小麥淀粉合成關鍵酶基因和相關蛋白表達對不同施磷量的響應
2018-05-11 09:36:11李春艷張潤琪付凱勇朱長安
麥類作物學報 2018年4期
李春艷,張潤琪,付凱勇,李 超,朱長安,李 誠
(1.石河子大學農學院/新疆兵團綠洲生態農業重點實驗室,新疆石河子 832000;2.新疆盛源種業有限責任公司,新疆五家渠 831300)
小麥是中國主要的糧食作物之一,其產量和品質對國家糧食和食品安全有舉足輕重的戰略意義。磷素是小麥生長過程中重要的大量元素之一,對小麥產量和淀粉品質形成有重要作用。深入細化研究磷素在小麥淀粉產量和品質形成中的作用,可有效緩解磷礦資源日益匱乏及過量施用磷肥造成的環境和經濟問題,為小麥生產“減肥、增效、提質”提供理論依據。
普通小麥籽粒中淀粉約占干重的60%~70%。淀粉的生物合成是一個復雜的過程,涉及多種酶的共同作用。腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)是淀粉合成的關鍵酶,無機磷酸與3-PGA的比例會影響其活性,并影響淀粉的生物合成過程[1]。顆粒結合淀粉合成酶(GBSS)主導直鏈淀粉的延長,也可能對支鏈淀粉側鏈的延長有一定的作用[2]。GBSS I存在于造粉體中,幾乎完全是顆粒結合酶[3]。淀粉粒結合蛋白(SGPs)主要包括淀粉粒表面的和鑲嵌在淀粉顆粒內部的蛋白[4]。目前,已在小麥中檢測到4種主要的淀粉粒結合蛋白:SGP-1、SGP-2、SGP-3和Waxy蛋白,各SGP分別對應不同的淀粉合成相關酶[5]。通過改良蛋白提取方法,結合2D電泳、質譜、TEM免疫雜交和突變體材料,Benmoussa等[6]從玉米淀粉粒蛋白中鑒別出結構蛋白、膜蛋白以及與淀粉合成重要相關的酶類(GBSS、AGPase等)。Matsushima等[7-8]通過水稻突變體發現與淀粉粒增大和質體發育有關的蛋白SSG4和SSG6,并將其定位于水稻胚乳質體的被膜。除此之外,關于淀粉合成/代謝過程的重要基因和蛋白的研究正在逐步深入[9-11]。但小麥生長過程中施用不同水平的磷肥對胚乳可溶性蛋白和淀粉粒結合蛋白有何影響尚未見報道。因此,本研究從小麥籽粒灌漿進程、關鍵酶基因表達、酶活性、蛋白含量和譜帶特征以及胚乳微觀特性等角度入手,研究不同施磷量對小麥籽粒發育的影響,以期為小麥生產中磷肥的合理施用以及深入研究小麥淀粉的生物合成機理提供理論依據。
1 材料與方法
1.1 試驗材料與田間管理
供試材料為新疆主栽冬小麥品種新冬20號,種子由石河子大學麥類作物研究所提供。新冬20號具有廣泛的適應性,產量潛力較高(7 500~8 250 kg·hm-2),抗逆性較強。試驗于2014 年10 月至2015 年6 月在石河子大學農學院試驗站進行。試驗地0~20 cm土壤有機質15.4 g·kg-1,堿解氮63 mg·kg-1,速效磷15 mg·kg-1,速效鉀208 mg·kg-1。試驗地前茬為小麥,收獲后種植向日葵,并將向日葵作為綠肥壓青,壓青時施入75 kg·hm-2尿素作為基肥。試驗設3個磷素(以P2O5計)水平:0 kg·hm-2(對照,記作CK)、105 kg·hm-2(常規施磷,低磷,記作LP)和210 kg·hm-2(過量施磷,高磷,記作HP),隨機區組設計,每處理3次重復。所用磷肥為過磷酸鈣,含P2O515.57%,于小麥返青期一次性開溝條施。小麥行距20 cm,在兩行中間開溝,溝深5~10 cm。對照開溝,但不施肥。冬前滴灌水3次,返青期至成熟期滴灌水6次,每隔10~12 d滴灌1次,拔節期和揚花期各追施尿素75 kg·hm-2。尿素為市售產品,N含量46%。6月28日種子成熟后收獲。
1.2 樣品采集
小麥開花后,選擇穗形一致的主莖標記開花期,分別于開花后7、14、21、28、35 d取樣。從標記穗子中隨機摘取150穗,剝出穗中部籽粒,一部分烘干至恒重,備用;一部分用鋁箔紙包裹,立即置于液氮速凍5 min,并在-80 ℃超低溫冰箱保存,備用。
1.3 籽粒灌漿速率測定
以開花后天數為自變量,每次取樣所得平均單粒重(由千粒重折合)為因變量,用Logistic方程y=K/(1+ae-bt)對灌漿過程進行擬合[12],式中,K為最大單粒重,t為開花后天數,a和b為常數。由Logistic方程推導出一系列灌漿參數包括籽粒灌漿速率最大值Vmax和籽粒灌漿速率最大值出現時間Tmax。
1.4 淀粉合成關鍵酶活性測定
AGPase活性測定參照Nakamura等[13]的方法;GBSS活性測定參照Schaffer等[14]的方法。
1.5 agp和gbss基因相對表達量檢測
1.5.1 引物設計
