CRH介導妊娠期慢性應激致子代雄鼠抑郁
2018-05-23 02:45:24呂逸麗韓振敏姚余有
安徽醫科大學學報 2018年4期
況 亮,呂逸麗,韓振敏,陳 鵬,姚余有,2,唐 偉
抑郁癥是一種常見的心境障礙,實驗研究[1]顯示,抑郁的發病與下丘腦、杏仁核、大腦皮層和海馬等部位異常有關。本課題組前期研究[2]顯示,妊娠期慢性應激可引起子代雄鼠呈現抑郁癥狀,但其機制不明。促腎上腺皮質激素釋放激素(corticotropin-releasing hormone,CRH)主要由下丘腦室旁核分泌,大腦皮層、杏仁核、海馬等腦組織也表達一定量的CRH[3]。以往研究[2,4]顯示高水平CRH可能與抑郁的發生有關,但CRH是否介導妊娠期慢性應激致抑郁尚不清楚。因此,該研究擬用妊娠期慢性不可預見性復合應激建立子代雄鼠抑郁模型,并用CRHR1拮抗劑(antalarmin)對子鼠進行干預,觀察子代雄鼠抑郁程度并進一步探討其作用機制。
1 材料與方法
1.1試劑與儀器antalarmin hydrochloride(美國Sigma公司);雷帕霉素靶蛋白(mammalian target of rapamycin, mTOR)單克隆抗體(美國Abcam公司);β-actin單克隆抗體、山羊抗兔二抗、化學發光檢測試劑盒(上海碧云天生物技術有限公司);小鼠CRH酶聯免疫試劑盒(武漢華美生物公司);電泳儀、凝膠成像分析系統(北京Tanon公司);強迫游泳圖像采集及分析系統(上海欣軟公司);曠場實驗圖像采集及分析系統(荷蘭Noldus公司)。
1.2實驗動物與分組8周齡SPF級小鼠,雌性40只,雄性10只,由安徽醫科大學實驗動物中心提供。遵循安徽醫科大學生物醫學倫理委員會動物飼養要求,室溫為(23±1)℃,攝食飲水自由,50%的相對濕度,飼養1周。按照雌雄比例2 ∶1在晚上19:00合籠,第2天早上07:00前檢查陰栓,查到陰栓的小鼠單獨飼養,并標記為受孕第1天,并將孕鼠按2 ∶1隨機分為慢性應激組和正常處理組。慢性應激組正常飼養1周,受孕第8天施加應激:束縛、禁水(24 h)、夾尾(5 min)、溫水游泳(30 ℃,15 min)、禁食(24 h)、改變居住環境(讓鼠在潮濕的墊料上生存24 h)、冰水游泳(4 ℃,5 min)等7種應激方式。小鼠每天隨機接受一種應激方法(禁水、禁食除外),直至小鼠分娩。正常處理組正常飼養即可。子代雄鼠母乳喂養21 d斷奶,隨機分為三組(n=10):正常對照組(Control組)、產前應激+溶劑對照組(CPS組)、產前應激+CRHR1拮抗劑組(CPS+ANT組),CPS+ANT組子代雄鼠于產后21~41 d每天腹腔注射微量CRHR1拮抗劑[5](20 μg/g),CPS組和Control組注射等量的生理鹽水。
1.3行為學實驗
1.3.1曠場實驗 每組隨機選取8只子代雄鼠于產后45 d進行曠場實驗,在實驗的過程中確保實驗室安靜,曠場實驗時將小鼠轉移至試驗箱的中央,于2 min后開始測試,記時3 min,記錄小鼠爬行總路程、平均速度、穿越象限次數、站立次數。每結束一次實驗,用酒精擦拭曠場實驗箱并清理小鼠的糞便和尿液,待酒精揮發完,再換下一只小鼠進行實驗。
1.3.2強迫游泳實驗 每組隨機選取8只子代雄鼠于產后47 d進行強迫游泳實驗,第1天將三組小鼠置于裝有25 ℃水的鋼化玻璃桶中游泳15 min(水深30 cm,直徑20 cm),一次實驗只能用一只鼠,以免小鼠互相影響。第2天,隨機將子代雄鼠放入裝有水的鋼化玻璃桶中(水深及溫度與前1 d相同),錄像、記錄并觀察小鼠6 min內后4 min小鼠在桶內禁止不動時間,即小鼠在水面漂浮不動或僅有四肢微小的動作維持身體平衡的時間。
1.3.3糖水偏好實驗 每組隨機選取4只子代雄鼠于產后50 d進行糖水偏好實驗,小鼠單籠飼養在安靜的鼠房內,每籠放置兩個水瓶(大小、形狀,顏色相同)。第一個24 h,兩瓶都放置相同重量1%的蔗糖水,使小鼠適應蔗糖的甜味;第二個24 h,讓小鼠適應兩種不同甜味的飲用水,其中一瓶裝有1%的蔗糖溶液,第二瓶裝有相同體積的飲用水;第三個24 h,禁食禁水24 h,隨后給予相同重量的蔗糖水和純水進行糖水偏好實驗,24 h后對兩水瓶進行稱重,計算糖水偏好率。
