高鈣和缺鈣環境對花鰻鱺血液生理指標及其NCXs和PMCAs基因表達的影響
2018-08-07 06:30:44曹全全胡亞東張宏葉李欣茹尹紹武
海洋漁業 2018年4期
關鍵詞:環境
曹全全,胡亞東,王 濤,張宏葉,顧 杰,王 丹,李欣茹,尹紹武
(1.南京師范大學生命科學學院,海洋科學與工程學院,江蘇南京 210023;2.江蘇大學生命科學學院,江蘇鎮江 212000)
花鰻鱺(Anguilla marmorata)屬鰻鱺目(Anguilliformes),鰻鱺科(Anguillidae),鰻鱺屬,廣泛分布于太平洋周圍的熱帶和亞熱帶區域,為降海洄游性魚類。因其肉質鮮美細嫩,市場需求大,導致捕撈過度,被《中國瀕危動物紅皮書》列為瀕危物種,并于上20世紀90年代被我國列為二級保護動物[1]。在自然條件下,鰻鱺因具特殊的生活史洄游于淡水和海水之間且頻繁地面對滲透壓的挑戰。鹽度和鈣離子濃度是影響魚類滲透壓最重要的兩個外界指標[2]。鈣離子是海水中的重要組成部分之一,且其濃度在海水表層與深水中變動較大,而在魚體內卻維持在一個較窄的范圍內[3]。因此維持鈣穩態對于鰻鱺屬魚類來說具有重要的意義。
鈉鈣交換體NCX是一種膜內在蛋白[4],其中NCX1、NCX2和NCX3這3種亞型已在哺乳動物如 鼠 (Rattus norvegicus)[5]、豚 鼠 (Cavia porcellus)[6]和狗(Canis lupus familiaris)[7]中克隆得到。NCX1在機體組織中廣泛分布,而NCX2和NCX3主要分布于腦和骨骼肌中[8]。在魚類研究中,NCX1在虹鱒(Oncorhynchus mykiss)[9]、莫桑比克羅非魚(Oreochromis mossambicus)[10]和斑馬魚(Barchydanio rerio)[11]等魚類中已被克隆。NCX的主要功能是將鈣離子從細胞內排出以防止胞內儲存的鈣離子超負荷輸出,從而維持胞漿中鈣離子濃度的恒定[12]。質膜 Ca2+泵(PMCA)是一種Ca2+-Mg2+-ATP酶,通過水解ATP來維持Ca2+的輸出從而達到維持細胞低濃度鈣離子的目的。PMCA包含10個跨膜結構域、5個推測的胞外域和3個主要細胞結構域蛋白。這些結構域均屬于功能位點,是鈣離子綁定位點,被鈣調蛋白激活;也是ATP綁定位點,對于酰基磷酸酐的形成和水解有著重要的作用[12-13]。人類PMCA分為4個亞型,分別為 PMCA1、2、3和4。PMCA1和 PMCA4在哺乳動物中具有廣泛分布,被譽為維持細胞內鈣穩態的看家亞型,而PMCA2和PMCA3的表達模式具有組織特異性[14]。
