靶向抑制TROP 2基因表達對胰腺癌細胞生物學特性的影響研究
2018-05-16 11:42:02周童張遠鵬張啟文
癌癥進展 2018年3期
關(guān)鍵詞:差異
周童,張遠鵬,張啟文
山東省萊蕪市人民醫(yī)院普外科,山東萊蕪2711000
胰腺癌是一種常見的消化系統(tǒng)惡性腫瘤,具有惡性程度高和預后差等特點[1]。該病的發(fā)生、發(fā)展涉及原癌基因的激活和抑癌基因的失活等復雜過程,因此,從分子水平研究該病的發(fā)生、發(fā)展機制,對臨床治療有重要的意義[2]。腫瘤相關(guān)鈣信號傳導因子 2(tumor-associated calcium signal transducer 2,TROP2)是近年發(fā)現(xiàn)的與腫瘤密切相關(guān)的基因,是一種可以調(diào)節(jié)腫瘤細胞生長的信號傳導分子[3]。在正常的組織中TROP2不表達或低表達,而在肺癌和食管癌等腫瘤細胞中高表達,其表達與腫瘤的發(fā)生、發(fā)展及預后等相關(guān),因此TROP2可以作為一種預測癌癥預后的獨立因子[4-5]。有研究指出,在一些腫瘤細胞中,如大腸癌和乳腺癌等,抑制TROP2基因表達,可以降低癌細胞的侵襲和遷移能力[6-7],然而TROP2基因在胰腺癌中的作用機制尚不清楚。本研究通過RNA干擾技術(shù)抑制胰腺癌細胞中TROP2基因的表達,觀察細胞的增殖和凋亡情況,進一步研究其作用機制,為研究胰腺癌的基因治療提供新的思路。
1 材料與方法
1.1 材料
收集2016年6月至2017年7月山東省萊蕪市人民醫(yī)院保存的胰腺癌組織和相應的癌旁組織(距離腫瘤邊緣1~2 cm)各46例。所有患者的病歷資料完整且術(shù)前均未行化療和放療,所有組織均經(jīng)病理學檢查確診,所有患者及家屬均對本研究知情同意。46例標本所屬患者中,男24例,女22例;年齡為35~71歲,平均為(54.2±8.5)歲;組織病理學分級按照Kloppel標準分為高分化腺癌11例,中分化腺癌25例,低分化腺癌10例;臨床分期按照國際抗癌協(xié)會(UICC)1992年標準分為Ⅰ~Ⅱ期20例,Ⅲ~Ⅳ期26例;有淋巴結(jié)轉(zhuǎn)移27例,無淋巴結(jié)轉(zhuǎn)移19例。本研究經(jīng)山東省萊蕪市人民醫(yī)院倫理委員會審批通過。
1.2 主要試劑與儀器
人胰腺癌PANC-1細胞購于中國科學院細胞庫。胰酶、胎牛血清、RPMI 1640培養(yǎng)基購于美國Gibco公司,TROP2、cleaved caspase 3、NOTCH1、HES1抗體購于美國Abcam公司,CCK8細胞增殖試劑盒、BCA試劑盒、Annexin V-FITC/PI凋亡試劑盒均購于碧云天生物技術(shù)研究所,熒光定量試劑盒和反轉(zhuǎn)錄試劑盒購于日本TaKaRa公司,酶標儀購于美國Bio-Rad公司,流式細胞儀購于美國Becton Dickinson公司。
1.3 檢測方法