1.4HE染色觀察病理形態學改變所有行為學實驗結束后,隨機選取5只小鼠,眼球取血,頸椎脫臼處死,在冰上取腦。用4%多聚甲醛將取下來的腦組織固定,然后脫水、包埋,最后于前囪后3 mm作6 μm的冠狀切片。應用HE染色技術將每組相同位置的腦組織切片進行染色,每組取6張,在顯微鏡下觀察海馬CA3區神經元的形態變化,拍照保存。然后用Image-Pro Plus Image 軟件計算出神經元的數目并求出平均值。
1.5Westernblot法檢測海馬組織mTOR的表達量冰上取腦結束后,將取出的全腦在冰上進行海馬剝離,置于勻漿機中,提取總蛋白,應用BCA試劑盒測定海馬組織蛋白的總濃度并定量,變性10 min。灌制8%的膠,每孔上樣28 μg;電泳(60 V/50 min,120 V/300 min);轉膜(200 mA/300 min);室溫封閉1.5 h;洗膜3次;一抗4 ℃孵育24 h;洗膜3次,二抗室溫孵育1.5 h;洗膜,滴加顯影液,測量條帶的光密度值。所用抗體濃度:一抗 β-actin(兔單抗,1 ∶2 000)、mTOR(兔單抗,1 ∶1 000),二抗(羊抗兔,1 ∶2 000)。
1.6ELISA法檢測海馬CRH水平用研磨機將海馬研碎,按照說明書離心取海馬組織上清液并測量海馬組織CRH濃度。
2 結果
2.1CRHR1拮抗劑抑制妊娠期慢性應激致子代雄鼠抑郁與Control組相比,CPS組強迫游泳實驗中小鼠不動時間增多(P<0.05);糖水偏好實驗中糖水偏好率下降(P<0.05);曠場實驗中鼠爬行總路程、平均速度、穿越象限次數、站立次數減少(P<0.05);與CPS組相比,CPS+ANT組強迫游泳實驗中小鼠不動時間減少(P<0.05),糖水偏好率增高(P<0.05),曠場實驗中鼠爬行總路程、平均速度、穿越象限次數、站立次數增加(P<0.05)。見表1。
2.2CRHR1拮抗劑改善妊娠期慢性應激所致的子代雄鼠海馬CA3區神經元病理改變HE染色結果顯示,Control組小鼠海馬CA3區神經元染色清晰,數量相對較多,排列緊密,細胞核表現出圓形,核仁較明顯;CPS組神經元排列疏松,數量相對較少,脫失現象明顯,部分細胞核濃染、固縮。CPS+ANT組小鼠海馬CA3區神經元染色清晰,數量較多,呈椎體狀、圓球狀,細胞核呈圓形,核仁明顯。用Image-Pro PlusImage分析軟件計數海馬CA3區神經元總數,結果顯示與Control組(173.000±8.888)相比,CPS組(85.667±8.737)神經元總數明顯減少(P<0.05);與CPS組相比,CPS+ANT組(171.667±6.506)神經元總數明顯增多(P<0.05)。見圖1。
2.3CRHR1拮抗劑上調妊娠期慢性應激子代雄鼠海馬下降的mTOR的表達Western blot結果顯示,與Control組相比,CPS組小鼠海馬組織mTOR表達量減少(P<0.05);與CPS組相比,CPS+ANT組小鼠海馬組織的mTOR表達量增加(P<0.05);Control組與CPS+ANT組小鼠海馬組織mTOR表達量差異無統計學意義。見圖2。
2.4妊娠期慢性應激致子代雄鼠海馬CRH水平升高ELISA法檢測小鼠海馬CRH結果顯示,3組子代雄鼠海馬CRH濃度差異有統計學意義,與Control組相比,CPS組子代雄鼠海馬CRH水平升高(P<0.05);與CPS組相比,CPS+ANT組小鼠海馬CRH水平降低(P<0.05),見圖3。
3 討論
CRH神經元處于HPA軸頂端、具有整合心理和物理刺激的功能,被公認為是應激反應的中樞驅動力[6]。CRH主要由下丘腦室旁核分泌,大腦皮層、海馬、杏仁核等腦組織也表達一定量的CRH[7]。本研究結果顯示,CPS組較Control組海馬組織CRH濃度上升,說明妊娠期慢性應激可致子代海馬組織CRH升高。

表1 CRHR1拮抗劑抑制妊娠期慢性應激致子代雄鼠抑郁
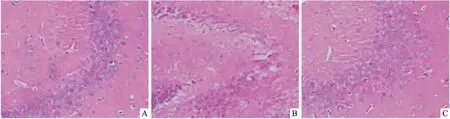
圖1 CRHR1拮抗劑改善妊娠期慢性應激所致的子代雄鼠海馬CA3區神經元 HE×40