鈣離子轉運體NCX、PMCA等通過上皮細胞Ca2+的運輸和轉運,來控制鈣離子進出體內的凈含量,進而影響多個生理學進程。此外,鈣離子的循環量受到機體嚴格的限制,魚類需具備感知外界鈣離子的能力進而調節鈣離子的穩態[15]。在哺乳動物的腎臟中,鈣離子通過鈣通道透過質膜,然后依托鈣結合蛋白擴散穿出質膜[16]。FLIK等[17]在魚鰓中提出一個模型,與哺乳動物腎臟鈣離子的重吸收相似,Ca2+依附胞液鈣結合蛋白從胞液排出到血液中,需細胞底外側PMCA和NCX以及頂膜鈣離子泵(ECaCs)的參與,但關于這些鈣轉運體對整個鈣離子吸收進程的相對重要性并沒有闡述。
為了探究花鰻鱺NCXs和PMCAs基因在極性鈣環境下(高鈣10 mM和缺鈣0 mM)的表達規律,本研究測量了花鰻鱺成鰻從對照組轉移到高鈣組和缺鈣組時的血清鈣濃度、血清滲透壓和5項血液指標(白細胞數目、紅細胞數目、葡萄糖濃度、血紅蛋白濃度和血小板數目),并采用qRTPCR技術對NCXs和PMCAs基因在花鰻鱺13種不同組織(腦、鰓、頭腎、后腎、前腸、中腸、后腸、肝、心、肌肉、鰾、脾和皮膚)中的表達進行了分析,最后研究了花鰻鱺應對極性鈣濃度下(高鈣10 mM和缺鈣0 mM)NCXs和PMCAs基因在鰓組織中的mRNA表達水平變化規律。以期為進一步解析花鰻鱺鈣穩態的分子機制提供基礎資料。
1 材料與方法
1.1 實驗材料
花鰻鱺體質量(550±10)g,取自中國海南文昌金山花鰻鱺科技有限公司(公司具有中華人民共和國水生動物捕撈資格),暫養于玻璃循環缸中15 d,每日定期投喂飼料,攝食30 min后虹吸排污,每日換1/3水體。實驗用水為曝氣24 h的自來水,養殖水溫保持在25~27℃。確定魚體健康無病后開始實驗。
1.2 總RNA的提取及cDNA第一鏈的合成
用北京百泰克公司的高純總RNA快速抽提試劑盒(離心柱型)提取花鰻鱺組織總RNA,用RNA濃度測定儀測定每個樣品的RNA濃度及純度,確保 OD 260/280在1.9~2.1范圍內。用1%瓊脂糖凝膠電泳檢測其完整性。所有樣品凍存于-80℃冰箱中備用。以總RNA為模板,根據 HiScriptTM1ST strand cDNA Synthesis kit(南京諾唯贊生物科技有限公司)的說明進行反轉錄,獲得cDNA第一條鏈。
1.3 花鰻鱺NCXs和PMCAs基因的組織表達分析
隨機取缸內3尾花鰻鱺成鰻,MS-222(Sigma)(150 mg·L-1)麻醉后解剖取 13種組織(腦、鰓、頭腎、后腎、前腸、中腸、后腸、肝、心、肌肉、鰾、脾和皮膚),提取總RNA后,反轉錄獲得cDNA第一鏈。根據花鰻鱺轉錄組及其近緣物種的NCXs和PMCAs序列,分別用Primer 5設計特異性引物 NCX1-F、NCX1-R、NCX2-F、NCX2-R、NCX3-F、 NCX3-R、 PMCA2-F、 PMCA2-R、PMCA4-F、PMCA4-R以及作為內參基因的花鰻鱺actin基因特異性上下游引物actin7、actin8(表1)。