羅氏沼蝦致病性溶藻弧菌的鑒定及藥敏分析
2018-06-04 02:08:45苗鵬飛譚淑雯陳言峰彭鐘琴吳勇亮
水產科學 2018年3期
苗鵬飛,楊 映,譚淑雯,陳言峰,彭鐘琴,吳勇亮,于 輝
( 佛山科學技術學院 生命科學與工程學院,廣東 佛山 528231 )
羅氏沼蝦(Macrobrachiumrosenbergii)亦稱馬來西亞大蝦、金錢蝦、淡水長臂大蝦、萬氏對蝦等,屬長臂蝦科、沼蝦屬,原產地在厄瓜多爾沿岸,是世界上養殖量最高的三大對蝦之一[1]。該品種具有生長快、食性廣、肉質營養成分好、養殖周期短等優點,是深受養殖戶喜愛的養殖品種。自東南亞地區引進我國后,經過30多年的養殖,羅氏沼蝦已經成為我國重要的淡水蝦類,養殖廣泛分布在江蘇、廣東、上海、福建、廣西、浙江等16個省市,養殖面積約3.34×104hm2,年產量逾1.2×105t[2]。隨著羅氏沼蝦養殖業不斷發展,高密度、高換水率的養殖模式嚴重影響周邊的水體環境,破壞了水體生態的平衡,繼而導致羅氏沼蝦的病害發生[3-4]。細菌是導致羅氏沼蝦病害發生的一個主要因素[5],弧菌(Vibrio)、氣單胞菌(Aeromonas)和假單胞菌(Pseudomonas)被認為是羅氏沼蝦感染細菌性疾病的主要原因[6]。筆者從廣東省湛江市某養殖場患病羅氏沼蝦體內分離到一株優勢菌QY170324,對其進行細菌形態觀察、生理生化特征鑒定、16S rDNA序列分析、藥敏試驗,旨在為羅氏沼蝦疾病研究提供理論依據,為羅氏沼蝦健康養殖提供一些參考資料。
1 材料與方法
1.1 試驗材料
患病幼蝦采自廣東省湛江市某羅氏沼蝦養殖場。癥狀為患病幼蝦行動遲緩、大部分浮于水面、紅體等。健康的幼蝦取自廣東省佛山市某養殖場。
硫代硫酸鹽—檸檬酸鹽—膽鹽—蔗糖弧菌選擇性培養基、營養肉湯、營養瓊脂、堿性蛋白胨水購于廣東環凱微生物科技有限公司;SuperMix、GelStain核酸染料(1000×)、瓊脂糖購于北京全式金生物技術有限公司;DNA Maker