根據NCBI公布的小麥 agp1、 agp2、 gbss1和 gbss2基因序列設計引物,引物由生工生物工程(上海)股份有限公司合成。以小麥actin基因(NCBI:DN551593)作為內參基因。通過梯度PCR驗證引物特異性并優化PCR條件。PCR反應體系:cDNA模板0.5 μL,10×PCR反應緩沖液2 μL,dNTPs 1.25 μL,上游引物和下游引物各0.5 μL,TaqDNA聚合酶0.75 μL,雙蒸水4.5 μL。反應條件:94 ℃ 5 min,94 ℃ 35 s,退火30 s,72 ℃ 30 s,35個循環,72 ℃10 min。具體引物序列見表1。
1.5.2 RNA提取和cDNA合成
RNA提取采用Fruit-mate(TaKaRa,9192,Japan)和RNAiso plus(TaAaRa,9108,Japan)試劑盒提取,按說明書操作。瓊脂糖凝膠電泳檢驗總RNA質量。使用TIANScript cDNA第一鏈合成試劑盒(Tiangen,KR104-02,China)合成cDNA,用合成的cDNA作為模板擴增actin基因以檢驗其質量。
表1實時熒光定量PCR引物序列
Table1PrimersusedtodetectdifferentialgeneexpressionbyusingQuantitativePCR

基因Gene上游引物序列(5′-3′)Forwardprimersequence下游引物序列(5′-3′)Reverseprimersequence退火溫度/℃Annealingtemperatureagp1AGAAGCGGCAAGGGAGACCCTAATACATCAAGGGAGAAAA60agp2TCAAACAAGGAGGGAGTGGCAGAGGTTGTCGCAGA60gbss1GACACTATCGTGGAAGGCAAGTTGACCATCTCATGGTACGC60gbss2CACAGAATGCCAGAGGCATAGGAACAGATGGGAATCACTCCA60actinGGAAAAGTGCAGAGAGACACGTACAGTGTCTGGATCGGTGGT60
1.5.3 實時熒光定量PCR
使用SYBR Premix ExTaq試劑盒(TaKaKa,RR420A,Japan)和實時熒光定量PCR儀(Roche LightCycler 480 Ⅱ)檢測各基因的擴增情況,具體操作按說明書進行。
1.6 胚乳可溶性蛋白和淀粉粒結合蛋白的提取及含量測定
胚乳可溶性蛋白和淀粉粒結合蛋白的提取參照Peng等[15]的方法;蛋白質含量測定參考劉子凡[16]的考馬斯亮藍G-250法,用牛血清白蛋白繪制標準曲線;SDS-PAGE聚丙烯酰胺凝膠電泳參照高正等[17]的考馬斯亮藍R-250染色法進行。
1.7 胚乳橫切面的甲苯胺藍染色及掃描電鏡觀察
胚乳橫切面的甲苯胺藍染色方法參照余靜等[18]的文獻。用于掃描電鏡觀察的樣品處理程序:取不同處理的籽粒約3 mm厚的橫切片置于帶蓋小瓶,加入5 mL磷酸鈉緩沖液配制的4%(v/v)戊二醇溶液,4 ℃固定24 h,室溫靜置 1 h,再用磷酸鈉緩沖液漂洗3次,每次10 min,用系列濃度(35%、50%、70%、85%、95%)酒精脫水,每次10 min,再用100%酒精脫水3次,每次10 min,經E3100 CO2臨界點干燥儀(Quorum Technologies-Ringmer,East Sussex,UK)干燥及離子濺射儀(Denton Vacuum-Moorestown,NJ,USA)鍍金膜后,用掃描電鏡(Hitachi S570,Hitachi High Technologies,Tokyo,Japan)觀察拍照,加速電壓10 kV。
1.8 統計分析
采用Microsoft Office Excel 2013和SPSS 17.0統計軟件整理分析數據,采用Microsoft Office Excel 2013和Adobe Photoshop軟件繪制圖表。每項指標重復測定3次。采用獨立t檢驗法比較2個樣本平均數,采用Duncan新復極差法進行多重比較,顯著水平為0.05。
2 結果與分析
2.1 小麥粒重及灌漿速率的變化
由表2可知,3種施磷水平下,小麥品種新春20號在相同取樣時期的粒重差異顯著。花后7 d 和14 d,低磷處理下的粒重顯著低于對照;花后7、21和35 d,高磷處理小麥粒重顯著高于低磷和對照;花后28 d和35 d,低磷處理的粒重顯著高于對照。說明籽粒灌漿后期磷肥對粒重的促進作用逐漸增強。
由表3可知,不同施磷處理對籽粒灌漿速率最大值出現時間無明顯影響,均為花后16 d。低磷處理下,小麥籽粒灌漿最大積累速率顯著高于對照和高磷處理。
表2不同磷素水平下小麥籽粒的單粒重
Table2Singlegrainweightofwheatunderdifferentphosphorustreatmentsmg