1.3.1TROP 2基因mRNA表達的檢測方法利用Trizol法提取胰腺癌和相應的癌旁組織中的總RNA。根據(jù)逆轉(zhuǎn)錄試劑盒的操作說明,將總RNA反轉(zhuǎn)錄為cDNA。利用Primer premier 6.0引物設(shè)計軟件設(shè)計基因TROP2及內(nèi)參基因GAPDH的逆轉(zhuǎn)錄-聚合酶鏈反應(reverse transcription-polymerase chain reaction,RT-PCR)引物,每個樣品設(shè)置6個重復孔,通過實時熒光定量PCR儀對TROP2和GAPDH基因進行擴增。引物序列如下:①TROP2引物序列,上游引物為5'-CCTCATCGCCGTCATCGT-3',下游引物為5'-CGGTTCCTTTCTCAACTCCC-3';②GAPDH引物序列,上游引物為5'-GAAGGTGAAGGTCGGAGTC-3',下游引物為5'-GAAGATGGTGATGGGATTTC-3'。PCR的擴增條件為:95℃下5 min;95 ℃下30 s;60 ℃下15 s,72 ℃下20 s,共40個循環(huán)。72℃延伸15 min,4℃保存。根據(jù)熒光定量PCR所得循環(huán)數(shù)(cycle threshold,Ct)值,利用2-△△Ct法計算TROP2的mRNA相對表達量。實驗重復3次。
1.3.2 細胞培養(yǎng)與轉(zhuǎn)染的方法人胰腺癌PANC-1細胞在37℃、5%CO2培養(yǎng)箱中用含有10%胎牛血清和青鏈霉素雙抗的RPMI 1640細胞培養(yǎng)基中傳代培養(yǎng),取生長對數(shù)期的細胞用于轉(zhuǎn)染。轉(zhuǎn)染前24 h,將傳代培養(yǎng)后的細胞接種于6孔細胞培養(yǎng)板中,每孔加入 2 ml的 PANC-1細胞(濃度為 1×106/孔)。PANC-1細胞生長融合度>80%時進行轉(zhuǎn)染,轉(zhuǎn)染按照美國Invitrogen公司Lipofectamine TM 2000轉(zhuǎn)染說明進行操作。轉(zhuǎn)染分為對照組(不經(jīng)任何處理)、陰性對照組(轉(zhuǎn)染NC-siRNA)和TROP2沉默組(轉(zhuǎn)染TROP2-siRNA)。
1.3.3 TROP 2、cleaved caspase 3、NOTCH 1、HES 1蛋白表達的檢測方法將對照組、陰性對照組和TROP2沉默組的細胞,轉(zhuǎn)染48 h,加入RAPI細胞裂解液,提取細胞中的蛋白。取少量的蛋白樣品,利用BCA試劑盒對蛋白進行定量分析。在蛋白樣品與上樣緩沖Buffer充分混勻后,煮沸變性10 min。取30 μg的變性蛋白,依次進行SDSPAGE分離、PVDF轉(zhuǎn)膜、5%的脫脂奶粉封閉,孵育TROP2、cleaved caspase 3、NOTCH1、HES1作為一抗(均1∶1000稀釋),4℃過夜,TBST洗滌;加入辣根過氧化物酶標記的羊抗鼠IgG作為二抗(1∶1000稀釋),37℃孵育2 h。增強化學發(fā)光法(enhanced chemiluminescence,ECL)顯色,顯影,定影。實驗重復3次。
1.3.4 細胞增殖的檢測方法將對照組、陰性對照組和TROP2沉默組轉(zhuǎn)染后的細胞,以1×104/ml的濃度接種于96孔細胞培養(yǎng)板中,37℃、5%CO2培養(yǎng)箱中培養(yǎng)48 h,每組設(shè)置6個復孔,收集細胞。每孔加入CCK8試劑10 μl,置于培養(yǎng)箱繼續(xù)孵育2 h,收集細胞。利用空白對照孔調(diào)零,選擇酶標儀上450 nm處測定各孔的吸光度(A值),重復3次。細胞的增殖率=轉(zhuǎn)染組細胞A值/對照組細胞A值×100%。
