云南蝙蝠攜帶新型漢坦病毒及其基因組分析
2018-05-26 04:24:39鄧洪巖王靜林李利利信云云劉蒙蒙王云段招軍
中華實驗和臨床病毒學雜志 2018年2期
關鍵詞:分析
鄧洪巖 王靜林 李利利 信云云 劉蒙蒙 王云 段招軍
266021青島大學基礎醫學院病原生物系病原生物學專業(鄧洪巖、劉蒙蒙、王云);650224 昆明,云南省畜牧獸醫科學院/云南省熱帶亞熱帶動物病毒重點實驗室(王靜林);100051 北京,中國疾病預防控制中心病毒病預防控制所(李利利、段招軍);410003 長沙,湖南師范大學附屬第一醫院兒童醫學中心(信云云)
由于漢坦病毒(hantavirus, HV)的多樣性,2016 年國際病毒學分類委員會(ICTV)規定 HV 屬于布尼亞病毒目(bunyavirales)漢坦病毒科(hantaviridae)漢坦病毒屬(genus hantavirus),是有包膜分節段的單股負鏈 RNA 病毒。HV 基因組由大(L)、中(M)、小(S)3個節段組成,每個基因節段都只有一個開放閱讀碼框(open reading frame, ORF),L 節段編碼RNA依賴的 RNA 多聚酶(RdRp),M 節段編碼糖蛋白(G1、G2)的前體糖蛋白,S 節段編碼核衣殼蛋白[1]。人類感染HV后主要導致兩種嚴重疾病,即腎綜合征出血熱(hemorrhagic fever with renal syndrome, HFRS)和漢坦病毒肺綜合征(hantavirus pulmonary syndrome, HPS)[2]。每年全世界發生的 HFRS 病例中有超過 90% 發生在亞洲,其中多數感染病例分布于中國的29個省份和地區[3-5]。近幾年HV的研究進展較快,新型或新亞型病毒不斷被發現,在證實的40多個 HV 血清型/基因型中有7個型的 HV 可引起 HFRS,15個型引起 HPS,其余型別的 HV 與人類疾病的關系尚不太清楚[3]。
HV 的天然宿主是嚙齒類動物[3-4, 6];但最近發現蝙蝠和食蟲目動物也攜帶多種 HV[7-9]。蝙蝠是多種新發現的高致病性病毒的重要儲存庫,比如,狂犬病毒、尼帕病毒和SARS樣冠狀病毒,由于蝙蝠具有長距離飛行的能力和冬眠的特性,使其具有傳播病原體范圍較廣且能夠攜帶多種病毒的特點[10],因此對于蝙蝠體內新型病毒的研究具有重要意義。本研究利用病毒宏基因組學方法在云南普洱地區蝙蝠脾臟發現了HV的存在,同時我們對該病毒進行了篩查、全基因組擴增、相似性和進化分析。
1 材料與方法
1.1標本來源和處理本實驗中的標本采集通過了中國疾控中心病毒病預防控制所動物實驗倫理委員會的審查(編號:20160715023)。于2016年7~8月份在云南普洱地區隨機捕捉 84 只蝙蝠,經低溫保存并運送至本實驗室,在二級生物安全柜中解剖肝、脾、肺、腸道和肌肉組織,每種組織取黃豆粒大小放入 2 ml 離心管并加入 1 ml DMEM,其中肌肉組織用于鑒定蝙蝠種類,組織研磨后按照本實驗室建立的高通量測序前標本處理方法進行樣本過濾、混合、消化、提取核酸并擴增后進行高通量測序。
1.2所用試劑核酸提取試劑盒Viral Nucleic Acid Extraction Kit Ⅱ購于 Geneaid 公司;SuperscriptⅢ反轉錄酶,RNA 酶抑制劑,dNTP mix 購于 Invitrogen 公司;I-5TM2×High-Fidelity Master Mix 購自于北京擎科新業生物技術有限公司;Random Primers 購于 Promega 公司;引物合成及 PCR 產物測序均委托于北京擎科新業生物技術有限公司。
1.3病毒核酸提取及cDNA的合成吸取上述蝙蝠組織研磨液上清 200 μl,按照 Viral Nucleic Acid Extraction Kit Ⅱ 說明書提取病毒核酸,利用 SuperscriptⅢ 進行逆轉錄合成 cDNA。
1.4病毒基因組的擴增對注釋到 HV 的高通量測序讀長拼接后設計引物進行 PCR 驗證,根據獲得的序列設計引物擴增未知序列區域,根據漢坦病毒3‘和 5′端保守區序列設計引物擴增兩端序列。擴增產物經過 1.2% 瓊脂凝膠電泳鑒定并送北京擎科新業生物技術有限公司測序。