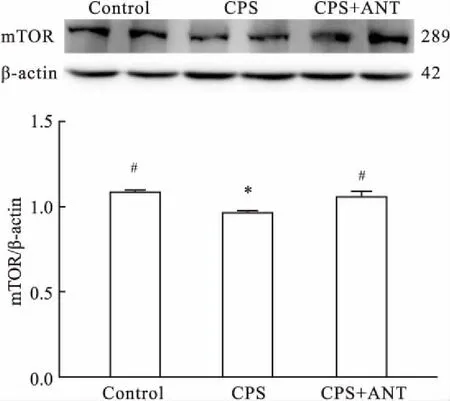
圖2 CRHR1拮抗劑上調妊娠期慢性應激子代雄鼠海馬下降的mTOR的表達量
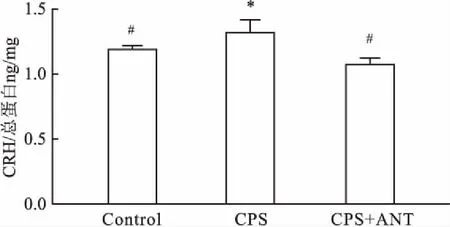
圖3 妊娠期慢性應激致子代雄鼠海馬CRH水平升高
強迫游泳實驗、糖水偏好實驗和曠場實驗是目前測量小鼠抑郁程度的常用實驗。實驗很好的分析了小鼠的無助程度、對快感缺乏的程度和在新環境中的探索行為。本研究行為學實驗顯示CPS組子代雄鼠較Control組小鼠靜止漂浮時間增多;糖水偏好實驗中糖水偏好率下降;曠場實驗中鼠爬行總路程、平均速度、穿越象限次數、站立次數減少。CPS+ANT組較CPS組小鼠靜止漂浮時間減少,糖水偏好率上升,曠場實驗中鼠爬行總路程、平均速度、穿越象限次數、站立次數增多。說明妊娠期慢性應激導致子代雄鼠行為學改變,使子代雄鼠產生抑郁樣行為,CRHR1拮抗劑可逆轉妊娠期慢性應激致子代雄鼠抑郁,提示CRH介導了妊娠期慢性應激致子代雄鼠抑郁。
實驗研究[8-9]顯示,海馬部位異常與抑郁的發病有關,其中CA3區與抑郁的形成密切相關。因此該區神經元數目、形態結構和功能的改變必然會導致行為學的改變。本實驗的海馬CA3區HE染色結果顯示,CPS組較Control組海馬CA3區神經元數量明顯減少,并且排列疏松紊亂,異常神經元明顯增多,部分細胞核濃染、固縮,而CPS+ANT組較CPS組海馬CA3區神經元染色清晰,數量較多,核濃染、固縮減少。說明CRHR1拮抗劑逆轉妊娠期慢性應激致子代雄鼠海馬CA3區神經元損傷,提示CRH致海馬CA3區神經元損傷可能是妊娠期慢性應激致子代雄鼠抑郁的原因。
mTOR信號通路是調控細胞生長與增殖的一個關鍵通路,該通路將從營養分子、能量狀態以及生長因子傳來的信號整合在一起,從而調控生命過程。mTOR分為mTORC1 和mTORC2,mTORC1信號在調節神經元的存活和分化、樹突的長度和分支、軸突的生長和突觸的可塑性中發揮重要作用,同時也介導抑郁的發病[10-11]。本研究顯示,CPS組較Control組海馬mTOR明顯下降,CPS+ANT組較CPS組海馬mTOR明顯上升并接近Control組水平。說明CRH受體拮抗劑可上調mTOR水平。提示妊娠期慢性應激通過升高的CRH下調mTOR的表達進而致子代雄鼠抑郁。
本研究結果還顯示, CPS+ANT組較CPS組海馬組織CRH濃度下降,提示CRHR1拮抗劑可以降低海馬CRH水平,其機制是否與CRHR1拮抗劑下調糖皮質激素濃度有關尚不清楚,有待于進一步研究。
綜上所述,CRH介導了妊娠期慢性應激致子代抑郁[12-13],其機制可能與妊娠期慢性應激致子代海馬CRH升高,下調mTOR蛋白表達,進而引起海馬 CA3 區神經元損傷有關。
參考文獻
[1] Duman R S, Aghajanian G K. Synaptic dysfunction in depression: potential therapeutic targets.[J]Science, 2012, 338(6103): 68-72.
[2] 韓振敏, 況 亮, 王正玉, 等. 妊娠期慢性應激對不同性別子代抑郁的影響[J]. 安徽醫科大學學報, 2017, 52(7): 998-1002.
[3] Ivy A S, Rex C S, Chen Y, et al. Hippocampal dysfunction and cognitive impairments provoked by chronic early-life stress involve excessive activation of CRH receptors[J]. J Neurosci, 2010, 30(39): 13005-15.
[4] Brunton P J. Effects of maternal exposure to social stress during pregnancy: consequences for mother and offspring[J]. Reproduction, 2013, 146(5): R175-89