處理 Treatment7DAA14DAA21DAA28DAA35DAACK6.94±0.12b16.01±0.06a31.4±0.07c38.02±0.01b40.8±0.02cLP6.74±0.013c14.74±0.01c32.01±0.02b38.20±0.05a41.0±0.03bHP7.34±0.09a15.6±0.02b32.21±0.01a37.8±0.05c41.4±0.08a
表中數據為“平均值±標準誤”,同列數據后不同小寫字母表示在5%水平差異顯著。DAA:開花后天數。表4同。
Values are “mean±standard error” of three replications.Values followed by different letters within columns are significantly different atP<0.05.DAA:Days after anthesis.The same in table 4.
表3不同磷素水平下小麥籽粒灌漿速率特征參數
Table3Parametersofgrainfillingrateofwheatunderdifferentphosphorustreatments

處理Treatment回歸方程RegressionequationR2Vmax/(mg·d-1)Tmax/dCKy=41.673/(1+21.604e-0.196t)0.998??2.042b15.678aLPy=41.730/(1+25.313e-0.205t)0.994??2.137a15.778aHPy=42.248/(1+21.151e-0.194t)0.991??2.048b15.739a
Vmax:籽粒灌漿速率最大值;Tmax:籽粒灌漿速率最大值出現時間。
Vmax:The maximum grain-filling rate;Tmax:The time reaching the maximum grain-filling rate.
2.2 不同磷處理對agp和gbss基因相對表達量的影響
由圖1可知,花后7、14和28 d,低磷處理下 agp1基因的相對表達量顯著高于對照,同時顯著或極顯著高于高磷處理;而高磷處理下 agp1基因的相對表達量在花后14、21 和28 d 顯著或極顯著低于對照(圖1A)。低磷處理下, agp2基因的相對表達量在花后7 d和14 d均極顯著高于對照和高磷處理。花后21 d和35 d,高磷處理下 agp2基因相對表達量極顯著高于對照(圖1B)。低磷處理下, gbss1基因相對表達量在花后7~35 d 均顯著或極顯著高于對照(圖1C),除花后14 d外,均顯著或極顯著高于高磷處理;花后14 d 和21 d該基因的相對表達量在對照和高磷處理間差異不顯著。除花后21 d外,低磷處理下 gbss2基因的相對表達量均顯著或極顯著高于對照和高磷處理;高磷處理下,花后7 d和14 d該基因相對表達量極顯著低于對照(圖1D)。
2.3 不同磷處理對小麥籽粒AGPase和GBSS活性的影響
AGPase是小麥合成、貯存多糖過程的限速酶。由圖2可知,不同施磷處理下,小麥籽粒中AGPase活性均呈“M”型變化趨勢,花后7、21和28 d,各處理AGPase活性無顯著差異;花后14 d,低磷處理下AGPase活性極顯著高于高磷處理;花后35 d,高磷處理下AGPase活性顯著低于對照和低磷處理。GBSS主要負責直鏈淀粉合成。花后7 d,不同處理間GBSS活性差異不顯著;花后14 d,低磷處理下GBSS活性分別顯著或極顯著高于對照和高磷處理;花后21 d,施磷處理下GBSS活性均極顯著高于對照。花后28 d和35 d,低磷和高磷處理下GBSS活性差異顯著,與對照無顯著差異(圖2)。
2.4 不同施磷水平對小麥淀粉粒結合蛋白和胚乳可溶性蛋白含量的影響
由表4可知,花后14 d,高磷處理淀粉粒結合蛋白含量顯著高于對照和低磷處理;花后21 d,低磷處理淀粉粒結合蛋白含量顯著高于對照和高磷處理;花后28 d,對照處理淀粉粒結合蛋白含量顯著高于施磷處理;花后35 d高磷處理淀粉粒結合蛋白含量最高。

*和**分別代表0.05和0.01水平上對照和施磷處理間差異顯著;以小麥actin基因CNCBI編號:DN551593)作為內參基因。下圖同。
* and ** represent the significance of probability at 0.05 and 0.01 levels between CK and P treatment,respectively;Wheat actin gene CNCBI Accession No. DN551593) was used as the reference control. The same in Fig.2.
