以色列野生二棱大麥籽粒氮素含量的基因型及其生態差異
2018-05-11 09:36:13程曉彬阮景軍程劍平NEVOEviatar
麥類作物學報 2018年4期
關鍵詞:環境
程曉彬,李 念,阮景軍,趙 鋼,程劍平,NEVO Eviatar,嚴 俊
(1.四川民族學院環境與生命科學系,四川康定 626001; 2.貴州大學麥作研究中心,貴州貴陽 550025;3.成都大學藥學與生物工程學院,四川成都 610106; 4.海法大學進化研究所,以色列海法 31905)
大麥是世界上最古老的作物之一,具有早熟、生育期短、適應性廣、豐產以及營養豐富等特性,分為栽培大麥和野生大麥2種。大麥因兼具食用、飼用、釀造用以及醫藥等多種用途而受到人們越來越多的重視[1]。大麥育種多以高產為主要目的,在人工選擇過程中容易失去其長期進化產生的遺傳多樣性,而遺傳多樣性的缺乏和喪失會導致大麥對環境及病蟲害的抗性減弱[2-3]。野生二棱大麥[Hordeumspontancum(Aberg) Shao]受生態環境的影響,在長期自然選擇過程中積累了大量抗病、抗蟲、耐寒、耐旱、耐貧瘠、耐鹽等多種抗逆基因[4],是大麥遺傳育種、品種改良的重要材料[5]。中東新月沃地(包括以色列、約旦等國)是野生二棱大麥起源地和遺傳多樣性中心。野生二棱大麥是栽培大麥的祖先,可直接用于雜交育種,也可作為轉基因技術育種的目的基因供體[6]。
籽粒蛋白質含量是評價大麥品質優劣的標準之一,提高籽粒蛋白質含量是大麥育種工作的重要目標[7]。對于飼用大麥,通常育種目標是在注重提高品種產量的同時提高其蛋白質含量;而對于釀造大麥,高蛋白會導致啤酒釀造過程中糖化困難,易混濁,但如果麥芽的蛋白質含量過低,其釀造的啤酒也可能因酵母氨基酸含量過低而影響品質,所以培育籽粒中氮素含量高或適中的大麥品種是滿足各種市場需求的首要任務。野生大麥豐富的遺傳多樣性可為該項育種工作提供豐富的遺傳資源[7-8]。氣候條件對大麥籽粒蛋白質含量也有一定的影響,年份不同,其籽粒蛋白質含量變化較大[9-10]。本研究對采自以色列不同生長環境的9個群體90個野生二棱大麥基因型的有機氮及無機氮營養水平進行分析測定,以期篩選出不同蛋白質含量和氮素含量的大麥基因型,為栽培大麥種質創新研究提供一定的實驗基礎和理論依據。
1 材料與方法
1.1 材 料
供試種子采自以色列從南到北不同區域,共9個群體90個基因型(表1)。于2012和2013年種于貴州大學麥作農場、2013年種于安順農科所試驗農場和貴州大學園藝農場,共4個處理,分別為:2012貴州大學麥作農場田間(Y2012 Farm)、2013安順農科所試驗農場田間(Y2013 AS)、2013貴州大學麥作農場盆栽(2013 9F)和2013貴州大學園藝農場田間(Y2013 Garden)。貴州大學麥作農場位于106.4°~106.9°E,26.2°~26.6°N,海拔1 100 m;安順農科所試驗農場(Y2013 AS)位于105.9°E,26.3°N,海拔1 395 m。田間試驗采用隨機區組排列,播種方式為條播,行長1.5 m,行間距0.4 m,一行為同一基因型,每個基因型3次重復,三葉期定苗,每行留長勢基本一致的5株苗;盆栽實驗每盆裝土8.0 kg,一盆一個基因型,每個基因型3次重復,三葉期定苗,每盆留長勢一致的3株苗。麥穗成熟后收獲,室內自然風干后脫粒,去雜和挑去不完整籽粒,用去離子水將種子洗凈烘干后妥善保存,備用。