[5] Liu R, Yang X D, Liao X M, et al. Early postnatal stress suppresses the developmental trajectory of hippocampal pyramidal neurons: the role of CRHR1[J]. Brain Struct Funct, 2016, 221(9): 4525-36.
[6] Galesi F L, Ayanwuyi L O, Mijares M G, et al. Role of Hypothalamic-Pituitary-Adrenal axis and corticotropin-releasing factor stress system on cue-induced relapse to alcohol seeking[J]. Eur J Pharmacol, 2016, 788: 84-9.
[7] Maras P M, Baram T Z. Sculpting the hippocampus from within: stress, spines, and CRH[J]. Trends Neurosci, 2012, 35(5): 315-24.
[8] Morales-Medina J C, Iannitti T, Freeman A, et al. The olfactory bulbectomized rat as a model of depression: The hippocampal pathway[J]. Behav Brain Res, 2017, 317: 562-75.
[9] Gilbert P E, Brushfield A M. The role of the CA3 hippocampal subregion in spatial memory: a process oriented behavioral assessment[J]. Prog Neuropsychopharmacol Biol Psychiatry, 2009, 33(5): 774-81.
[10] Child N D, Benarroch E E. mTOR: its role in the nervous system and involvement in neurologic disease[J]. Neurology, 2014, 83(17): 1562-72.
[11] Li N, Lee B, Liu R J, et al. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists[J]. Science, 2010, 329(5994): 959-64.
[12] Chen Y, Dube C M, Rice C J, et al. Rapid loss of dendritic spines after stress involves derangement of spine dynamics by corticotropin-releasing hormone[J]. J Neurosci, 2008, 28(11): 2903-11.
[13] Erhardt A, Muller M B, Rodel A, et al. Consequences of chronic social stress on behaviour and vasopressin gene expression in the PVN of DBA/2OlaHsd mice-influence of treatment with the CRHR1-antagonist R121919/NBI 30775[J]. J Psychopharmacol, 2009, 23(1): 31-9.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