1.3.5 細胞凋亡的檢測方法采用Annexin V/PI雙染法檢測細胞凋亡。取對照組、陰性對照組和TROP2沉默組轉(zhuǎn)染后培養(yǎng)48 h的細胞,加入預冷的PBS洗滌細胞,再加入300 μl結(jié)合緩沖液懸浮細胞,細胞濃度調(diào)整為 1×106/ml,取 100 μl的細胞懸液至流式管中,加入Annexin V-FITC及PI各5 μl,混勻后室溫避光孵育15 min,反應管中再加入400 μl的PBS。1 h內(nèi)通過流式細胞儀檢測細胞的凋亡情況。實驗重復3次。
1.4 統(tǒng)計學方法
采用SPSS 21.0統(tǒng)計軟件分析數(shù)據(jù),計量資料以均數(shù)±標準差(±s)表示,兩組比較采用獨立樣本t檢驗,多組間比較采用單因素方差分析,多組間兩兩比較采用LSD-t檢驗。P<0.05為差異有統(tǒng)計學意義。
2 結(jié)果
2.1 胰腺癌組織中TROP 2基因mRNA表達
采用RT-PCR法檢測TROP2基因在胰腺癌組織和癌旁組織中的mRNA表達,TROP2基因在胰腺癌組織中的mRNA相對表達量為(5.987±0.583),明顯高于癌旁組織的(0.953±0.049),差異有統(tǒng)計學意義(P<0.01)。(圖1)
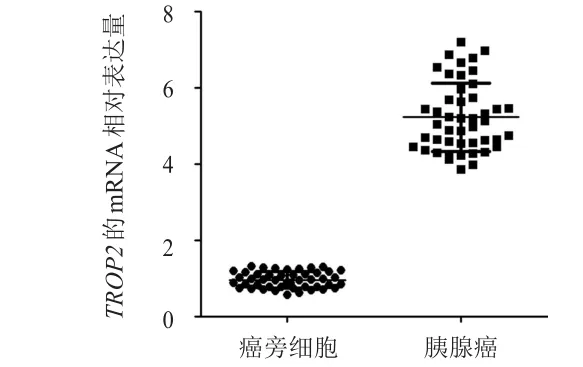
圖1 胰腺癌中TROP 2基因的mRNA表達
2.2 轉(zhuǎn)染TROP 2-siRNA后PANC- 1細胞中TROP 2蛋白表達
TROP2-siRNA轉(zhuǎn)染PANC-1細胞48 h后,采用蛋白質(zhì)印跡法檢測對照組、NC-siRNA組和TROP2-siRNA組TROP2蛋白表達,分別為(0.355±0.032)、(0.350±0.036)和(0.148±0.024),3組比較,差異有明顯統(tǒng)計學意義(F=43.342,P<0.01);TROP2-siRNA組TROP2蛋白表達明顯低于對照組,差異有統(tǒng)計學意義(P<0.01),而NC-siRNA組TROP2蛋白表達與對照組比較,差異無統(tǒng)計學意義(P>0.05)。(圖2)
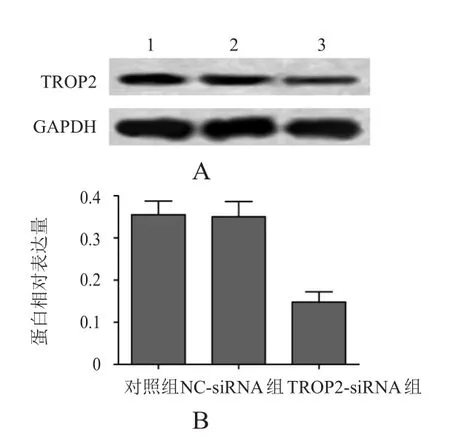
圖2 轉(zhuǎn)染TROP 2-siRNA后的PANC- 1細胞中TROP 2的蛋白表達
2.3 轉(zhuǎn)染TROP 2-siRNA降低PANC- 1細胞增殖
對照組、NC-siRNA組和TROP2-siRNA組細胞的平均存活率分別為(100±2.12)%、(98.88±3.23)%、(67.45±4.32)%,3組比較,差異有明顯統(tǒng)計學意義(F=91.484,P<0.01);TROP2-siRNA組細胞的存活率明顯低于對照組,差異有統(tǒng)計學意義(P<0.01),而NC-siRNA組細胞的存活率與對照組比較,差異無統(tǒng)計學意義(P>0.05)。(圖3)
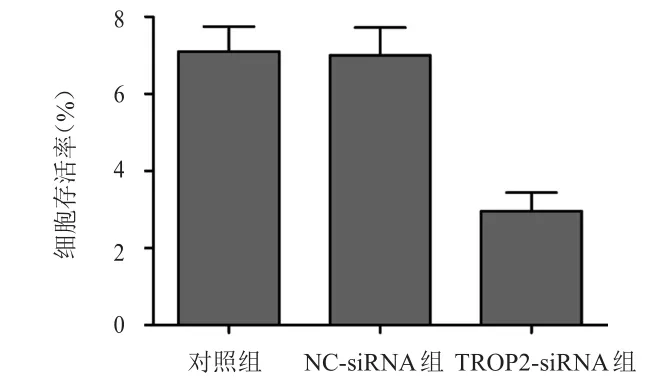
圖3 轉(zhuǎn)染TROP 2-siRNA對PANC- 1細胞增殖的影響
2.4 轉(zhuǎn)染TROP 2-siRNA促進PANC- 1細胞凋亡
對照組、NC-siRNA組和TROP2-siRNA組細胞的凋亡率分別為(1.83±0.54)%、(1.85±0.51)%和(17.21±1.36)%,3組比較,差異有明顯統(tǒng)計學意義(F=295.137,P<0.01);TROP2-siRNA組細胞的凋亡率明顯高于對照組,差異有統(tǒng)計學意義(P<0.01),而NC-siRNA組細胞的凋亡率與對照組比較,差異無統(tǒng)計學意義(P>0.05)。(圖4)