1.2 分析方法
樣品分析在貴州大學麥作研究中心實驗室完成,隨機取10粒籽粒用萬分之一電子天平稱量,計算單粒重;按照國標法(GB2905-82)采用KJ 8400凱氏定氮儀測定籽粒總蛋白質含量,蛋白質N素換算系數為6.25;通過UV1000分光光度計采用考馬斯藍染色法測定可溶性蛋白含量。所有測定設3次重復,取其平均值進行數據分析。
1.3 數據分析
采用Microsoft Excel 2007處理數據,變異系數≥25 %的基因型重測,直到<25%為止。采用JMP6.0和SigmaPlot12.0分析數據,通過Tukey HSD檢驗進行多重比較,P<0.05為差異顯著,P<0.01為差異極顯著。
表1試驗材料
Table1Materialsusedintheexperiment

群體Population樣品數量Samplequantity基因型GenotypeGolanHigh20GN1GN2GN3GN4GN6GN7GN9GN10GN11GN12GN18GN19GN20GN21GN22GN25GN26GN28GN29GN30Shlomi20SH1SH3SH4SH6SH7SH9SH10SH12SH13SH15SH16SH17SH18SH19SH20SH21SH22SH23SH24SH25Hermon13HM1HM2HM3HM4HM5HM7HM9HM14HM16HM17HM18HM19HM20Mehola3ME13ME14ME15Atlit1A5Tabigha8T4T5T6T7T8T9T12T14RoshPina14R1R2R3R6R7R10R11R12R13R15R16R17R19R23Gilboa9G3G4(M)G4(R)G8G10G11G19G20G21ECNorth?facingslope2NF1NF2
2 結果與分析
2.1 不同環境下籽粒總氮含量及單粒重的差異
經Tukey HSD檢驗方法分析,Y2012 Farm和Y2013 AS環境下90個野生大麥基因型籽粒總氮及單粒重差異顯著(圖1),在Y2012 Farm環境下野生大麥籽粒總氮含量顯著高于Y2013 AS,Y2013 AS環境下單粒重顯著高于Y2012 Farm。由此可見,環境對野生大麥籽粒總氮含量和單粒重影響顯著,單粒重變化趨勢與籽粒中總氮含量變化相反,說明每粒種子能富集的氮素有一定上限并具有一定的保守性。
2.2 不同環境下籽粒可溶性蛋白質含量的差異
4個環境下野生二棱大麥籽粒可溶性蛋白質存在差異(圖2)。在2013AS環境下野生大麥籽粒可溶性蛋白質含量顯著高于其他3個環境,Y2013 Garden、2012 Farm和2013 9F三個環境間差異不顯著,說明野生二棱大麥籽粒氮素營養積累受種植環境的影響。在Y2013 AS環境下可溶性蛋白質含量最高,但總氮含量相對偏低,而在Y2012 Farm環境下有相反的結果,這對調控大麥庫爾巴哈值有一定意義。

不同小寫字母表示差異顯著(P<0.05)。下同。
Letters above the bars indicate significant differences (P<0.05).The same below.