圖1不同磷處理下小麥胚乳agp和gbss基因相對表達量的變化
Fig.1Relativeexpressionofagpandgbssgenesinwheatendospermunderdifferentphosphorustreatments
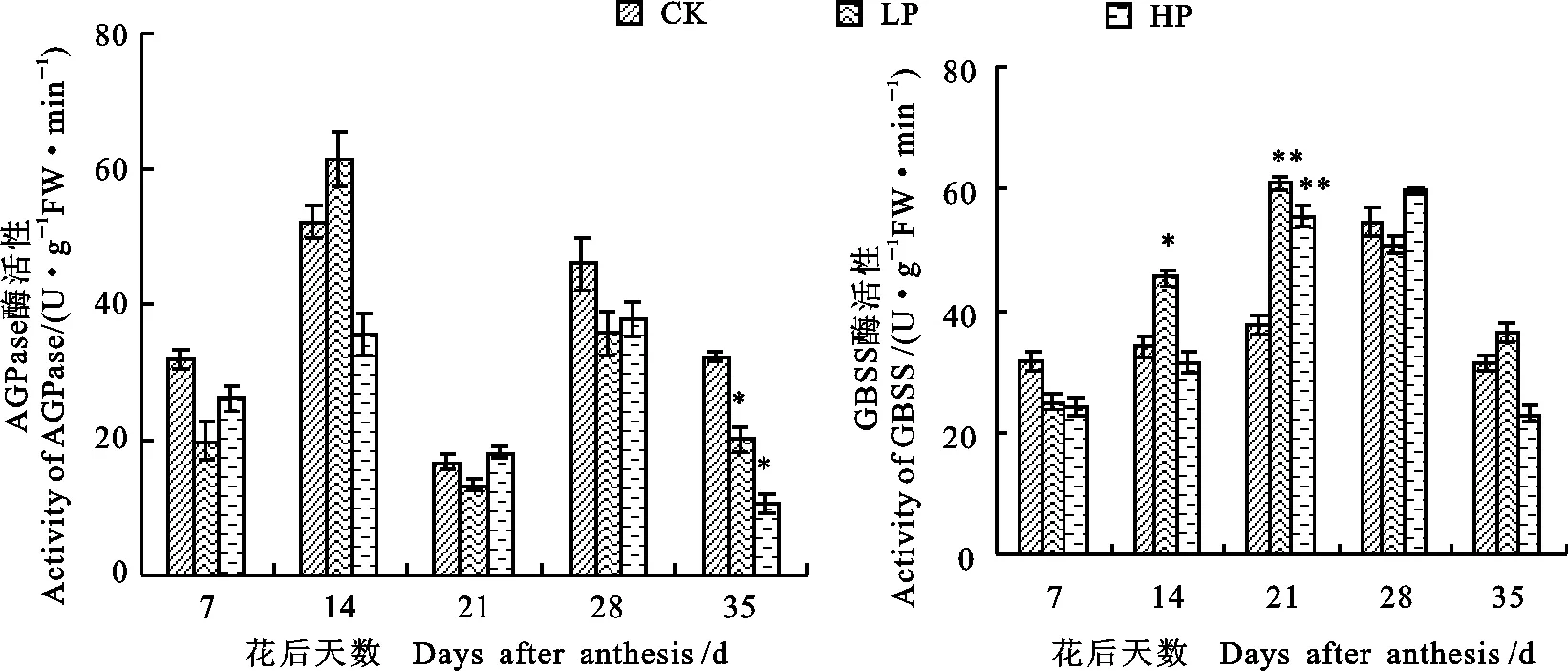
圖2 不同磷處理下小麥籽粒AGPase和GBSS活性
小麥籽粒AGPase主要存在于胚乳可溶性部分,該酶是由2個大亞基和2個小亞基組成的異源四聚體。2個大亞基分子質量為55~60 kD,2個小亞基分子質量為50~55 kD,大亞基是酶活性的調節中心,而小亞基是酶活性的催化中心[19]。本研究通過聚丙烯酰氨凝膠電泳分析,經濃縮分離的胚乳可溶性蛋白條帶分子質量均小于72 kD,其中2條分子質量在60 kD左右,另外3條清晰的條帶分別出現在26~43、34~26和17 kD左右。推測分子質量在72~55 kD的蛋白可能與AGPase有關(圖3)。施磷處理下,花后7 d 胚乳可溶性蛋白條帶的表達量均明顯弱于對照;花后35 d,3種處理下分子質量為26~34 kD的條帶(黑色箭頭)均較其他取樣時期明顯。
表4不同磷水平下小麥淀粉粒結合蛋白含量
Table4Contentofstarchgranule-boundproteinunderdifferentphosphorustreatments

×10-3 μg·mg-1

圖3 不同施磷水平下小麥胚乳可溶性蛋白凝膠電泳
2.5 不同施磷水平對小麥胚乳微觀特性的影響