以獲得的13種組織的cDNA為模板,進行實時熒光定量 PCR分別檢測花鰻鱺 NCXs和PMCAs在13種組織中mRNA的表達量。PCR反應體系為20μL:Faststart Universal SYBR Green Master 10μL,正反引物(2μmol·μL-1)各3μL,cDNA模板(5 ng·μL-1)4μL。反應程序為95℃預變性5 min;94℃ 30 s,55℃ 30 s,72℃ 1 min,35個循環;72℃ 延伸5 min,4℃保存。

表1 花鰻鱺NCXs和PMCAs基因熒光定量的引物Tab.1 Primers used for Anguilla marmorata NCXs and PMCAs in qRT-PCR
1.4 人工鈣離子水的配置
參照CHEN等[18]所描述的鈣離子溶液的配置方法,用 CaSO4、MgSO4、NaCl、K2HPO4、KH2PO4和去離子水(Milli-RO60,Millipore,Bedford,MA)配置不同鈣梯度的水溶液。實驗鈣濃度分為:1)組 A:對照組(pH=7,2 mM Ca2+);2)組B:缺鈣組(pH=7,0 mM Ca2+);3)組 C:高鈣組(pH=7,10 mM Ca2+),其它3種溶液的鈣離子濃度相同([Na+],0.5 mM;[Mg2+],0.16 mM;[K+],0.3 mM)。實驗用魚先放置在組A中馴化1周,1周后把魚放置到組B和組C中進行相關的時序表達實驗。為保持水中鈣濃度穩定,每6 h換1次相應濃度的鈣離子水,換水量為50%,實驗過程中持續充氧。
1.5 Ca2+脅迫對花鰻鱺NCXs和PMCAs基因表達的影響
為進一步探究花鰻鱺在不同鈣脅迫下、不同時段的鰓組織中NCXs和PMCAs基因表達水平的變化規律,實驗各組分別于第1、4和7天,每個缸內隨機取3尾魚,用MS-222(Sigma)麻醉,取其鰓組織,液氮速凍10 min后于-80℃保存。隨即用注射器經主動脈抽出血液,靜置后取血清,用原子吸收光譜法(SpectrAA-220FS,Varian,US)測量鈣離子濃度。血清滲透壓用蒸汽壓滲透壓計(Wescor,5500XR,Logan,UT)測量其滲透壓值。用自動血液分析儀(BC-2800vet,Manrui,China)測量紅細胞數目、白細胞數目和血小板數目。葡萄糖濃度和血紅蛋白濃度用標準試劑盒測定(Jiancheng Bioengineering,Nanjing,China)。提取鰓組織總RNA并檢測NCXs和PMCAs基因的相對表達量。
1.6 數據處理
