26]張龍翔. 生化實(shí)驗(yàn)方法和技術(shù)[M]. 北京:高等教育出版社, 1981.
[27]朱明軍, 梁世中, 陳峰, 等. 利用添加乙醇增加Phaffiarhodozyma蝦青素產(chǎn)量[J]. 華南理工大學(xué)學(xué)報(bào) (自然科學(xué)版), 2001,29(10):61-64.
[28]HU Zhong-Ce,ZHENG Yu-Guo,WANG Zhao, et al. Effect of sugar-feeding strategies on astaxanthin production byXanthophyllomycesdendrorhous[J]. World Journal of Microbiology & Biotechnology, 2005,21(5):771-775.
[29]肖安風(fēng), 倪輝, 李利君, 等. 幾種糖代謝產(chǎn)物對(duì)法夫酵母產(chǎn)蝦青素影響的對(duì)比研究[J]. 食品科學(xué), 2009,30(19):268-272.
[30]YAMANE Y I, HIGASHIDA K, NAKASHIMADA Y, et al. Influence of oxygen and glucose on primary metabolism and astaxanthin production byPhaffiarhodozymain batch and fed-batch cultures: kinetic and stoichiometric analysis.[J]. Applied & Environmental Microbiology, 1997, 63(11):4 471-4 478.
猜你喜歡
中學(xué)生數(shù)理化·中考版(2022年10期)2022-11-10 09:37:42
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年12期)2022-02-14 07:08:42
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(shè)(2018年6期)2018-04-22 03:16:54
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