圖1不同環境下籽粒總氮含量及單粒重的差異
Fig.1Differenceofgraintotalnitrogenandsinglegrainweightintwoenvironments
2.3 不同群體氮素相關指標的比較
2.3.1 不同環境下群體總氮含量的比較
對種植于Y2013 AS和Y2012 Farm環境下6個基因型較多的野生二棱大麥群體籽粒含氮量進行分析,結果(圖3)顯示,2個環境中Tabigha群體平均總氮含量均最高,在Y2012 Farm環境中顯著高于Golan High 和 Shlomi 2個群體,在Y2013 AS環境中顯著高于Hermon群體。在Y2012 Farm環境下,Golan High群體平均總氮含量最低,但與群體Shlomi、Gilboa和 Hermon無顯著差異,僅與Tabigha和RoshPina群體差異顯著,此外,Shlomi、Gilboa、 Hermon和RoshPina 4個群體相互間也無顯著差異。在Y2013 AS環境下,Hermon群體平均總氮含量最低,但與除Tabigha外的其他5個群體間并無顯著差異。由此可以看出,不同群體的野生大麥在氮素營養性狀上存在一定的進化差異和保守性。

圖2 不同環境下籽粒可溶性蛋白質含量的差異

圖3 不同野生大麥群體平均籽粒含氮量的差異
2.3.2 不同環境下群體可溶性蛋白質含量的比較
對6個野生二棱大麥群體籽粒可溶性蛋白質含量進行Tukey HSD檢驗發現(圖4),在Y2013 9F和Y2013 Garden 2個環境下,Rosh Pina群體的可溶性蛋白質含量顯著高于Golan High和Shlomi群體。在Y2013 Garden環境中,Gilboa群體的可溶性蛋白質含量顯著高于Golan High和Shlomi群體;在Y2012 Farm環境中,Gilboa群體的可溶性蛋白質含量顯著高于Golan High和Rosh Pina群體;在Y2013 AS環境中,Gilboa群體顯著高于Hermen、Golan High、Tabigha和Shlomi群體。總體來看,Y2012 Farm、Y2013 AS及Y2013 Garden 3個環境中的Gilboa群體的可溶性蛋白質含量最高,均分別顯著高于各自環境下的2個以上群體。此外,每個環境下野生二棱大麥籽粒可溶性蛋白質含量在4個群體間均無顯著差異。以上結果說明,野生二棱大麥籽粒可溶性蛋白質含量仍具有一定的保守性。
2.4 極端氮素營養基因型的篩選結果
由表2可知,籽粒總氮含量較高的基因型有 T8、 R3、 GR13和 R23,其中 T8最高,為3.42%;可溶性蛋白質含量較高的基因型有 R23、 T8、 G20、 R3和 GR13,其中 T8最高,為10.22%。綜合分析可知,籽粒總氮和可溶性蛋白質含量同時都較高的基因型為 T8,均較低的是 HM17。不同基因型野生大麥的平均可溶性蛋白含量和平均總氮含量均不同,變異幅度相差較大,說明野生二棱大麥基因型籽粒可溶性蛋白和總氮含量存在豐富的遺傳多樣性,基因型 T8和 HM17均可作為改良現代栽培大麥的優異基因材料。
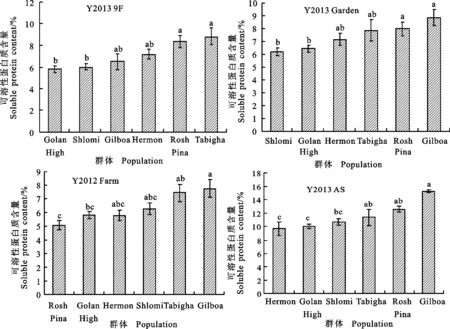
圖4 不同野生大麥群體籽粒可溶性蛋白含量的差異
Table2Comparisonofgenotypeswithsignificantgrainnitrogen-relatedindicators

基因型Genetype平均籽粒可溶性蛋白含量Averagegraincontentofsolubleprotein/%平均籽粒總氮含量Averagegraintotalnitrogencontent/%GN267.35d2.01dGN286.10e2.35cR2310.06ab3.04bR39.49c3.12bR199.28c2.96bT810.22a3.42aG209.92b2.93bG219.26c3.06bHM173.23f2.00d
3 討 論
3.1 不同環境下籽粒總氮含量及可溶性蛋白質含量的差異