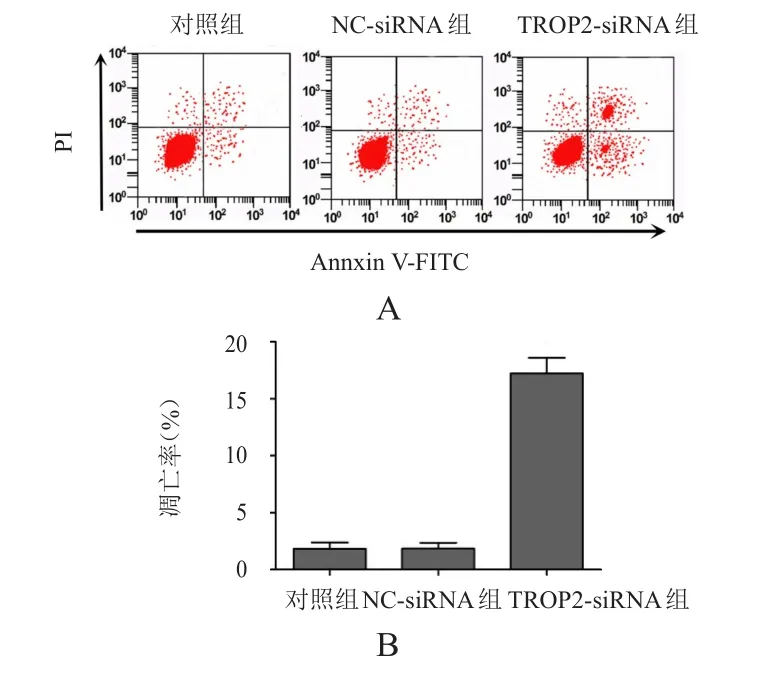
圖4 轉(zhuǎn)染TROP 2-siRNA對PANC- 1細胞凋亡的影響
2.5 轉(zhuǎn)染TROP 2-siRNA對PANC- 1細胞cleaved caspase 3、NOTCH 1、HES 1蛋白表達的影響
采用蛋白質(zhì)印跡法檢測凋亡相關(guān)蛋白cleaved caspase 3和NOTCH1信號通路相關(guān)蛋白NOTCH1、HES1的表達,對照組、NC-siRNA組和TROP2-siRNA組cleaved caspase 3蛋白表達分別為(0.03±0.01)、(0.03±0.01)和(0.13±0.02),NOTCH1的蛋白表達分別為(0.54±0.04)、(0.53±0.04)和(0.25±0.02),HES1的蛋白表達分別為(0.24±0.03)、(0.25±0.03)和(0.09±0.02),3組cleaved caspase 3、NOTCH1和HES1蛋白表達比較,差異均有明顯統(tǒng)計 學 意 義(F=47.45、64.58、44.84,P< 0.01);TROP2-siRNA組cleaved caspase 3蛋白表達明顯高于對照組,NOTCH1、HES1蛋白表達明顯低于對照組,差異均有統(tǒng)計學意義(P<0.01),而NC-siRNA組cleaved caspase 3、NOTCH1、HES1蛋白表達與對照組比較,差異均無統(tǒng)計學意義(P>0.05)。(圖5)
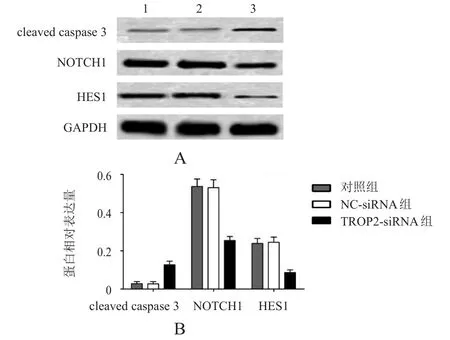
圖5 轉(zhuǎn)染TROP 2-siRNA對PANC- 1細胞cleavedcaspa se 3、NOTCH 1、HES 1蛋白表達的影響
3 討論
惡性腫瘤具有無限、快速的增殖能力,嚴重地威脅人類的生命健康,其發(fā)生的原因包括抑癌基因、原癌基因和DNA損傷修復基因等表觀遺傳學的改變或突變。目前,多種腫瘤分子標志物在腫瘤的發(fā)生過程中的作用受到人們廣泛地關(guān)注[8-9]。TROP2屬于TACSTD家族,又稱為EGP-1、TACSTD2、GA733-1,定位于人類染色體1p32,是一種單次跨膜表面的糖蛋白。近年,研究者通過基因表達串聯(lián)分析、基因芯片和免疫組化等方法,發(fā)現(xiàn)在多種惡性腫瘤中均有TROP2基因的高表達,其表達與遠處轉(zhuǎn)移及預后不良有密切的關(guān)系[10-13]。研究顯示,卵巢癌組織中TROP2基因高表達,其表達程度與腹腔積液轉(zhuǎn)移、淋巴結(jié)轉(zhuǎn)移及總生存期等有密切的關(guān)系[14-15]。宮頸癌組織中TROP2基因的高表達與臨床分期、淋巴結(jié)轉(zhuǎn)移、組織學分化程度及浸潤深度等有密切的關(guān)系[16-17]。這些研究結(jié)果說明,TROP2基因的高表達對惡性腫瘤的發(fā)生、浸潤、轉(zhuǎn)移和預后等有重要的影響;深入研究該基因在腫瘤中的生物學作用,尋找其作用機制,TROP2有望成為潛在的治療腫瘤的分子靶點。