所有數據采用均值±標準差(Means±SD)表示。采用 2-ΔΔCt法進行數據處理[21]。利用統計學軟件SPSS 23對時間和鈣離子濃度進行雙因子方差分析(two-way ANOVA)(表2),組織表達用單因子方差分析(one-way ANOVA),通過t檢驗計算P值,若P<0.05,則差異顯著。用Origin 8.6繪制柱狀圖。

表2 時間和鈣離子濃度的雙因素分析Tab.2 Two-way ANOVA of time and Ca2+concentration
2 結果與分析
2.1 不同鈣離子脅迫下花鰻鱺血清鈣濃度和血清滲透壓的變化規律
如圖1所示,血鈣濃度(圖1-A)和血清滲透壓(圖1-B)與周圍鈣濃度成正相關。在高鈣離子環境中,血鈣濃度和血清滲透壓隨著時間的延長成下調趨勢;在缺鈣環境中,血鈣濃度和血清滲透壓隨著時間的增加成上調趨勢。3種鈣濃度組血鈣濃度和血清滲透壓在第7天差異均不顯著(P>0.05)。
2.2 不同鈣離子濃度下對花鰻鱺血液學指標的影響
如圖2所示,白細胞數目(圖2-A)、紅細胞數目(圖2-B)、血小板數目(圖2-C)、血紅蛋白濃度(圖2-D)和葡萄糖濃度(圖2-E)和周圍鈣濃度成正相關。在高鈣與缺鈣環境中,白細胞數目、紅細胞數目、葡萄糖濃度、血紅蛋白濃度和血小板數目隨著時間的延長成下調趨勢,且較對照組在第1、4天具有顯著性差異(P<0.05)。而在第7天白細胞數目、紅細胞數目、葡萄糖濃度、血紅蛋白濃度和血小板數目各組的差異并不顯著(P>0.05)。
2.3 NCXs和PMCAs基因在花鰻鱺不同組織表達分布
qRT-PCR結果顯示(圖3),NCXs和 PMCAs基因在花鰻鱺腦、鰓、頭腎、后腎、前腸、中腸、后腸、肝、心、肌肉、鰾、脾和皮膚這13種組織中均有表達,具有廣泛表達的特點。其中,花鰻鱺NCX1(圖3-A)和 PMCA4(圖3-E)基因在頭腎中表達量最高,NCX2(圖3-B)和 NCX3(圖3-C)在皮膚中表達量最高,PMCA2(圖3-D)在后腎中表達量最高,而在前腸中5個基因表達量則最低。
2.4 不同鈣處理、不同時段下花鰻鱺NCXs和PMCAs基因的表達模式
在急性高鈣脅迫下,花鰻鱺NCX1(圖4-A)和PMCA4(圖5-B)基因在鰓組織的mRNA表達在第1天均顯著上調;在缺鈣環境中,其mRNA表達量顯著下調。對于NCX1基因,隨著適應時間的延長,在第4天和第7天高鈣環境下的mRNA表達量顯著上升,而在缺鈣環境中mRNA表達量顯著降低,在第7天各處理組差異顯著(P<0.05);對于 PMCA4,隨著適應時間的延長,在第4天和第7天高鈣環境下PMCA4基因的mRNA表達量顯著下調,而在缺鈣環境中mRNA表達上調,在第7天各處理組無顯著差異(P>0.05)。但對于 NCX2(圖 4-B)、NCX3(圖 4-C)和PMCA2(圖5-A),其表達量隨時間無明顯的變化。
