梅毒螺旋體黏附素Tp0751核酸菌影的構建及免疫原性研究*
2018-04-20 02:29:36張佳俐曹二龍曹龍古趙飛駿唐一之曾鐵兵
中國免疫學雜志 2018年4期
關鍵詞:小鼠
張佳俐 曹二龍 曹龍古 趙飛駿 余 堅 唐一之 符 波 段 武 曾鐵兵
(南華大學病原生物學研究所,衡陽 421001)
梅毒是由梅毒螺旋體(Treponemapallidum,Tp)感染引起的一種嚴重危害成人和新生兒健康[1]的性傳播疾病,并促進艾滋病傳播[2]。梅毒全球流行,近年來疫情呈上升趨勢[3],尤其在男-男同性性行為者[4]和孕婦[5]中,亟待疫苗的研發。目前尚無疫苗防控梅毒,阻礙疫苗研究的因素主要包括:Tp不可人工培養;Tp外膜蛋白(Outer membrane protein,OMP)稀少[6];梅毒疫苗方法學相對其他病原體疫苗方法學落后;研究者較少[7,8]。
Tp的OMP(免疫應答主要靶位)稀少、缺乏脂多糖,也不產生明顯外毒素,但具有強大侵襲力,能通過生殖道黏膜入侵并經血流播散,因此抗黏附作用一直成為梅毒疫苗的研究焦點[7,9]。Tp0751是目前研究最多和最重要的Tp黏附素[7-9],能吸附血管內皮細胞[10]和細胞外基質(Extracecellular matrix,ECM)[11]并降解后者[12],導致Tp播散[10,12];最近觀察到,以重組Tp0751蛋白免疫新西蘭兔后宿主器官的Tp負荷明顯降低[13],而且Tp0751在各臨床株間高度保守[13],以上表明Tp0751是很有希望的梅毒疫苗候選分子。
目前普遍認為,理想的梅毒疫苗應該能持久誘發系統和局部黏膜的Th1和Th2混合型應答[7-9]。而作為第三代疫苗的核酸疫苗,在應用適當佐劑/DNA遞送系統條件下可誘導上述特點的免疫應答。為此,本研究構建Tp0751的核酸疫苗,并以新型菌影(Bacterial ghost,BG)作為天然佐劑和DNA遞送系統,應用DNA初免-蛋白加免的異源加免(Heterologous prime-boost) 策略,觀察其誘導小鼠產生體液免疫和細胞免疫應答的能力,為發展梅毒新型疫苗奠定基礎。
1 材料與方法
1.1材料
1.1.1質粒、菌株、細胞株與實驗動物 質粒pcDNA3.1(+)、Tp(Nichols株)、空大腸埃希菌菌影(空EBG)、E.coliJM109菌株、鼠源性巨噬細胞Raw264.7均為本研究所保存[14]。雌性BALB/c小鼠購自湖南斯萊克景達實驗動物有限公司。
1.1.2主要試劑 Tp感染兔血清、重組Tp0751蛋白(rTp0751)為本研究所保存;Taq DNA聚合酶、BamHⅠ、XhoⅠ限制性內切酶、T4 DNA連接酶購自美國NEB公司;Tp DNA提取試劑盒購自天根生化科技(北京)有限公司,質粒提取試劑盒、DNA純化試劑盒為美國Omega生物技術公司產品;HRP-羊抗兔IgG、HRP-羊抗鼠IgG與SIgA為美國Proteintech公司產品;CCK-8淋巴細胞增殖試劑盒購自海門碧云天生物技術研究所,IFN-γ ELISA試劑盒為美國eBioscience公司產品。