DL2000購于大連寶生物工程公司;細菌DNA提取試劑盒購于北京艾萊德生物科技有限公司;1%NaCl葡萄糖產氣、1%NaCl葡萄糖磷酸鹽凍水(V-P)、1%NaCl蛋白胨水、1%NaCl蔗糖、1%NaCl甘露糖、1%NaCl阿拉伯糖、1%NaCl肌醇、1%NaCl賴氨酸脫羧酶、1%NaCl精氨酸雙水解酶、無鹽胨水、3%NaCl胨水、6%NaCl胨水、8%NaCl胨水、10%NaCl胨水14種生化鑒定管購于廣東環凱微生物科技有限公司。
1.2 病原菌的分離、純化和保存
患病幼蝦于無菌狀態下勻漿處理后,用接種劃線針接種于硫代硫酸鹽—檸檬酸鹽—膽鹽—蔗糖培養基上,28 ℃培養24 h;挑取形態一致、單個優勢菌落接種于含3%NaCl的營養肉湯中,28 ℃,200 r/min搖床培養18 h。菌液中加入體積系數為30%的甘油,置于-80 ℃冰箱中保存。
1.3 病原菌的生理、生化鑒定
用硫代硫酸鹽—檸檬酸鹽—膽鹽—蔗糖瓊脂培養基培養致病菌,觀察其形態、菌落大小。對單個優勢菌落進行革蘭氏染色,于顯微鏡下觀察。挑取新鮮菌落接種于含3%NaCl的營養肉湯中培養18 h,取70 μL含3%NaCl的營養肉湯培養物依次加入微量生化管中,37 ℃下培養8~48 h,觀察結果。
1.4 人工感染試驗
試驗健康蝦為淡化后仔蝦,平均體長約為2.0 cm。人工感染試驗在20 L水族箱中進行,溫度控制在28 ℃,連續充氧。試驗組與對照組分別放入20尾仔蝦,試驗前仔蝦在水族箱內暫養一周,無異常情況后開始試驗。試驗組水體中的細菌密度調至108cfu/mL,對照組不加入菌體。每日正常投喂、換氣、吸污換水,并觀察記錄發病癥狀及死亡情況。
1.5 菌株16S rDNA基因序列分析及系統發育樹的構建
將分離得到的弧菌接種于含3%NaCl的營養肉湯中,28 ℃,200 r/min搖床培養16~18 h,用細菌的通用引物擴增16S rDNA,P1:5′-AGAGTTTGATCCTGGCTCAG-3′,P2:5′-ACGGCTACCTTGTTACGACTT-3′。引物由上海生工生物工程有限公司合成。根據細菌基因組DNA抽提試劑盒的說明書提取分離細菌的DNA。PCR反應體系(50 μL):14 μL 2× Easy Taq Super Mix(含Mg2+、dNTP、DNA聚合酶、buffer), 10 mmol/L正向和反向引物各1 μL,2 μL模板,ddH2O 32 μL。