由圖4可知,花后21 d,甲苯胺藍染色顯示對照和高磷處理下胚乳中被染色的細胞核(圖中紅色箭頭所示)數量多于低磷處理。低磷和高磷處理下甲苯胺藍著色較深低。低磷處理下小麥胚乳中可以觀察到明顯的顆粒狀淀粉粒,而相同放大倍數下,對照和高磷處理下的淀粉粒則不明顯。對照籽粒斷面細胞排列緊密,胚乳外層柵欄狀細胞與內層胚乳細胞結合緊密,兩者分層不明顯(圖4A);低磷和高磷處理下胚乳外部柵欄狀細胞形態明顯,細胞排列較松散,與內胚乳細胞結合不緊密,兩者分層明顯(圖4B和圖4C),而低磷處理此現象較高磷處理更明顯。
掃描電鏡結果顯示,不同磷處理下胚乳淀粉粒形態未發生明顯變化,均由透鏡狀的A型淀粉粒(直徑大于10 μm)和球形的B型淀粉粒(直徑小于10 μm)組成。淀粉粒均鑲嵌于蛋白基質中。低磷處理下蛋白基質較少,主要存在于淀粉粒間,部分附著在淀粉粒表面,淀粉粒與蛋白質結合較疏松(圖4E);而對照和高磷處理的蛋白質基質很多,形成完整的蛋白質包被鞘,淀粉粒深陷在包被中,淀粉粒和蛋白質結合較緊密(圖4D和圖4F),其中高磷處理下的此現象更明顯。
3 討 論
淀粉是小麥籽粒中含量最多的物質,與小麥的產量與品質有直接關系[20-21]。本研究發現新冬20號籽粒灌漿不同時期磷素對粒重的影響存在處理間差異;花后7 d和14 d,低磷處理粒重顯著低于對照。前期研究亦表明,花后7 d低磷處理下淀粉粒的形態與對照差異明顯,且平均粒徑顯著低于對照[22],可能是該施磷水平促進了小麥灌漿前期的營養生長,并為灌漿后期籽粒的營養積累儲備了能量。在籽粒灌漿后期(花后28~35 d),相同取樣時期高磷和低磷處理下小麥籽粒的成熟度高于對照,且施磷后總淀粉、支鏈淀粉和直鏈淀粉含量顯著高于對照,并以低磷處理下最高[23]。此結果與籽粒灌漿關鍵時期淀粉合成相關酶AGPase和GBSS的活性和酶基因表達量的趨勢一致。