1.2方法
1.2.1pcD/Tp0751真核表達載體構建 根據GenBank數據庫,設計去除信號肽序列的Tp0751擴增引物,P1:5′CGCGGATCCCGGGACACCGCCGCA 3′(劃線為BamHⅠ酶切位點);引物 P2:5′CCGCTC-GAGTTAGGGCGAAGGAGCACTAGCC 3′(劃線為XhoⅠ酶切位點),引物由上海生工公司合成。以Tp DNA為模板(本所保存),PCR擴增Tp0751基因,將其克隆入pcDNA3.1(+)質粒,構建成真核表達重組體pcD/Tp0751,酶切及測序鑒定(廣州英濰捷基公司測序)。
1.2.2pcD/Tp0751裝載菌影 在制備的100 μl pcD/Tp0751中,依次加入用溫控法預先制備的100 μlE.coliJM109空菌影(50 mg/ml)、250 μl CaCl2(0.1 mol/L),最后加PBS(0.01 mol/L,pH7.4)至總體積1.0 ml,同時用核酸蛋白儀測定質粒濃度(C裝載前),混勻后24℃孵育30 min,然后12 000 r/min離心5 min,取上清測定質粒濃度(C裝載后),計算質粒裝載率:η′=(1-C裝載后/C裝載前)×100%。
1.2.3重組質粒轉染RAW264.7細胞與表達蛋白鑒定 取裝載pcD/Tp0751的菌影100 μl轉染RAW264.7細胞,培養24 h。提取轉染細胞蛋白,電泳轉膜,以Tp感染兔血清(1∶100稀釋)為一抗,HRP-羊抗兔IgG(1∶2 000稀釋)為二抗,Western blot鑒定目的蛋白表達。
1.2.4動物免疫與血清收集 選用6~7周齡的BALB/c雌性小鼠48只,隨機分為6組,每組8只,雙側股四頭肌等量肌注,每次100 μl,間隔兩周免疫一次,共三次。A、B、C組為對照組,分別注射PBS、空EBG[1 mg/(只·次)]、空質粒pcD[20 μg/(只·次)],D、E、F組為實驗組,分別注射裸pcD/Tp0751[20 μg/(只·次)]、pcD/Tp0751-BG[1 mg/(只·次),約含20 μg DNA]、pcD/Tp0751-BG[免疫2次,1 mg/(只·次)]+rTp0751[加免1次,0.3 mg/(只·次)]。首次免疫后第2、4、6、12周,采集小鼠尾靜脈血分離血清,同時收集生殖道沖洗液,-80℃保存備用。
1.2.5特異性血清IgG與生殖道SIgA檢測 用rTp0751(10 μg/ml)以每孔100 μl包被96孔酶標板,以HRP-羊抗鼠IgG/SIgA(1∶10 000稀釋)為二抗,間接ELISA分別檢測待測血清(1∶100稀釋)和生殖道沖洗液(1∶50稀釋)中特異性IgG和SIgA的A450值。測定F組在末次加免后IgG和SIgA效價。