PCR反應條件為:94 ℃預變性5 min,94 ℃ 30 s,55 ℃ 45 s,72 ℃ 90 s,30個循環,72 ℃ 5 min。擴增產物經1%瓊脂糖凝膠電泳檢測后,送至上海生工生物工程有限公司純化并測序。將測序所得序列與GenBank中的核酸序列進行Blast分析,選取同源性較高的序列,采用Clustal X 1.83軟件進行多重序列比對分析,并通過MEGA 6軟件采用鄰接法構建系統發育樹。
1.6 藥敏試驗
在無菌環境下,取100 μL分離純化的新鮮菌液,用無菌涂布棒均勻涂布于含3%NaCl的營養瓊脂平板上,用鑷子無菌操作將每種藥敏片輕輕的貼在上面,28 ℃培養24 h,測量抑菌圈直徑。根據美國臨床實驗室標準化研究所抗菌藥物敏感性試驗執行標準[7],判定該菌對17種抗生素的敏感程度。
2 結果和分析
2.1 病原形態特征
自羅氏沼蝦組織中分離獲得1株細菌QY170324,在硫代硫酸鹽—檸檬酸鹽—膽鹽—蔗糖瓊脂平板上培養18 h后,呈現為圓形光滑、黃色突起的菌落。在含3%NaCl營養肉湯培養時,菌液呈現均勻渾濁。顯微鏡下觀察為革蘭氏陰性菌(圖1)。
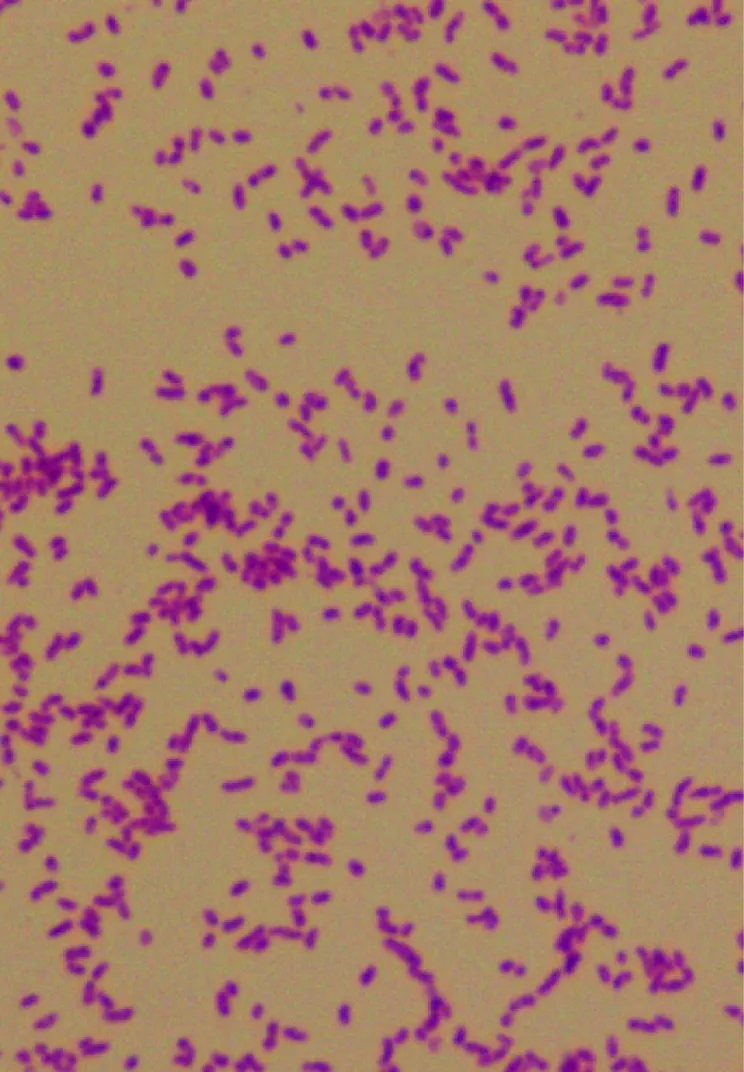
圖1 菌株QY170324革蘭氏染色結果(1000×)
2.2 病原菌的生理、生化鑒定
由分離菌的生理、生化特性的檢測結果可知,該病原菌與溶藻弧菌(V.alginolyticus)極為相似(表1),依據《伯杰氏細菌鑒定手冊》,初步鑒定該病原菌為溶藻弧菌。
2.3 人工感染試驗
人工感染3 d后,羅氏沼蝦開始出現死亡,7 d后全部死亡,病死率達100%。觀察期內取感染死亡仔蝦勻漿處理后進行細菌分離,并進行革蘭氏染色鏡檢、生化鑒定及分子生物學鑒定,結果顯示分離菌特征與原感染菌基本一致,所以初步判斷廣東省湛江市某養殖場羅氏沼蝦發病是由該致病菌所致。
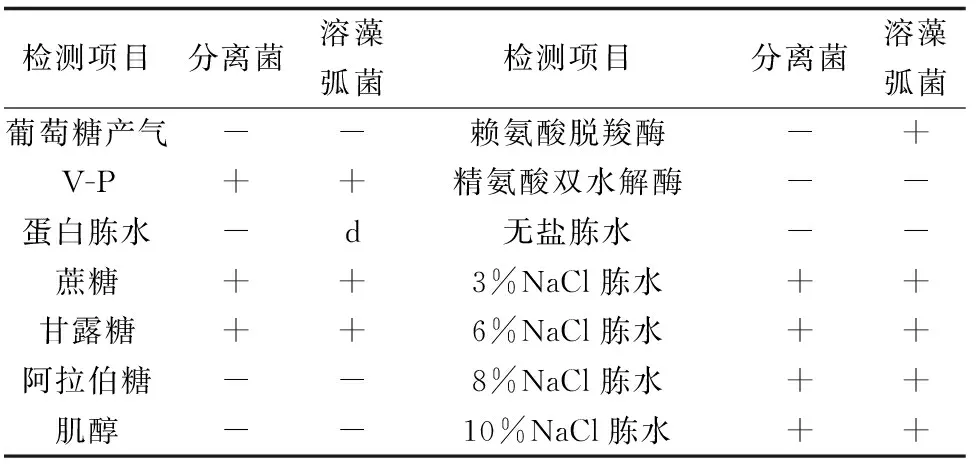
表1 分離菌的生理生化特征
注:“+”表示反應為陽性,“-”為陰性,“d”為結果未定.