1.5標本篩查和蝙蝠種類鑒定根據擴增的全長基因組設計篩查引物,對收集的 84 只蝙蝠的肺、脾共 168 份標本中的 DodeHV 流行情況進行篩查。蝙蝠種類的鑒定使用引物PCR方法對肌肉線粒體細胞色素 B 基因進行擴增[11]。
1.6非同義突變替換率/同義突變替換率(dN/dS)比率分析dN/dS 比值使用在線軟件 DataMokeny[12]與同源性最高的蝙蝠 HV(XSV)3個節段分別進行計算分析。
1.7系統進化和同源性分析下載已知的蝙蝠 HV 及其他嚙齒類動物、鼴鼠和人群中發現的相關 HV 的 L、M、S 基因序列和M節段的氨基酸序列,將 DodeHV與上述下載的序列利用 MEGA6.0 軟件分別進行多序列比對并采用鄰接法(neighbor-joining)建立系統發育樹,將 bootstrap 值設定為1,000;用 MegAlign 進行同源性分析。
2 結果
2.1蝙蝠種類本研究中的 84 只蝙蝠經過特異性 PCR 鑒定,結果顯示 17 只為中華菊頭蝠、67 只為小蹄蝠。
2.2DodeHV的發現與篩查對蝙蝠高通量測序數據進行生物信息學分析,發現在脾臟組織中有27條讀長(Reads)注釋到 HV,根據拼接后的序列設計引物進行驗證和篩查,發現有 4 只小蹄蝠的肺臟和脾臟顯示 HV 陽性,陽性率為 5.97%(4/67),將該 HV 命名為 DodeHV。檢出 DodeHV 陽性的蝙蝠均為來自云南普洱地區的小蹄蝠(H. pomona andersen)。
2.3DodeHV的基因組本研究獲得的DodeHV 3個節段的長度分別為 S 1,702 bp、M 2,994 bp、L 6 521b,序列分析發現:S節段包括完整的 1 284 bp ORF 區以及 57 bp 完整的 5′ 端非編碼區和 361 bp 不完整的 3′ 端非編碼區,編碼 427 個氨基酸的核衣殼蛋白;M 節段是部分核苷酸序列;L 節段獲得了完整的序列,ORF 區位于nt40~6477 的位置,5′ 和 3′ 端包含 9 bp 高度保守的的反向互補序列(TAGTAGTAG),可使 5′ 和 3′ 形成穩定的鍋柄狀結構,這是布尼亞病毒科基因組的典型特征。S、M、L 的基因序列在 GenBank 中的登錄號分別為:MG637438、MG637437、MG637436。
2.4DodeHV序列相似性分析序列分析結果顯示 DodeHV 3個節段與 2013 年越南富壽省小蹄蝠體內發現的 Xuan son virus(XSV)[13]各基因節段相似度最高,S、M、L節段核苷酸序列和氨基酸序列相似度分別為: 79.0%和 93.4%, 79.2%和 94.8%,79.9%和 96.6%。DodeHV 與2015 年在廣西來賓市蝙蝠中發現的 LBV[14]相似度相對較小,和在中國湖北蝙蝠中發現的兩種病毒 HUPV 和 LQUV 基因序列相似度更低(表1)。我們同時將本次四份陽性標本的 M 節段核苷酸部分編碼區序列進行了同源性分析,發現它們之間沒有差別。DodeHV 與 XSV 的 dN/dS 比值分別為 S 0.00420、M 0.00736、L 0.000247。
2.5遺傳進化分析S、M、L 3個節段核苷酸序列進化分析發現漢坦病毒均可以分成了 4 個簇(Ⅰ-Ⅳ),而且顯示 DodeHV 在進化關系中均與越南富壽省報道的 XSV最接近,這與基因序列相似性分析的結果相一致,提示DodeHV 與 XSV 來自共同的祖先(圖1、2、3)。
表 1 DodeHV 與 GenBank 中已有蝙蝠漢坦病毒核苷酸序列和氨基酸序列的相似度(%)比較
Tab.1Nucleotide and amino acid sequence of bat hantaviruses from GenBank and their identity (%) with DodeHV
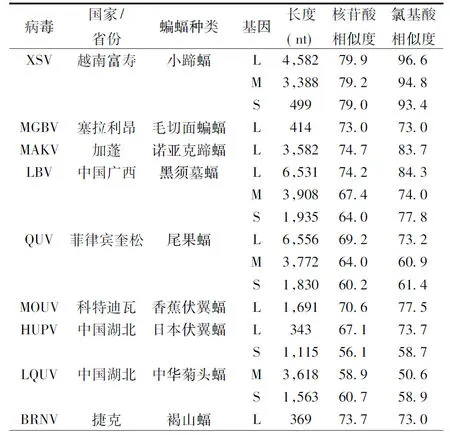
注:病毒名稱縮寫與 GenBank編號:
Note: Abbreviation and accession No XSV, Xuan son virus-VN1982B4 (JX912953, KU976427, KC688335); MGBV, Magboi virus 1209 (JN037851); MAKV, Makokou virus GB303 (KT316176); LBV, Laibin virus BT20 (KM102249, KM102248, KM102247); QUV, Queaon virus MT1720/1657 (KU950715, KU950714, KU950713); MOUV, Mouyassue virus (JQ287716); HUPV, Huangpi virus Pa-1 (JX465369, JX473273); LQUV, Lonquan virus Ra-25 (JX456397, JX465415); BRNV, Brno virus (KR920360)
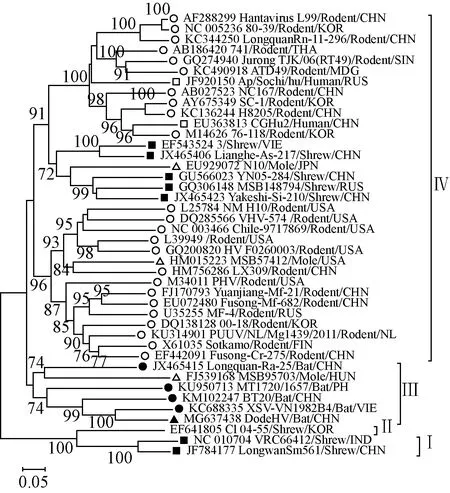
注:▲是本研究中的 DodeHV,●是已報道的蝙蝠體內發現的 HV,■是宿主為嚙齒類動物的HV,□是宿主為人類的HV,■是宿主為鼩鼱的HV,△是宿主為鼴鼠的HV圖1 DodeHV S 節段部分核苷酸序列(位置 731-1,230)系統進化樹Note:▲indicates DodeHV of this study, ●indicates bat-borne HV already found, ■indicates rodent-borne HV, □indicates HV found in human, ■indicates shrew-borne HV, △indicates mole-borne HVFig.1 Phylogenetic tree for partial nucleotide sequence of DodeHV S segment (positions 731-1,230)
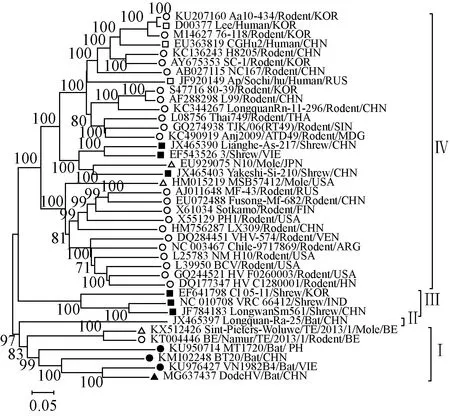
注:▲是本研究中的 DodeHV,●是已報道的蝙蝠體內發現的 HV,■是宿主為嚙齒類動物的HV,□是宿主為人類的HV,■是宿主為鼩鼱的HV,△是宿主為鼴鼠的HV圖2 DodeHV M 節段部分核苷酸序列(位置 300-2,810)系統進化樹Note:▲indicates DodeHV of this study, ●indicates bat-borne HV already found, ■indicates rodent-borne HV, □indicates HV found in human, ■indicates shrew-borne HV, △indicates mole-borne HVFig.2 Phylogenetic tree for partial nucleotide sequence of DodeHV M segment (positions 300-2,810)

注:▲是本研究中的 DodeHV,●是已報道的蝙蝠體內發現的 HV,■是宿主為嚙齒類動物的HV,□是宿主為人類的HV,■是宿主為鼩鼱的HV,△是宿主為鼴鼠的HV圖3 DodeHV L 節段部分核苷酸序列(位置 2,960-3,274)系統進化樹Note:▲indicates DodeHV of this study, ●indicates bat-borne HV already found, ■indicates rodent-borne HV, □indicates HV found in human, ■indicates shrew-borne HV, △indicates mole-borne HVFig.3 Phylogenetic tree for partial nucleotide sequence of DodeHV L segment (positions 2,960-3,274)
3 討論