1.2.6淋巴細胞增殖試驗 首次免疫后第12周處死小鼠,無菌取小鼠脾臟,制備淋巴細胞,調整細胞數至1×104/孔,陽性對照每孔以100 μg ConA、陰性對照組加PBS、實驗組每孔以10 μg rTp0751刺激培養48 h。培養結束前4 h,每孔加入CCK-8溶液10 μl(5 mg/ml),培養至結束,酶標儀測定A450值,計算刺激指數(SI)=刺激組A450值/無刺激陰性對照組A450值。
1.2.7IFN-γ分泌水平檢測 調整淋巴細胞數至1×105/孔,每孔以10 μg rTp0751刺激培養48 h,離心細胞,收集上清液,用ELISA試劑盒檢測IFN-γ水平,測定A450值,以標準品做標準曲線,計算IFN-γ含量。
1.3統計學分析 實驗數據用SPSS13.0軟件進行統計學分析,每組標本取平均值,應用t-Test檢驗,P<0.05表示差異有統計學意義。
2 結果
2.1真核表達載體構建鑒定與菌影裝載率 真核重組表達質粒pcD/Tp0751經雙酶切得到大小與預期位置相符的5 400 bp和555 bp兩個片段(圖1),測序結果與公布Tp0751序列完全一致,表明真核重組質粒構建成功。重組質粒對空菌影的裝載率η′=(1-15.6/65.4)×100%=76.1%。
2.2pcDNA3.1(+)/Tp0751真核重組蛋白鑒定 WB結果顯示,重組質粒裝載的菌影作用RAW264.7細胞24 h后,細胞表達的目的蛋白與Tp感染兔血清出現了明顯的特異性反應(圖2),表明重組質粒轉染細胞后成功表達目的蛋白。
2.3特異性抗體水平 D、E、F實驗組小鼠特異性血清 IgG與生殖道SIgA水平均隨免疫次數增加而增加,各時間點均顯著高于三個對照組(P<0.01),于末次加免后第8周(初免后第12周)達到峰值,此時F組IgG與SIgA效價分別為1∶102 400與1∶12 800;首次加免2周(初免后第4周)及以后,E、F組均顯著高于D組(P<0.01);末次加免2周(初免后第6周)及以后,F組顯著高于E組(P<0.01)(圖3A、B)。
2.4小鼠脾淋巴細胞增殖 首次免疫后第12周(末次加免后第8周),D、E、F組的SI均顯著高于A、B和C組(P<0.01),E、F組均顯著高于D組(P<0.01),F組高于E組(P<0.05)(表1)。
2.5鼠脾淋巴細胞分泌IFN-γ水平 首次免疫后第12周(末次加免后第8周),D、E、F組的IFN-γ水平均顯著高于A、B和C組(P<0.01),E、F組均顯著高于D組(P<0.01),F組高于E組(P<0.05)(圖4)。
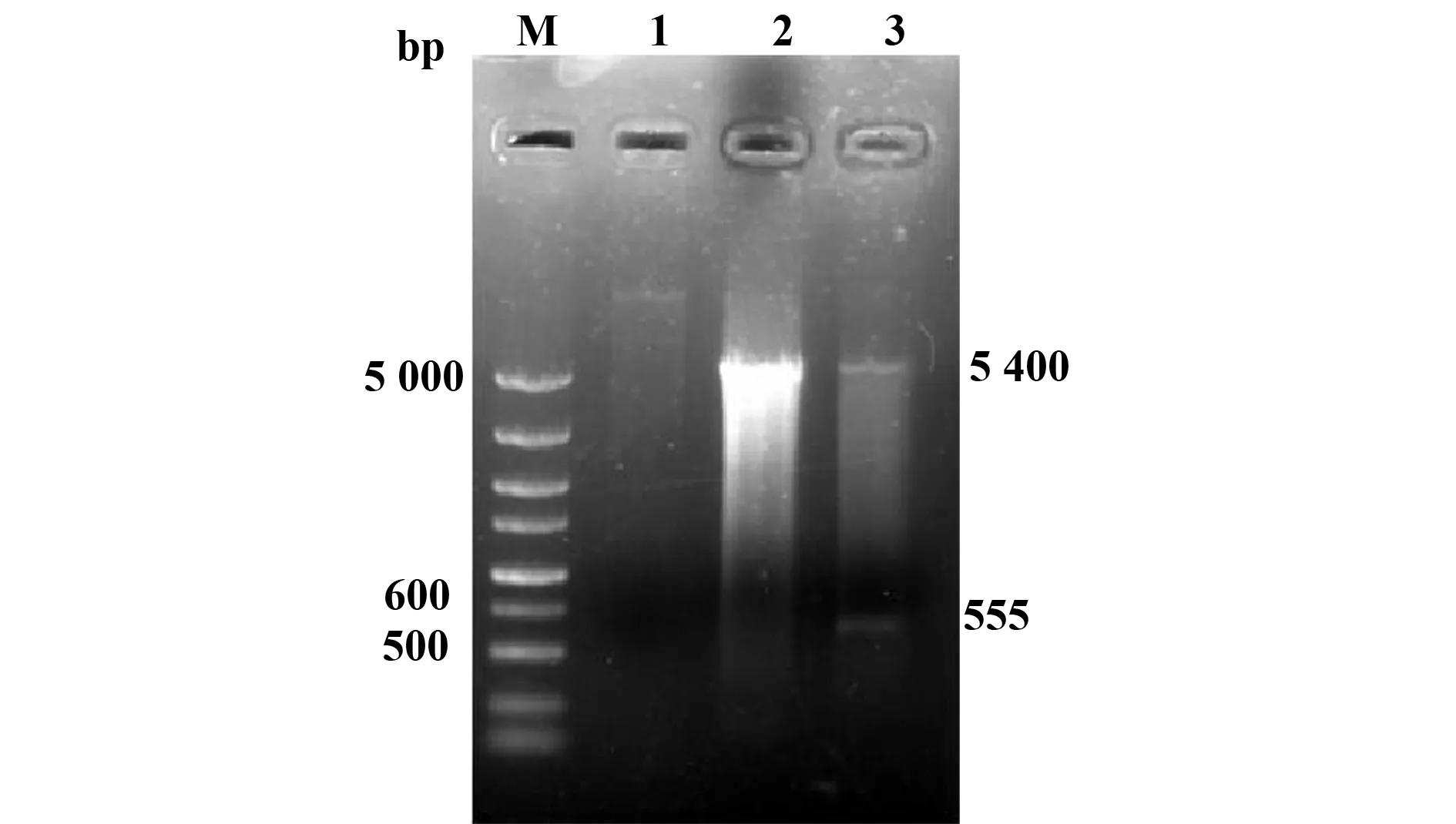
圖1 pcDNA3.1/Tp0751重組質粒的酶切鑒定Fig.1 Identification of recombinant pcDNA3.1/Tp0751 digested with BamHⅠ and Xho ⅠNote: M.DNA marker;1.Negtive control;2.Empty pcDNA3.1;3.Recombinant pcDNA3.1/Tp075.