淀粉合成的第一步是由G-1-P和ATP在AGPase作用下生成腺苷二磷酸葡萄糖(ADPG),ADPG是淀粉合成的直接前體,因此,AGPase是淀粉合成的限速酶[1]。GBSS則主導直鏈淀粉的延長[2]。AGPase和GBSS活性和酶基因表達量的提高對淀粉的合成與積累有重要作用。小麥籽粒灌漿過程中胚乳細胞的分裂、增殖以及充實對磷素的需求可能存在一個臨界范圍,準確把握不同小麥品種籽粒灌漿過程中對磷素需求的適宜范圍可以增加磷肥的有效利用率并提高小麥產量。
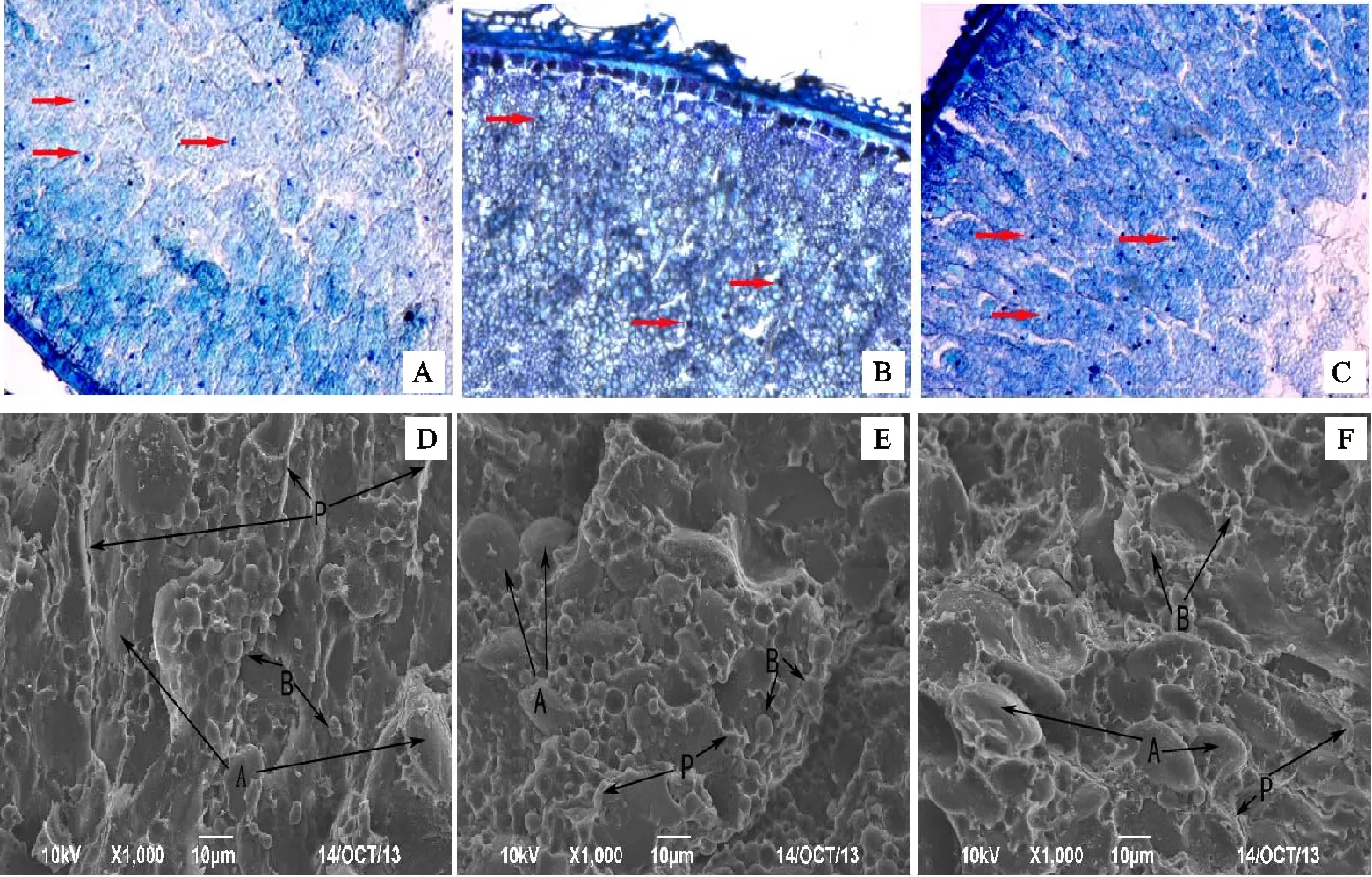
A、B、C:胚乳橫切面甲苯胺藍染色(×130);D、E、F:胚乳橫切面掃描電鏡圖片(×1 000);A、D:CK處理;B、E:LP 處理; C、F:HP 處理。電鏡掃描圖D、E、F中A指示為A型淀粉粒,B指示B型淀粉粒;P指示蛋白基質。
A,B and C:Cross sections stained by toluidine(×130); D,E and F:Cross sections observed by scanning electron microscopy(×1 000); A and D were samples from CK treatment; B and E were samples from LP treatment; C and F were samples from HP treatment.Within the SEM figures D,E,and F,‘A’ indicates the A type starch granules; ‘B’ indicates the B type starch granules; ‘P’ indicates the protein matrix.