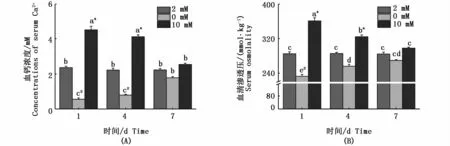
圖1 不同鈣濃度下花鰻鱺脅迫1、4和7 d的血鈣濃度和血清滲透壓Fig.1 Serum osmolarity and Ca2+concentration in Anguilla marmorata adapted to different Ca2+ concentrations for 1 day,4 or 7 days注:不同時間點和不同鈣離子濃度的差異顯著性用不同字母表示。同一時間點不同處理組之間的差異性用不同符號*(高鈣組與對照組具有顯著性差異)和#(缺鈣組與對照組具有顯著性差異)表示(P<0.05,two-way ANOVA)Note:Significant differences between Ca2+concentrations and different time points are identified by different letters;and different symbols*and#indicate significant differences between groups at the same time point(P<0.05,two-way ANOVA)
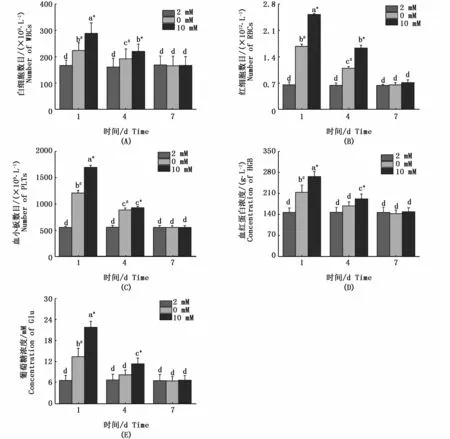
圖2 不同鈣濃度下花鰻鱺脅迫1、4和7 d的各項血液指標Fig.2 Hematology index in Anguilla marmorata adapted to different Ca2+ concentrations for 1 day,4 or 7 days注:不同時間點和不同鈣離子濃度的差異顯著性用不同字母表示。同一時間點不同處理組之間的差異性用不同符號*(高鈣組與對照組具有顯著性差異)和#(缺鈣組與對照組具有顯著性差異)表示 (P<0.05,two-way ANOVA)Note:Significant differences between Ca2+concentrations and different time points are identified by different letters;and different symbols*and#indicate significant differences between groups at the same time point(P<0.05,two-way ANOVA)
3 討論