圖2 Tp0751真核重組蛋白的Western blot鑒定Fig.2 Identification of eukaryotic expression of recombi-nant Tp0751 by Western blotNote: 1.RAW264.7 cells untreated with pcD/Tp0751-BG;2,3.RAW264.7 cells treated with pcD/Tp0751-BG;4.RAW264.7 cells treated with empty pcDNA3.1.

圖3 不同時間點各免疫組小鼠特異性血清IgG(A)及生殖道SIgA(B)Fig.3 IgG and SIgA against Tp0751 in vaccinated mice at different time pointNote: Vs groups A,B,C,*.P<0.01;vs group D,#.P<0.01;vs group E,△.P<0.01.
表1小鼠脾淋巴細胞刺激指數(SI)
Tab.1StimulationIndexofspleenlymphocytesofimmun-izedmice

GroupsSIA:PBS1.019±0.135B:EBG1.079±0.025C:pcDNA1.032±0.031D:pcD/Tp07511.447±0.1111)E:pcD/Tp0751-BG9.896±0.2591)2)F:pcD/Tp0751-BG+rTp075111.985±0.3191)2)3)G:ConA2.318±0.145
Note:Vs groups A,B,C,1)P<0.01;vs group D,2)P<0.01;vs group E,3)P<0.05.

圖4 小鼠脾淋巴細胞分泌IFN-γ水平(pg/ml)Fig.4 Levels of IFN-γ produced by spleen lymphocytes in vaccinated mice(pg/ml)Note: Vs groups A,B,C,*.P<0.01;vs group D,#.P<0.01;vs group E,△.P<0.05
3 討論
重組蛋白疫苗是目前國外唯一應用的梅毒疫苗類型,但存在一些不足[7],如通過E.coli高水平表達時可能難以準確保持天然蛋白的構象;易降解,純化技術要求高;以常規佐劑免疫難以誘導局部黏膜的免疫應答等,因此需發展其他類型的疫苗[7-9]。核酸疫苗是近20年來發展起來的第三代疫苗。與傳統疫苗相比,核酸疫苗具有明顯的優勢[15,16]:能模擬病原體自然感染,在宿主細胞內表達近乎天然構象的目的蛋白,不僅能持久誘導體液免疫,而且能產生強有力的細胞免疫。此外,DNA疫苗還具有制備簡單、使用安全等諸多優點成為近年研究的熱點,具有良好的應用前景。
核酸疫苗在真核細胞中的有效轉染和表達目的蛋白是誘導免疫應答的前提。本研究中 pcD/Tp0751真核質粒在轉染真核細胞RAW264.7中能有效表達目的蛋白,為下一步研究提供了基礎。
目前普遍認為,Th1型免疫應答是早期清除Tp的主要機制[7-9],通過CD4+Th1細胞等分泌IFN-γ為主[17]的Th1型細胞因子活化巨噬細胞,促進其吞噬殺傷Tp[18]。本研究發現,pcD/Tp0751-BG同源加免和異源加免均在末次加免后8周仍能誘導非常明顯的小鼠脾淋巴細胞增殖和IFN-γ分泌,表明Tp0751的核酸菌影能有效誘導小鼠系統性細胞免疫應答。
體液免疫應答也是必不可少的[7-9],局部黏膜特異性抗黏附素的SIgA能早期阻止Tp從生殖道黏膜吸附上皮細胞而入侵[8],而血清中特異性IgG等能防止Tp黏附血管內皮細胞從而向遠處組織播散,還能特異性介導巨噬細胞靶向吞噬殺傷Tp(調理吞噬)[19],與細胞免疫應答協同作用最終清除Tp[7-9]。本研究發現,pcD/Tp0751-BG同源加免和異源加免誘導IgG與SIgA水平均隨著免疫次數的增加而上升,于末次加免后8周到達峰值,水平最高的異源加免組效價能分別達到1∶102 400和1∶12 800,表明Tp0751的核酸菌影能有效誘導小鼠系統性與黏膜的體液免疫應答。