2.4 菌株16S rDNA基因序列的鑒定及發育樹的構建
將分離菌所擴增的16S rDNA基因序列提交到GenBank,獲得序列號為MF187358,Blast比對結果顯示,菌株QY170324基因與溶藻弧菌相應基因相似率達99%。所構建的系統發育樹見圖2,菌株QY170324與溶藻弧菌聚為一支。
2.5 藥敏試驗
藥敏試驗結果顯示,溶藻弧菌QY170324對慶大霉素、四環素、氧氟沙星、哌拉西林、頭孢他啶、復方新諾明等抗生素敏感,對頭孢曲松、卡那霉素、新霉素、米諾環素、麥迪霉素、頭孢唑林、多黏菌素B中度敏感,對克林霉素、羧芐西林、頭孢氨芐耐藥(表2)。
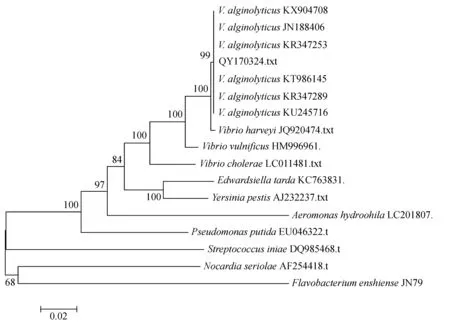
圖2 基于16S rDNA序列所構建的系統發育樹
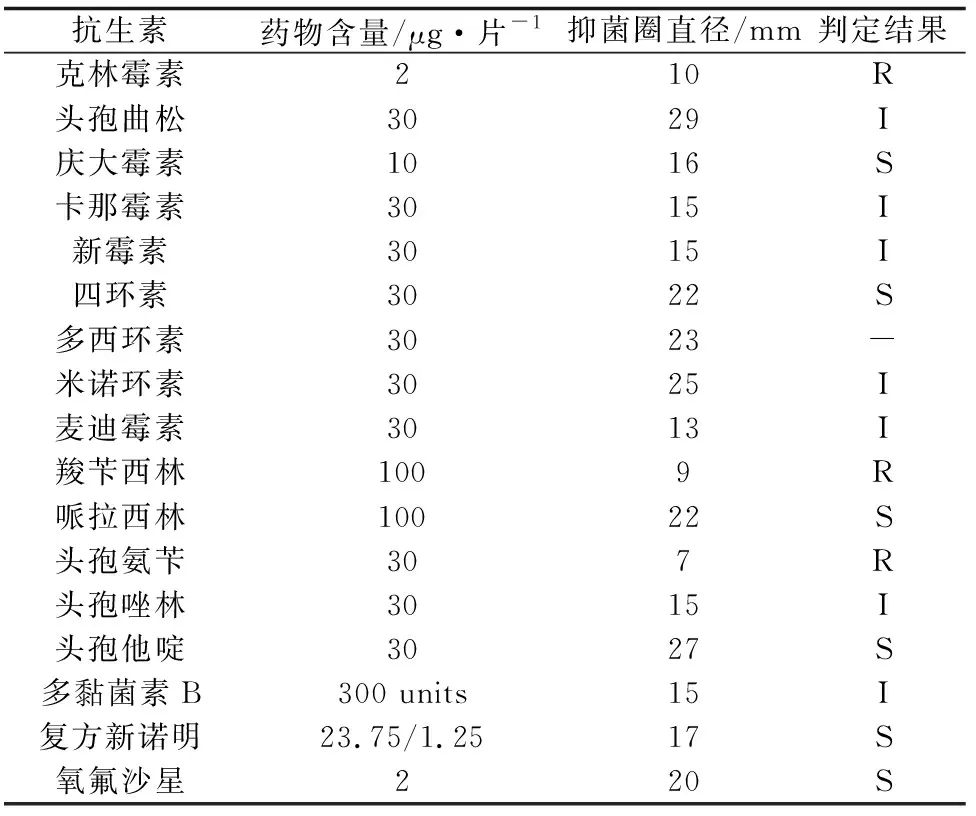
抗生素藥物含量/μg·片-1抑菌圈直徑/mm判定結果克林霉素210R頭孢曲松3029I慶大霉素1016S卡那霉素3015I新霉素3015I四環素3022S多西環素3023-米諾環素3025I麥迪霉素3013I羧芐西林1009R哌拉西林10022S頭孢氨芐307R頭孢唑林3015I頭孢他啶3027S多黏菌素B300 units15I復方新諾明23.75/1.2517S氧氟沙星220S
注:S為高度敏感,I為中度敏感,R為不敏感;-:無對應的參考標準.