圖4花后21d不同磷處理下小麥籽粒橫切面的差異
Fig.4Crosssectionsofwheatgrainat21DPAunderdifferentphosphorustreatments
小麥、玉米、高粱和谷子等禾本科作物淀粉粒表面均能觀察到微孔,是淀粉粒本身的屬性特征。Han等[24]通過特異性蛋白探針(CBQCA)處理淀粉粒并用激光共聚焦顯微鏡觀察到淀粉粒通道內存在蛋白。玉米中關于淀粉粒通道蛋白的研究已有報道,但小麥中鮮見報道。Benmoussa等[6]從玉米淀粉粒通道蛋白中鑒別出與淀粉合成相關的酶類(GBSS、AGPase等),認為玉米淀粉粒微通道可能是淀粉質體微管的殘留物,對淀粉合成中ADPG的運輸有重要作用,便于淀粉聚合物和顆粒的合成。本研究中,不同磷處理下各時期淀粉粒結合蛋白譜無明顯差異(因無顯著差異故圖片未顯示),均呈5條主要的譜帶,且分子質量均大于60 kD,分別對應SGP-A1、SGP-B1、SGP-D1、SGP-2、SGP-3和Waxy蛋白[5]。花后35 d時淀粉粒結合蛋白的含量以高磷處理最高,胚乳可溶性蛋白也以高磷處理下花后28~35 d的表達量最高,此結果與掃描電鏡觀察籽粒橫切面的蛋白基質狀態一致。說明施磷處理對小麥籽粒淀粉和蛋白質的合成均有重要影響,而與淀粉合成相關的酶類(AGPase、GBSS等)又可促進淀粉的合成,其中可能包括淀粉粒通道蛋白。石河子大學農學院冬小麥課題組研究發現,低磷和高磷處理下淀粉粒微通道結構較對照更加明顯[23],參照玉米中的研究[6],推測磷處理下淀粉粒的通道蛋白組分發生了變化,GBSS、AGPase含量增加,促進了淀粉的生物合成。該假設需進一步研究驗證。
前人研究表明,小麥淀粉粒甲苯胺藍染色結果的深淺與支鏈淀粉的精細結構和淀粉粒的晶體結構有關。支鏈淀粉分支鏈較短[25],分支化程度高[25-26],淀粉粒結構片層密度較低[27],可能使甲苯胺藍分子容易進入淀粉粒內部結合蛋白質[28]和帶負電荷的磷酸酯[29]將小麥淀粉染成藍色。本研究中,與低磷和高磷處理比較,對照處理下甲苯胺藍染色較淺。石河子大學農學院冬小麥課題組前期研究表明,花后21 d,不同磷素處理下新冬20號小麥籽粒支鏈淀粉結構中A鏈的比例表現為HP>LP>CK,淀粉粒的結晶度表現為CK>HP>LP[21],說明施磷處理導致淀粉粒結構松散。此外,石河子大學農學院冬小麥課題組通過激光共聚焦顯微鏡發現,低磷和高磷處理下的淀粉粒內部的微通道結構發生了變化[23];再結合該課題組研究發現,高磷和低磷處理下淀粉粒直徑顯著小于對照[22],小淀粉粒的比表面積大于大淀粉粒,可以結合更多的蛋白,因此,低磷和高磷處理下甲苯胺藍染色較CK顏色深。綜上所述,通過甲苯胺藍染色淀粉粒可以在一定程度上推測淀粉粒內部的結構。
參考文獻:
[1] MACDONALD P W,STROBEL G A.Adenosine diphosphate-glucose pyrophosphorylase control of starch accumulation in rust-infected wheat leaves [J].PlantPhysiology,1970,46(1):126.
[2] RAHMAN S,LI Z,BATEY I,etal.Genetic alteration of starch functionality in wheat [J].JournalofCerealScience,2000,31(1):91.
[3] IZUMO A,FUJIWARA S,SAKURAI T,etal.Effects of granule-bound starch synthase I-defective mutation on the morphology and structure of pyrenoidal starch in Chlamydomonas [J].PlantScience,2011,180(2):238.
[4] RAHMAN S,KOSARHASHEMI B,SAMUEL M S,etal.The major proteins of wheat endosperm starch granules [J].FunctionalPlantBiology,1995,22(5):793.
[5] YAMAMORI M,ENDO T R.Variation of starch granule proteins and chromosome mapping of their coding genes in common wheat [J].TheoreticalandAppliedGenetics,1996,93(1-2):275.
[6] BENMOUSSA M,HAMAKER B R,HUANG C P,etal.Elucidation of maize endosperm starch granule channel proteins and evidence for plastoskeletal structures in maize endosperm amyloplasts [J].JournalofCerealScience,2010,52(1):22.
[7] MATSUSHIMA R,MAEKAWA M,KUSANO M,etal.Amyloplast membrane protein SUBSTANDARD STARCH GRAIN6 controls starch grain size in rice endosperm [J].PlantPhysiology,2016,170(3):1445.
[8] MATSUSHIMA R,MAEKAWA M,KUSANO M,etal.Amyloplast-localized SUBSTANDARD STARCH GRAIN4 protein influences the size of starch grains in rice endosperm [J].PlantPhysiology,2014,164(2):623.
[9] FEIKE D,SEUNG D,GRAF A,etal.The starch granule-associated protein EARLY STARVATION1(ESV1) is required for the control of starch degradation in Arabidopsis thaliana leaves [J].PlantCell,2016,28(6):1472.
[10] SHAIK S S,OBATA T,HEBELSTRUP K H,etal.Starch granule re-structuring by starch branching enzyme and glucan water dikinase modulation affects caryopsis physiology and metabolism [J].PLoSONE,2016,11(2):e0149613.