質粒DNA本身免疫原性低,選擇合適的DNA遞送系統和佐劑對有效誘導應答十分關鍵。近年,菌影(BG)作為理想的天然抗原(DNA或蛋白)遞送系統和佐劑引起極大關注,目前已被廣泛地用于沙眼衣原體、幽門螺桿菌、HIV等許多病原體的疫苗[20,21]。BG是革蘭陰性細菌被溫控法或化學法裂解后失去胞漿成分而保留完整細胞壁和細胞膜結構和天然形態的細菌空殼。除了裝載重組蛋白外,BG還具有強大的外源DNA裝載能力,每個BG可在裂解通道中裝載數千個DNA拷貝,而且裝載簡單高效[20],在本研究中,BG裝載重組DNA質粒的裝載率達到了76%,顯示出高效的裝載能力;同時,BG保留了顆粒狀活菌天然的表面抗原結構(脂多糖、肽聚糖、菌毛等),APC表面的模式識別受體很容易將其識別和靶向性攝入而被充分活化,繼而激活T細胞啟動適應性免疫應答[20]。因此,BG 也是良好的天然免疫佐劑。菌影這些特性使其能增強DNA/重組蛋白疫苗誘導系統和局部的細胞免疫和體液免疫應答水平,且能維持持久免疫記憶[22]。本研究發現,pcD/Tp0751-BG同源加免和異源加免誘導體液免疫和細胞免疫的水平均明顯高于不加佐劑的裸pcD/Tp0751,而且在末次免疫后8周能達到最高水平,表明能維持較好的免疫記憶。
為進一步提供DNA疫苗誘導免疫應答水平,本研究設立了DNA初免兩次、繼以重組Tp0751蛋白加免一次的異源加免(prime-boost)組。結果顯示,與三次DNA同源加免組相比,異源加免誘導的體液免疫應答和細胞免疫應答均更高。當前DNA初免-蛋白加免(異源加免)被認為是理想的免疫策略,比同一蛋白或DNA的同源加免能誘導更強的免疫應答,但機制不甚明確,可能是DNA疫苗與蛋白疫苗以不同的機制刺激特異性應答。DNA疫苗能誘導細胞免疫應答,同時也高效誘導記憶性B細胞產生,當以蛋白抗原加免時,直接活化記憶性B細胞[23]。
本實驗首次構建Tp0751的真核重組菌影,有效誘導了小鼠系統和黏膜體液免疫應答及細胞免疫應答,下一步擬對免疫鼠進行抗Tp攻擊感染實驗以驗證其免疫保護性,為進一步研發新型梅毒疫苗奠定基礎。
參考文獻:
[1] Lawn JE,Blencowe H,Waiswa P,etal.Stillbirths:rates,risk factors,and acceleration towards 2030 [J].Lancet,2016,387(10018):587-603.
[2] Douglas JM.Penicillin treatment of syphilis:clearing away the shadow on the land [J].JAMA,2009,301(7):769-771.
[3] Abara WE,Hess KL,Neblett Fanfair R,etal.Syphilis trends among men who have sex with men in the United States and Western Europe:A systematic review of trend studies published between 2004 and 2015 [J].PLoS One,2016,11(7),e0159309.
[4] Peterman TA,Su J,Bernstein KT.Syphilis in the United States:on the rise?[J].Exp Rev Anti Infect Ther,2015,13(2):161-168.