3 討 論
3.1 病原菌的分離鑒定及對羅氏沼蝦的致病力
溶藻弧菌屬弧菌科、弧菌屬,專屬性嗜鹽,是一類廣泛分布于世界范圍內的革蘭氏陰性菌,在河口處和海水中最多[8]。該菌不僅對水生動物危害大,亦可導致人類的許多疾病,如引起人食物中毒、腸道炎、傷口感染、中耳炎等[9],是重要的食源性病原菌之一。溶藻弧菌能夠感染許多水產養殖動物,如紫貽貝(Mytilusgalloprovincialis)[10]、魁蚶(Scapharcabroughtonii)[11]、大黃魚(Pseudosciaenacrocea)[12-13]、花鱸(Lateolabraxjaponicus)[14]、凡納濱對蝦(Litopenaeusvannamei)[15]、三疣梭子蟹(Portunustrituberculatus)[16]、曼氏無針烏賊(Sepiellamaindroni)[17]等水產動物發病。近幾年有學者報道,溶藻弧菌也可以感染半滑舌鰨(Cynoglossussemilaevis)[18],本研究首次報道了在羅氏沼蝦養殖過程中溶藻弧菌作為潛在的細菌性病原體也可感染羅氏沼蝦導致死亡的案例,說明溶藻弧菌的感染范圍在擴大,應引起重視。
本試驗在廣東省湛江市某養殖場的患病死亡仔蝦體內分離得到一株病原菌QY170324,運用分子生物學方法測定16S rDNA基因序列。16S rDNA 基因檢測技術目前已成為病原菌檢測和鑒定的強有力工具,應用該技術可以實現對病原菌快速、微量和準確的分類鑒定[19]。因此,本研究運用16S rDNA基因、同時結合生理生化試驗及回歸感染試驗,綜合判定導致羅氏沼蝦死亡的致病菌為溶藻弧菌。本試驗分離到了賴氨酸脫羧酶為陰性的致病性溶藻弧菌,與徐曉麗等[18]從半滑舌鰨體內分離到的賴氨酸脫羧酶為陰性的溶藻弧菌結果相同,與Beleneva等[20]提出溶藻弧菌賴氨酸脫羧酶陽性(+)結果不同,可能是細菌代謝受環境、溫度等因素影響,導致菌株之間生化特性具一定的差異性。所以,在判定溶藻弧菌的種屬時,還應結合生理特征、分子生物學等方面的特性進行綜合判斷。本試驗對分離菌株QY170324的16S rDNA序列進行了分析,序列比對及系統發育樹的構建證實該菌為溶藻弧菌。
本研究中,人工感染后羅氏沼蝦的癥狀與自然發病時的癥狀相似,并自患病的幼蝦體內重新分離到了與溶藻弧菌QY170324形態、生化特征一致的細菌,證實這株細菌即為本次導致羅氏沼蝦發病的病原菌。
3.2 用于防治羅氏沼蝦溶藻弧菌病抗生素的篩選
目前,抗生素治療仍然是水產養殖動物細菌性疾病治療的一種重要手段[21]。由藥敏試驗結果可知,溶藻弧菌QY170324對慶大霉素、四環素、氧氟沙星、哌拉西林、頭孢他啶、復方新諾明6種藥物高度敏感,對頭孢曲松、卡那霉素、新霉素、米諾環素、麥迪霉素、頭孢唑林、多黏菌素B 8種藥物中度敏感,對克林霉素、羧芐西林、頭孢氨芐耐藥,所以,在疾病發生后選擇藥物時應結合藥敏試驗結果,有針對性地篩選適合的藥物來治療由該菌引起的疾病,避免造成誤用、濫用漁藥而導致副作用的發生和一些耐藥菌株的出現,同時還應考慮到養殖動物對藥物的耐受能力,合理安排用藥量或者選用一些窄譜抗生素,更加有目的性地對病原菌進行治療,以至于達到高效、低殘留的目的。近些年來,羅氏沼蝦大多以工廠化養殖為主,在養殖過程中,應降低養殖密度,定期對循環水設備進行維護,定期監測水體中弧菌總數的變化,同時也可在飼料中添加一定量的免疫增強劑來提高羅氏沼蝦的抗病力。
溶藻弧菌對水產養殖業造成巨大的經濟損失,同時對食品衛生安全也造成嚴重的隱患。對溶藻弧菌病原學特征和流行病學的研究,可為養殖場預防及防治溶藻弧菌提供科學依據。
[1] 謝駿,劉波明,俊超,等. 大黃蒽醌提取物對羅氏沼蝦高溫下抗力與熱應激蛋白70基因表達的影響[J]. 水產學報, 2010, 6(34):973-980.