研究顯示,抑制胃癌和前列腺癌組織中TROP2基因的表達,可以降低腫瘤細胞的侵襲和遷移能力[18-19];抑制肺腺癌組織中TROP2基因的表達,可以降低腫瘤細胞的增殖能力,促進細胞凋亡[20]。RNA干擾(RNA interference,RNAi)是由雙鏈RNA介導的可以使基因在轉(zhuǎn)錄后發(fā)生沉默的現(xiàn)象,有高度的特異性和有效性,是研究基因功能的有效途徑[21-22]。本研究通過RNA干擾技術(shù)沉默胰腺癌組織中TROP2基因的表達,檢測細胞的增殖和凋亡情況,結(jié)果顯示,抑制TROP2基因表達后細胞的增殖明顯降低,細胞凋亡增加。
細胞凋亡受到多種因素的調(diào)控,Caspase家族在細胞凋亡過程中發(fā)揮重要的作用。caspase 3是Caspase家族中的關(guān)鍵酶和效應蛋白,處在caspase級聯(lián)反應的下游,受到凋亡刺激后被激活,活化后可以使細胞凋亡進入不可逆階段[23],目前已在胰腺癌和胃癌等多種腫瘤細胞中得到證實[24-25]。NOTCH信號通路是一條進化保守途徑,參與細胞的增殖、凋亡、分化和機體發(fā)育等的調(diào)節(jié)過程,由NOTCH受體及配體、下游效應物、DNA結(jié)合蛋白組成,可以從多個方面對細胞的增殖、凋亡和機體發(fā)育等進行調(diào)控,影響細胞命運[26]。NOTCH1是其中的一個受體,其異常表達可以直接或間接地誘導腫瘤發(fā)生[27]。研究顯示,抑制胰腺癌和肺癌中NOTCH1信號通路,可以減緩腫瘤的發(fā)生、發(fā)展[28-29]。HES1是NOTCH1信號通路下游最重要的靶基因,HES1基因表達與否是NOTCH1信號通路是否被激活的標志[30]。本研究在抑制TROP2基因表達后檢測cleaved caspase 3、NOTCH1、HES1蛋白表達,顯示抑制TROP2基因表達后cleaved caspase 3蛋白表達明顯上調(diào),NOTCH1、HES1蛋白表達明顯下調(diào)。
綜上所述,抑制胰腺癌中TROP2基因的表達,可以阻斷NOTCH1信號通路,降低癌細胞的增殖能力,促進細胞凋亡。本研究為胰腺癌分子治療途徑的研究提供了一定的理論基礎(chǔ)。
[1]Waddell N,Pajic M,Patch AM,et al.Whole genomes redefine the mutational landscape of pancreatic cancer[J].Nature,2015,518(7540):495-501.
[2]Melo SA,Luecke LB,Kahlert C,et al.Glypican-1 identifies cancer exosomes and detects early pancreatic cancer[J].Nature,2015,523(7559):177-182.
[3]Guerra E,Trerotola M,Tripaldi R,et al.Trop-2 Induces tumor growth through AKT and determines sensitivity to AKT inhibitors[J].Clin Cancer Res,2016,22(16):4197-4205.
[4]Wang X,Long M,Dong K,et al.Chemotherapy agents-induced immunoresistance in lung cancer cells could be reversed by trop-2 inhibition in vitro and in vivo by interaction with MAPK signaling pathway[J].Cancer Biol Ther,2013,14(12):1123-1132.