自首次在黑線姬鼠被分離以來[15],漢坦病毒相繼在不同的宿主體內被分離或發現,如蝙蝠和食蟲目動物等[7-9]。蝙蝠具有攜帶并傳播多種人類病毒的優勢[10]。至今,蝙蝠攜帶的HV也不斷被報道,近3年時間就發現了4例,包括:中國廣西的 LBV[15]、非洲加蓬的 MAKV[16]、菲律賓奎松的 QUV[17]、歐洲捷克的 BRNV。蝙蝠攜帶HV的研究對人類動物性病毒病的預防和控制具有重要的公共衛生意義。
ICTV 第九次會議認為HV的分型標準之(https://talk.ictvonline.org/ictv-reports/ictv_9th_report):病毒和其他HV N蛋白氨基酸序列和G蛋白氨基酸序列相比至少有 7%的不同。DodeHV與XSV N和G蛋白氨基酸序列相似度分別為93.4%和94.8%。本研究DodeHV M序列不完整,但根據G蛋白534個氨基酸序列進化分析發現與M節段核酸序列進化分析結果一致。DodeHV 與越南 XSV 屬于HV同一基因型中不同的病毒株,這在我國境內屬于首次報道。
云南是多種病毒的疫源地[18-20],與越南、緬甸等東南亞國家接壤,在地理位置上具有重要的連通作用。本研究標本采集的普洱地區臨近邊境,對于充分理解邊境地區蝙蝠攜帶病毒的多樣性及東南亞國家蝙蝠病毒的系統進化關系具有重要意義。序列和進化分析顯示 DodeHV 與越南富壽省發現的 XSV 親緣關系最近,提示它們可能來源于同一祖先。但是 DodeHV 與廣西LBV、湖北龍泉 LQUV 以及湖北黃陂HUPV 進化關系相對較遠,且同源性較低,暗示漢坦病毒多樣性可能是通過病毒在種群間或個體間不斷傳播形成的,而本研究云南株與廣西和湖北流行株的差異較大可能是由于蝙蝠飛行距離的限制,導致了地理隔離。
dN(dS)表示每一個非同義(同義)位點上的平均非同義(同義)替換率,是描述同義替換和非同義替換的一個參數。DodeHV 與 XSV 的各節段 dN/dS 比率分別為 S 0.00420、M0.00736、L 0.000247,說明這3個節段都存在較多的同義突變,同時可以解釋該病毒與已知病毒核酸序列和氨基酸序列相似性差異較大的原因。
據報道 HV 與嚙齒類動物會具有共進化的關系,HV 在同一類宿主內傳播時,不會發生明顯的基因突變,但是當 HV 感染其他種類宿主時,病毒為了適應新宿主的遺傳環境,它會伴隨宿主而進化[21],本研究發現的DodeHV與同源性最近的越南XSV宿主都是小蹄蝠,而與其他蝙蝠種類所攜帶的漢坦病毒在進化關系中相差較遠,進一步提示了 HV 與宿主蝙蝠可能存在共進化關系,這可能也是 HV 在宿主轉換過程中可能產生病毒遺傳多樣性的原因。
總之,本研究發現的 DodeHV 不僅豐富了我國蝙蝠的病毒庫,而且促進了對漢坦病毒進化的理解。加大在全國各地對蝙蝠HV的監測與研究力度,充分掌握我國各地不同基因型HV的分布情況,才可以對新發暴發HV疫情做出及時有效的判斷,以最大限度挽救人們的生命財產。世界范圍內,不同種類蝙蝠體內不斷被發現各種新的病毒也提示我們應該更加深入研究蝙蝠 HV 的傳播對人類以及其他動物造成的影響。
利益沖突:無。
參考文獻
[1] Mir MA, Panganiban AT. The Hantavirus Nucleocapsid Protein Recognizes Specific Features of the Viral RNA Panhandle and Is Altered in Conformation upon RNA Binding[J]. J Virol, 2005,79(3):1824-1835. DOI:10.1128/JVI.79.3.1824-1835.2005.
[2] Schmaljohn CS, Hasty SE, Dalrymple JM, et al. Antigenic and genetic properties of viruses linked to hemorrhagic fever with renal syndrome[J]. Science, 1985,227(4690):1041.