[5] Wijesooriya NS,Rochat RW,Kamb ML,etal.Global burden of maternal andcongenital syphilis in 2008 and 2012:a health systems modelling study[J].Lancet Glob Health,2016,4:e525-e533.
[6] LaFond RE,Lukehart SA.Biological basis for syphilis [J].Clin Microbiol Rev,2006,19(1):29-49.
[7] Cullen PA,Cameron CE.Progress towards an effective syphilis vaccine:the past,present and future [J].Exp Rev Vaccines,2006,5(1):67-80.
[8] Lithgow KV,Cameron CE.Vaccine development for syphilis [J].Expert Rev Vaccines,2017,16(1):37-44.
[9] Cameron CE,Lukehart SA.Current status of syphilis vaccine development:need,challenges,prospects [J].Vaccine,2014,32(14):1602-1609.
[10] Kao WA,Pětrosová H,Ebady R,etal.Identification of Tp0751 (Pallilysin) as atreponemapallidumvascular adhesin by heterologous expression in the lyme disease spirochete [J].Sci Rep,2017,7(1):1538.
[11] Cameron CE,Brouwer NL,Tisch LM,etal.Defining the interaction of thetreponemapallidumadhesin Tp0751 with laminin[J].Infect Immun,2005,73(11):7485-7494.
[12] Houston S,Hof R,Honeyman L,etal.Activation and proteolytic activity of thetreponemapallidummetalloprotease,pallilysin[J].PLoS Pathog,2012,8(7):e1002822.
[13] Lithgow KV,Hof R,Wetherell C,etal.A defined syphilis vaccine candidate inhibits dissemination oftreponemapallidumsubspecies pallidum [J].Nat Commun,2017,8:14273.
[14] 江銀波,曹二龍,朱 洪,等.梅毒螺旋體粘附素Tp0751重組菌影的構建與鑒定[J].中南醫學科學雜志,2016,44(2)121-124,129.
Jiang YB,Cao EL,Zhu H,etal.Constructiong and identification of recombinant E.Coli bacterial ghosts expressingtreponemapallidumadhesin Tp 0751[J].Medical Science J Central South China,2016,44(2):121-124,129.
[15] Saade F,Petrovsky N.Technologies for enhanced efficacy of DNA vaccines[J].Exp Rev Vaccines,2012,11(2):189-209.
[16] 李開銘,謝梅花,余敏君,等.B群腦膜炎奈瑟菌外膜蛋白NMB0315核酸疫苗的免疫活性和免疫保護作用初步研究[J].中國免疫雜志,2015,31(12):1648-1653,1658.
Li KM,Xie MH,Yu M,etal.Immunocompetence and immunoprotection of a DNA vaccine NMB0315 against N.meningitidis sergroup B in mice[J].Chin J Immunol,2015,31(12):1648-1653,1658.
[17] Leader BT,Godornes C,VanVoorhis WC,etal.CD4+lymphocytes and gamma interferon predominate in local immune responses in early experimental syphilis[J].Infect Immunol,2007,75(6):3021-3026.
[18] Baker-Zander SA,Lukehart SA.Macrophage-mediated killing of opsonizedTreponemapallidum[J].J Infect Dis,1992,165(1):69-74.
[19] Cruz AR,Ramirez LG,Zuluaga AV,etal.Immune evasion and recognition of the syphilis spirochete in blood and skin of secondary syphilis patients:two immunologically distinct compartments[J].PLoS Negl Trop Dis,2012,6(7):e1717.
[20] Muhammad A,Champeimont J,Mayr UB,etal.Bacterial ghosts as carriers of protein subunit and DNA-encoded antigens for vaccine applications[J].Exp Rev Vaccines,2012,11(1):97-116.
[21] Langemann T,Koller VJ,Muhammad A,etal.The bacterial ghost platform system:production and applications[J].Bioeng Bugs,2010,1(5):326-336.
[22] Eko FO,Ekong E,He Q,etal.Induction of immune memory by a multisubunit chlamydial vaccine[J].Vaccine,2011,29(7):1472-1480.
[23] Kardani K,Bolhassani A,Shahbazi S.Prime-boost vaccine strategy against viral infections:Mechanisms and benefits[J].Vaccine,2016,34(4):413-423.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34