從野生大麥群體中篩選極端氮素水平的種質資源用于大麥育種,是提高和改良大麥蛋白質含量的有效方法和途徑[11]。本研究中,不同環境籽粒總氮含量和單粒重差異顯著,單粒重變化趨勢與籽粒中總氮含量變化相反,說明每粒種子富集的氮素具有一定上限并具有一定的保守性。Y2013 AS環境野生大麥籽粒可溶性蛋白質含量顯著高于其他3個環境,Y2013 Garden、Y2012 Farm和2013 9F三個環境間的可溶性蛋白質含量差異不顯著,說明野生二棱大麥籽粒氮素營養積累既受種植環境的影響,又有一定保守性。本研究選用的種子材料均由中以合作發掘與利用野生麥類資源優異基因課題組(成都大學與海法大學聯合成立)采自以色列自然環境,與之前選自以色列海法大學種子庫的群體和基因型不同,但仍得到一些相似的結果,尤其在同樣種植環境下,個別群體氮素營養顯著高于或低于其他群體,說明起源地生態地理環境對籽粒氮素積累有影響,并且籽粒氮素營養在長期的進化和對環境的適應過程中逐步穩定并形成了豐富的遺傳多樣性[12]。
3.2 不同基因型籽粒總氮、可溶性蛋白質含量和單粒重的比較
大麥高產、高蛋白質含量主要受遺傳因素和環境因素的影響[13-14],大麥蛋白質含量屬于數量性狀遺傳且受多個微效基因控制,單個基因效應較小,易受環境條件影響,且在分離世代中表現出連續變異,難以明確分組[10]。因此,在傳統的蛋白質遺傳研究中,通常將控制同一性狀的多基因看作一個整體,運用生物統計的方法計算其均值、方差、顯性效應、上位效應、加性效應等參數,用這些遺傳參數來描述群體的遺傳特征[15]。本研究發現,離散程度較明顯的基因型有 GN26、 HM17、 T8、 GR13、 R3和 R23。 GN26和 HM17分別為籽粒總氮和可溶性蛋白質含量較低的基因型; T8為籽粒總氮和可溶性蛋白質含量均較高的個體, R3、 R23和 GR13分別為籽粒總氮和可溶性蛋白質含量較高的基因型。由于研究條件限制,此次篩選出來的基因型個體在某些性狀上可能存在不足,測定的相關數據僅供參考,在今后的試驗中將進一步擴大種植環境與年份,選用更多起源地具有未知性狀的野生大麥種質材料,分析大麥籽粒蛋白質含量的變化特征和氮素積累機制。通過高、低蛋白大麥種質間的雜交構建遺傳群體,對籽粒蛋白質含量的遺傳特性及控制基因進行分析,盡可能探明影響籽粒蛋白質含量的遺傳效應和環境效應的關系,可為選育受環境條件影響小的優質大麥品種提供理論指導。
參考文獻:
[1] 盧良恕.中國大麥學[M].北京:中國農業出版杜,1996:1,58,93.
LU L S.Chinese barley science [M].Beijing:China Agriculture Press,1996:1,58,93.
[2] 方桂英,杜向紅,張保軍,等.約旦野生大麥種質資源形態與產量性狀分析 [J].西北農業學報,2010,19(6):82.
FANG G Y,DU X H,ZHANG B J,etal.Analysis of agronomic traits ofHordeumspontaneumC.Koch germplasm from Jordan [J].JournalofNorthwestAgriculturalSciences,2010,19(6):82.
[3] 禹代林,洛桑更堆,歐 珠,等.西藏野生大麥種質資源研究概述 [J].西藏農業科技,1998,20(2):18.
YU D L,LUO S G D,OU Z,etal.A Survey of wild barley germplasm resources in Tibetan [J].TibetanJournalofAgriculturalSciences,1998,20(2):18.
[4] 程曉彬,趙 鋼,薛文韜,等.約旦野生二棱大麥在川西高原的物候期和農藝性狀分析 [J].西南農業學報,2016,29(5):1027.
CHENG X B,ZHAO G,XUE W T,etal.Analysis of phenophase and agronomic characters of wild barley(Hordeumspontaneum) originated from Jordan in western Sichuan plateau [J].SouthwestChinaJournalofAgriculturalSciences,2016,29(5):1027.