[3] Jonsson CB, Figueiredo LTM, Vapalahti O. A Global Perspective on Hantavirus Ecology, Epidemiology, and Disease[J]. Clin Microbiol Rev, 2010,23(2):412-441. DOI: 10.1128/CMR.00062-09.
[4] Zhang YZ, Yang Z, Fu ZF, et al. Hantavirus Infections in Humans and Animals, China[J]. Emerg Infect Dis, 2010,16(8):1195. DOI: 10.3201/eid1608.090470.
[5] Kariwa H, Yoshimatsu K, Arikawa J. Hantavirus infection in East Asia[J]. Comp Immunol Microbiol Infect Dis, 2007,
30(5-6):341. DOI: 10.1016/j.cimid.2007.05.011.
[6] Schmaljohn C S, T. N S. Bunyaviridae[M]//Knipe D M H P. Fields Virology, fifth ed. Lippincott, Williams & Wilkins, Philadelphia: Morgan Kaufmann, 2007:1741-1789.
[7] Zhang YZ. Discovery of hantaviruses in bats and insectivores and the evolution of the genus Hantavirus[J]. Virus Res, 2014,187:15. DOI: 10.1016/j.virusres.2013.12.035.
[8] Guo WP, Lin XD, Wang W, et al. Phylogeny and Origins of Hantaviruses Harbored by Bats, Insectivores, and Rodents[J]. Plos Pathog, 2013,9(2):e1003159. DOI: 10.1371/journal.ppat.1003159.
[9] Sabrina W, Witkowski PT, Brita A, et al. Hantavirus in Bat, Sierra Leone[J]. Emerg Infect Dis, 2012,18(1):159-161. DOI: 10.3201/eid1801.111026.
[10] Calisher CH, Childs JE, Field HE, et al. Bats: Important Reservoir Hosts of Emerging Viruses[J]. Clin Microbiol Rev, 2006,19(3):531-545. DOI: 10.1128/CMR.00017-06.
[11] 何彪. 蝙蝠病毒組學及其新病毒的發現與鑒定[D]. 中國人民解放軍軍事醫學科學院, 2014.
[12] Delport W, Poon AFY, Frost SDW, et al. Datamonkey 2010: a suite of phylogenetic analysis tools for evolutionary biology[J]. Bioinformatics, 2010,26(19):2455. DOI: 10.1093/bioinformatics/btq429.
[13] Arai S, Nguyen ST, Boldgiv B, et al. Novel bat-borne hantavirus, Vietnam.[J]. Emerg Infect Dis, 2013,19(7):1159-1161. DOI: 10.3201/eid1907.121549.
[14] Xu L, Wu J, He B, et al. Novel hantavirus identified in black-bearded tomb bats, China[J]. Infect Genet Evol, 2015,31:158-160. DOI: 10.1016/j.meegid.2015.01.018.
[15] Lee HW, Lee PW, Johnson KM. Isolation of the etiologic agent of Korean Hemorrhagic fever.[J]. J Infect Dis, 1978,137(3):298.
[16] Witkowski PT, Drexler JF, Kallies R, et al. Phylogenetic analysis of a newfound bat-borne hantavirus supports a laurasiatherian host association for ancestral mammalian hantaviruses.[J]. Infect Genet Evol, 2016,41:113-119. DOI: 10.1016/j.meegid.2016.03.036.
[17] Arai S, Taniguchi S, Aoki K, et al. Molecular phylogeny of a genetically divergent hantavirus harbored by the Geoffroy′s rousette (Rousettus amplexicaudatus), a frugivorous bat species in the Philippines.[J]. Infect Genet Evol, 2016,45:26-32. DOI: 10.1016/j.meegid.2016. 08.008.
[18] 杜春紅, 趙文紅, 劉正祥, 等. 2012年云南省德欽縣鼠疫疫源地調查實驗室檢測結果與分析[J]. 疾病監測, 2013,28(11):917-919.
[19] 張玉芬, 杜春紅, 蘇超, 等. 云南省貢山縣鼠疫指示動物血清流行病學調查[J]. 疾病監測, 2015,30(11):922-924.
[20] Wang B, Feng Y, Li Z, et al. Distribution and diversity of hepatitis B virus genotypes in Yunnan, China.[J]. J Med Virol, 2014,86(10):1675. DOI: 10.1002/jmv.24002.
[21] Plyusnin A, Morzunov S P. Virus evolution and genetic diversity of hantaviruses and their rodent hosts.[J]. Curr Top Microbiol Immunol, 2001,256:47-75.
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