[11] YU H T,WANG T.Proteomic dissection of endosperm starch granule associated proteins reveals a network coordinating starch biosynthesis and amino acid metabolism and glycolysis in rice endosperms [J].FrontiersinPlantScience,2016,7:707.
[12] 王曉慧,張 磊,劉雙利,等.不同熟期春玉米品種的籽粒灌漿特性[J].中國農業科學,2014,47(18):3560.
WANG X H,ZHANG L,LIU S L,etal.Grain filling characteristics of maize hybrids differing in maturities [J].ScientiaAgriculturaSinica,2014,47(18):3560.
[13] NAKAMURA Y,YUKI K,PARK S Y.Carbohydrate metabolism in the developing endosperm of rice grains [J].PlantCellPhysiology,1989,30(6):833.
[14] SCHAFFER A A,PETREIKOV M.Sucrose-to-starch metabolism in tomato fruit undergoing transient starch accumulation [J].PlantPhysiology,1997,113(3):739.
[15] PENG M,GAO M,ESM A A,etal.Separation and characterization of A-and B-type starch granules in wheat endosperm [J].CerealChemistry,1999,76(3):375.
[16] 劉子凡.種子學實驗指南[M].北京:化學工業出版社,2011:34.
LIU Z F.Seed science:A laboratory manual [M].Beijing:Chemical Industry Press,2011,34.
[17] 高 正,王曉龍,張曉科,等.小麥HMW-GS檢測方法的優化及應用[J].麥類作物學報,2013,33(5):931.
GAO Z,WANG X L,ZHANG X K,etal.Optimization and application of the detection method of HMW-GS in common wheat [J].JournalofTriticeaeCrops,2013,33(5):931.
[18] 余 靜,冉從福,李學軍,等.陜糯1號與非糯小麥西農1330 胚乳發育及淀粉形態、粒徑分析 [J].中國農業科學,2014,47(22):4413.
YU J,RAN C F,LI X J,etal.Study on endosperm development and morphological features of starch granules in waxy wheat Shannuo 1 and non-waxy wheat Xinong 1330 [J].ScientiaAgriculturaSinica,2014,47(22):4413.
[19] JOHNSON P E,PATRON N J,BOTTRILL A R,etal.A low-starch barley mutant,Ris 16,lacking the cytosolic small submit of ADP-Glucose pyrophosphorylase,reveals the importance of the cytosolic isform and the identity of the plastidial small subunit [J].PlantPhysiology,2003,131(2):684.
[20] NI Y,WANG Z,YIN Y,etal.Starch granule size distribution in wheat grain in relation to phosphorus fertilization [J].JournalofAgriculturalScience,2012,150(1):45.
[21] LI C Y,LI C,ZHANG R Q,etal.Effect of phosphorus on the characteristics of starch in winter wheat [J].Starch/St?rke,2013,65(9):801.
[22] 付凱勇,徐芳芳,史曉艷,等.不同磷素水平對小麥淀粉粒形態及品質特性的影響[J].石河子大學學報(自然科學版),2015,33(4):413.
FU K Y,XU F F,SHI X Y,etal.Effect of phosphorus on the morphology and quality characteristics of starch granules in wheat [J].JournalofShiheziUniversity(NaturalScience),2015,33(4):413.
[23] ZHANG R Q,LI C,FU K Y,LI C,LI C Y.Phosphorus alters starch morphology and gene expression related to starch biosynthesis and degradation in wheat grain [J].FrontiersinPlantScience,2017,8:2252.
[24] HAN X Z,HAMAKER B R.Location of starch granule-associated proteins revealed by confocal laser scanning microscopy [J].JournalofCerealScience,2002,35(1):109.
[25] YOO S H,JANE J.Structural and physical characteristics of waxy and other wheat starches [J].CarbohydratePolymers,2002,49(3):297.
[26] 李春燕,封超年,王亞雷,等.不同小麥品種支鏈淀粉鏈長分配及其與淀粉理化特性的關系 [J].作物學報,2007,33(8):1240.
LI C Y,FENG C N,WANG Y L,etal.Chain length distribution of debauched amylopectin and its relationship with physicochemical properties of starch in different wheat cultivar [J].ActaAgronomicaSinica,2007,33(8):1240.
[27] JANE J,CHEN Y Y,LEE L F,etal.Effects of amylopectin branch chain length and amylose content on the gelatinization and pasting properties of starch [J].CerealChemistry,1999,76(5):629.
[28] SVIHUS B,UHLEN A K,HARSTAD O M.Effect of starch granule structure,associated components and processing on nutritive value of cereal starch:A review [J].AnimalFeedScienceandTechnology,2005,122(3):303.
[29] BALDWIN P M.Starch granule-associated proteins and polypeptides:A review [J].Starch/St?rke,2015,53(10):475.