[5]Trerotola M,Cantanelli P,Guerra E,et al.Upregulation of Trop-2 quantitatively stimulates human cancer growth[J].Oncogene,2013,32(2):222-233.
[6]Zhao P,Yu HZ,Cai JH.Clinical investigation of TROP-2 as an independent biomarker and potential therapeutic target in colon cancer[J].Mol Med Rep,2015,12(3):4364-4369.
[7]Lin H,Zhang H,Wang J,et al.A novel human Fab antibody for Trop2 inhibits breast cancer growth in vitro and in vivo[J].Int J Cancer,2014,134(5):1239-1249.
[8]Xia S,Lal B,Tung B,et al.Tumor microenvironment tenascin-C promotes glioblastoma invasion and negatively regulates tumor proliferation[J].Neuro Oncol,2016,18(4):507-517.
[9]Liu X,Chen L,Ge J,et al.The ubiquitin-like protein FAT10 stabilizes eEF1A1 expression to promote tumor proliferation in a complex manner[J].Cancer Res,2016,76(16):4897-4907.
[10]Guerra E,Trerotola M,Aloisi AL,et al.The Trop-2 signalling network in cancer growth[J].Oncogene,2013,32(12):1594-1600.
[11]Liu X,Li S,Yi F.Trop2 gene:a novel target for cervical cancer treatment[J].J Cancer Res Clin Oncol,2014,140(8):1331-1341.
[12]Gao XY,Zhu YH,Zhang LX,et al.siRNA targeting of Trop2 suppresses the proliferation and invasion of lung adenocarcinoma H460 cells[J].Exp Ther Med,2015,10(2):429-434.
[13]Chen MB,Wu HF,Zhan Y,et al.Prognostic value of TROP2 expression in patients with gallbladder cancer[J].Tumor Biol,2014,35(11):11565-11569.