[2] 朱錦裕,卜弘毅,胡沖沖,等, 潑灑糖蜜對池塘養殖羅氏沼蝦生長和水質的影響[J]. 水產科學, 2017,36(2):202-206.
[3] Alambra J R, Alenton R R R, Gulpeo P C R, et al. Immunomodulatory effects of turmeric,Curcumalonga(Magnoliophyta, Zingiberaceae) onMacrobrachium[J]. Aquaculture, Aquarium, Conservation & Legislation, 2012(5):13-17.
[4] 劉海春,董學洪,劉志國,等. 羅氏沼蝦健康養殖水質調控技術[J]. 江蘇農業科學, 2011, 39(10):338-341.
[5] Kennedy B. Bacterial flora associated with the giant freshwater prawnMacrobrachiumrosenbergii,in the hatchery system[J]. Aquaculture, 2006, 261(4):1156-1167.
[6] Lee S W, Najiah M. Multiple antibiotic resistance and heavy metal resistance profile of bacteria isolated from giant freshwater prawn (Macrobrachiumrosenbergii) hatchery[J]. Science Direct,2009,8(6):740-745.
[7] Wikler M A.M100-S18, Performance standards for antimicrobial susceptibility testing: Eighteenth Informationgal Supplement[S]. USA: Clinical and Laboratory Standards Institute, 2008.
[8] 滕勇勇,王琪,吳雷,等. 致病性弧菌的生物學特性和致病因子研究進展[J]. 熱帶醫學雜志, 2014, 14(10):1396-1399.
[9] 陳強,鄢慶枇,馬甡,等. 溶藻弧菌致病性研究進展[J]. 海洋科學, 2006, 30(8):83-89.
[10] 林永添. 紫貽貝(MytilusedulisLinnaeus)大量死亡原因的初步研究[J]. 現代漁業信息, 2006, 21(7):23-26.
[11] 韓風杰,徐文娜,劉敏,等. 魁蚶溶藻弧菌分離鑒定與藥敏試驗[J]. 山東畜牧獸醫, 2016,37(5):13-14.
[12] 桑本紅,徐曉津,毛勇,等. 大黃魚溶藻弧菌病組織中巨噬細胞移動抑制因子的檢測及意義[J]. 海洋學報, 2014, 36(8):111-116.
[13] 賴迎迢,陶家發,孫承文,等. 魚源溶藻弧菌生物學特性和病理組織學觀察[J]. 微生物學報, 2014, 54(11):1378-1384.
[14] 陳寅兒,金珊,王國良,等. 鱸魚溶藻弧菌病的血液生理生化指標研究[J]. 臺灣海峽, 2005, 24(1):104-108.
[15] 羅詞興,黃旭雄,李桑,等. 溶藻弧菌感染后凡納濱對蝦鰓組織免疫相關基因的表達[J]. 中國水產科學, 2014, 21(1):189-196.
[16] 劉淇,李海燕,王群,等. 梭子蟹牙膏病病原菌—溶藻弧菌的鑒定及其系統發育分析[J]. 海洋水產研究, 2007, 28(4):9-13.
[17] 李正義,賈俊濤,曹際娟,等. 一株分離自凍墨魚的溶藻弧菌的鑒定及其低溫下細胞脂肪酸的變化[J]. 水產科學, 2014, 33(5):317-321.
[18] 徐曉麗,葉紅梅,侯純強,等.半滑舌鰨弧菌病病原的分離鑒定[J]. 中國獸醫雜志, 2017(8):86-90.
[19] 程俊茗,萬明月,周晉揚,等.鯽源遲緩愛德華氏菌的分離鑒定及其毒力基因的檢測[J]. 微生物學通報, 2017, 44(10):2380-2390.
[20] Beleneva I A. Physiological and biochemical characteristics of the halophilic bacteriaVibrioparahaemolyticusandV.alginolyticusisolated from marine invertebrates of Peter the Great Bay, Sea of Japan[J]. Microbiology, 2004, 30(2):96-100.
[21] 刁菁,楊秀生,葉海斌,等. 一種水產遲鈍愛德華氏菌快速藥敏檢測方法的研究[J]. 水產科學, 2014,33(1):22-28.