[5] 任喜峰,孫東發.大麥雜交組合蛋白質及部分農藝性狀的遺傳分析 [J].湖北農業科學,2008,47(4):400.
REN X F,SUN D F.Genetic analysis of protein content and some agronomic traits in annual wild barley from Qing-Tibetan plateau [J].HubeiAgriculturalSciences,2008,47(4):400.
[6] 朱彩梅,張 京.中國糯大麥品種資源及地理分布研究 [J].中國農業科學,2008,41(12):4248.
ZHU C M,ZHANG J.Geographical distribution of waxy barley germplasm resources in China [J].ScientiaAgriculturaSinica,2008,41(12):4248.
[7] 黃志仁,周美學,黃友圣,等.大麥籽粒蛋白質含量的配合力研究 [J].遺傳學報,1991,18(3):264.
HUANG Z R,ZHOU M X,HUANG Y S,etal.Combining ability analysis for protein content in barley grain [J].JournalofGenetics,1991,18(3):264.
[8] 張想平,雷耀湖,何慶祥,等.黑糯大麥的品種特性及營養品質分析 [J].種子,2011,30(9):85.
ZHANG X P,LEI Y H,HE Q X,etal.Analysis on characteristic and nutrition quality of black and waxy barley [J].Seed,2011,30(9):85.
[9] 朱彩梅,張 京.應用SSR標記分析中國糯大麥種質的遺傳多樣性 [J].植物遺傳資源報,2010,11(1):63.
ZHU C M,ZHANG J.Genetic diversity analysis of waxy barley in China based on SSR markers [J].JournalofPlantGeneticResources,2010,11(1):63.
[10] 胡立勇,丁艷峰.作物栽培學(第1版)[M].北京:高等教育出版社,2008:574.
HU L Y,DING Y F.Crop cultivation(first edition) [M].Beijing:Higher Education Press,2008:574.
[11] 李 念,阮景軍,嚴 俊,等.以色列野生二棱大麥籽粒多組分營養性狀的相關性分析 [J].安徽農業大學學報,2016,43(6):1021.
LI N,RUAN J J,YAN J,etal.Correlation analysis on multiple nutrition traits ofHordeumspontaneumoriginating from different locations in Israel [J].JournalofAnhuiAgriculturalUniversity,2016,43(6):1021.
[12] 文玲玲,薛文韜,嚴 俊,等.約旦野生二棱大麥群體籽粒休眠特性的網絡分析 [J].植物生理學報,2014,50(8):1235.
WEN L L,XUE W T,YAN J,etal.The network analysis on characters of caryopsis dormancy in wild barley (Hordeumspontaneum) populations from Jordan [J].PlantPhysiologyJournal,2014,50 (8):1235.
[13] HAJMANSOOR S,BIHAMTA M R,ALISOLTANI A.Genetic diversity among and within Iranian and non-Iranian barely (HordeumvulgareL.) genotypes using SSR and storage proteins markers [J].BiochemicalSystematicsandEcology,2013,46:15.
[14] 辛培堯,普曉英,杜娟,等.大麥籽粒和苗粉蛋白質含量的檢測 [J].麥類作物學報,2016,36(1):60.
XIN P Y,PU X Y,DU J,etal.Protein content determination of Barley grain and seedling powder [J].JournalofTriticeaeCrops,2016,36(1):60.
[15] 汪軍妹,張新忠,林 峰,等.73 份優異啤酒大麥種質資源主要麥芽品質性狀的評價 [J].浙江農業學報,2012,24(1):4.
WANG J M,ZHANG X Z,LIN F,etal.The assessment of major malt qualities among 73 elite varieties in the germplasm for malting barley [J].ActaAgriculturaeZhejiangensis,2012,24(1):4.
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學院學報(2017年2期)2017-04-20 08:44:31
中國環境監察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38