[14]Goldenberg DM,Cardillo TM,Govindan SV,et al.Trop-2 is a novel target for solid cancer therapy with sacituzumab govitecan(IMMU-132),an antibody-drug conjugate(ADC)[J].Oncotarget,2015,6(26):22496-22512.
[15]Cardillo TM,Govindan SV,Sharkey RM,et al.Sacituzumab govitecan(IMMU-132),an anti-Trop-2/SN-38 antibody-drug conjugate:characterization and efficacy in pancreatic,gastric,and other cancers[J].Bioconjug Chem,2015,26(5):919-931.
[16]Liu T,Liu Y,Bao X,et al.Overexpression of TROP2 predicts poor prognosis of patients with cervical cancer and promotes the proliferation and invasion of cervical cancer cells by regulating ERK signaling pathway[J].PLoS One,2013,8(9):e75864.
[17]Wang XD,Wang Q,Chen XL,et al.Trop2 inhibition suppresses the proliferation and invasion of laryngeal carcinoma cells via the extracellular signal-regulated kinase/mitogen-activated protein kinase pathway[J].Mol Med Rep,2015,12(1):865-870.
[18]Zhao W,Zhu H,Zhang S,et al.Trop2 is overexpressed in gastric cancer and predicts poor prognosis[J].Oncotarget,2016,7(5):6136-6145.
[19]Trerotola M,Ganguly KK,Fazli L,et al.Trop-2 is up-regulated in invasive prostate cancer and displaces FAK from focal contacts[J].Oncotarget,2015,6(16):14318-14328.
[20]Li Z,Jiang X,Zhang W.TROP2 overexpression promotes proliferation and invasion of lung adenocarcinoma cells[J].Biochem Biophysical Res Commun,2016,470(1):197-204.
[21]Castel SE,Martienssen RA.RNA interference in the nucleus:roles for small RNAs in transcription,epigenetics and beyond[J].Nat Rev Genet,2013,14(2):100-112.
[22]Deng Y,Wang CC,Choy KW,et al.Therapeutic potentials of gene silencing by RNA interference:principles,challenges,and new strategies[J].Gene,2014,538(2):217-227.
[23]Cheng Q,Cao X,Yuan F,et al.Knockdown of WWP1 inhibits growth and induces apoptosis in hepatoma carcinoma cells through the activation of caspase3 and p53[J].Biochem Biophys Res Commun,2014,448(3):248-254.
[24]Arlt A,Sebens S,Krebs S,et al.Inhibition of the Nrf2 transcription factor by the alkaloid trigonelline renders pancreatic cancer cells more susceptible to apoptosis through decreased proteasomal gene expression and proteasome activity[J].Oncogene,2013,32(40):4825-4835.
[25]劉洪義,于金海,王大廣,等.Twist基因沉默促進人胃癌SGC7901細胞凋亡的機制[J].中國老年學雜志,2014,34(16):4618-4620.
[26]Hadland BK,Varnum-Finney B,Moon RT,et al.Notch signaling by either Notch1 or Notch2 mediates expansion of AGM-derived long-term HSC populations in vitro[J].Blood,2014,124(21):2897.
[27]Takebe N,Nguyen D,Yang SX.Targeting notch signaling pathway in cancer:clinical development advances and challenges[J].Pharmacol Ther,2014,141(2):140-149.
[28]Kunnimalaiyaan S,Trevino J,Tsai S,et al.Xanthohumolmediated suppression of Notch1 signaling is associated with antitumor activity in human pancreatic cancer cells[J].Mol Cancer Ther,2015,14(6):1395-1403.
[29]Baumgart A,Mazur PK,Anton M,et al.Opposing role of Notch1 and Notch2 in a Kras(G12D)-driven murine nonsmall cell lung cancer model[J].Oncogene,2015,34(5):578-588.
[30]Zhang K,Zhang YQ,Ai WB,et al.Hes1,an important gene for activation of hepatic stellate cells,is regulated by Notch1 and TGF-β/BMP signaling[J].World J Gastroenterol,2015,21(3):878-887.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