Ca2+是自然海水的重要組成成分,對魚類的神經元興奮性、肌肉收縮、細胞滲透性、細胞分化、激素釋放和骨組織的礦物化等生理功能均具有重要影響[19]。海水中鈣離子濃度約為10 mM,淡水中鈣離子濃度約為2 mM,洄游性魚類在洄游過程中需要面對不斷變化的鈣離子濃度[17]。因此,研究洄游性魚類體內控制鈣離子的進出機制及鈣離子的穩態具有非常重要的意義。CRUZ等[20]提出在魚類鰓細胞表面的跨膜轉運是由上皮鈣通道調節,而這個鈣通道則由質膜鈣ATP酶PMCA和鈣轉運體 NCX組成。HOENDEROP等[15]研究表明鈣離子的流出需一個較大的電化學梯度,位于吸收細胞質膜上的兩個鈣離子轉運體需NCX和PMCA負責將鈣離子排出。考慮到NCX和PMCA是主要的運輸Ca2+的跨膜蛋白,因此我們選擇NCX和PMCA為靶蛋白,通過極性鈣濃度脅迫來測定NCX基因3個亞型和PMCA基因2個亞型的mRNA時序表達,其次,測定了這5種基因亞型的組織表達分布,以及極性鈣濃度環境下花鰻鱺血清中的鈣濃度、滲透壓和血液生理指標,為魚類鈣穩態的研究提供基礎資料。
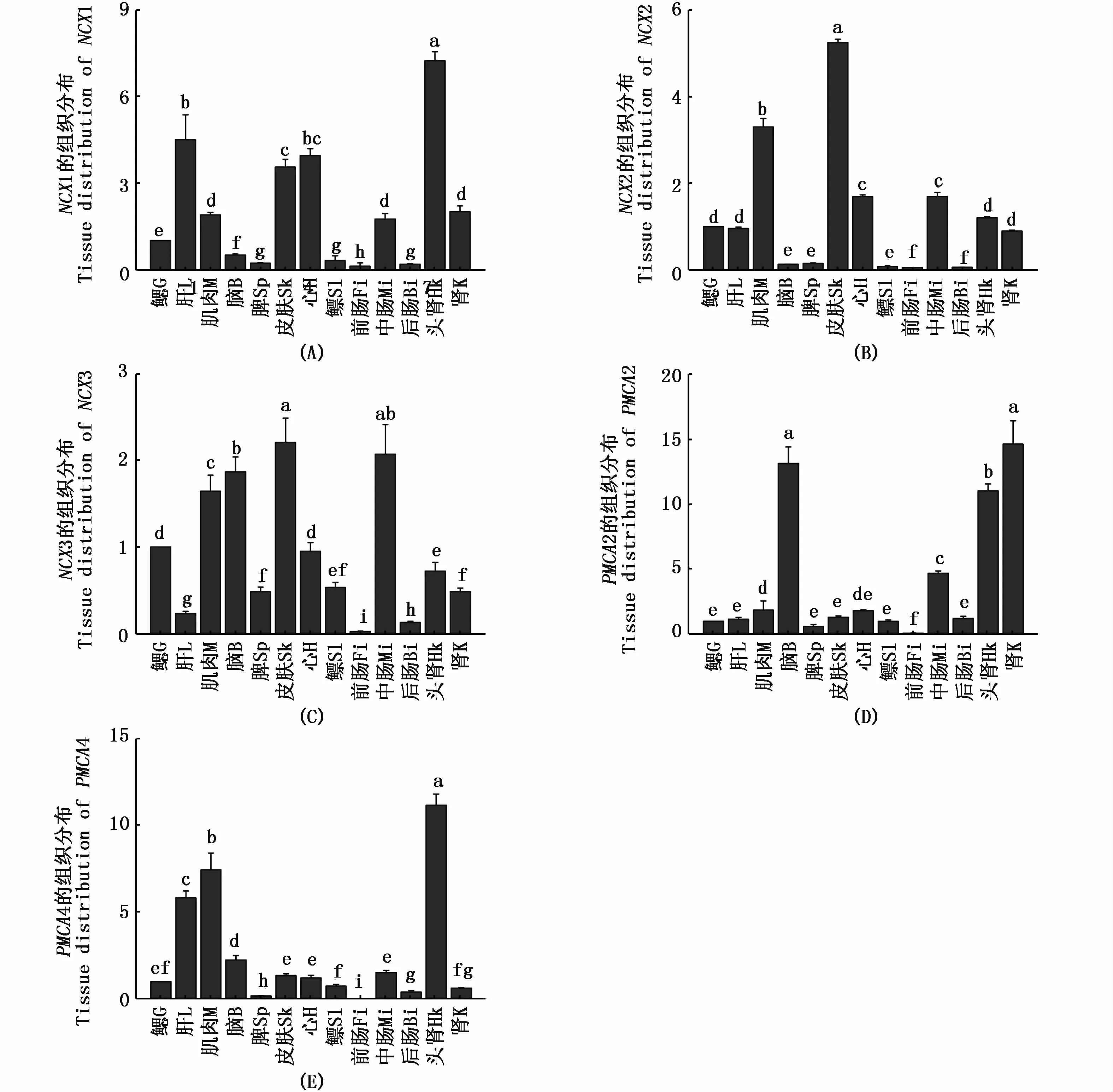
圖3 花鰻鱺 NCX1(A)、NCX2(B)、NCX3(C)、PMCA2(D)和 PMCA4(E)在不同組織中的表達規律Fig.3 Tissue distribution analysis of NCX1(A),NCX2(B),NCX3(C),PMCA2(D),and PMCA4(E)at the mRNA level注:不同組織相對表達的差異顯著性用不同字母表示 (P<0.05,一維方差分析)。G:gill;L:liver;M:muscle;B:brain;Sp:spleen;Sk:skin;H:heart;Sl:swim bladder;Fi:front intestine;Mi:middle intestine;Bi:back intestine;Hk:head kidney;K:kidneyNote:Significant differences among different tissues are indicated by different letters(P <0.05,one-way ANOVA).G:gill;L:liver;M:muscle;B:brain;Sp:spleen;Sk:skin;H:heart;Sl:swim bladder;Fi:front intestine;Mi:middle intestine;Bi:back intestine;Hk:head kidney;K:kidney
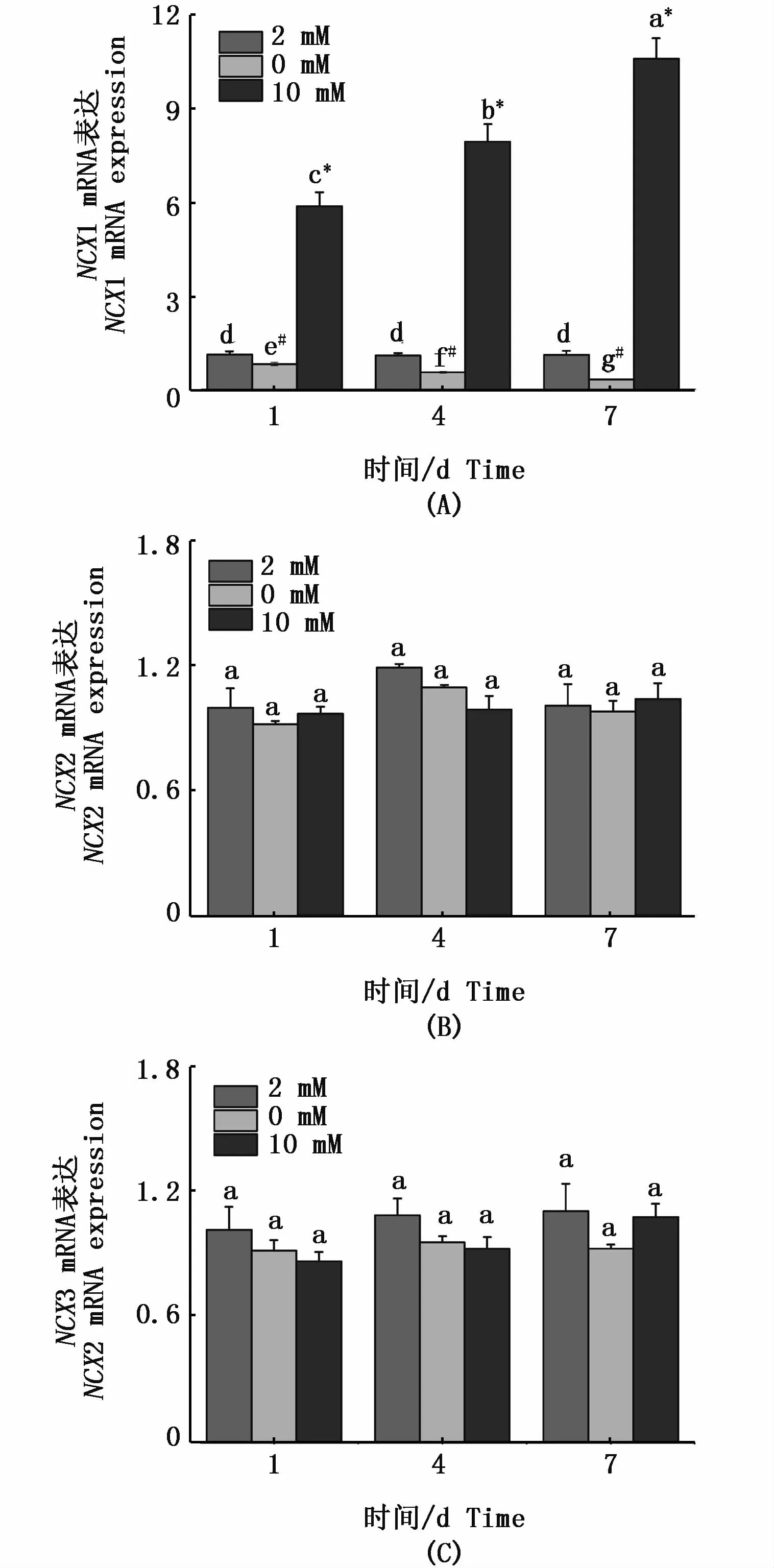
圖4 花鰻鱺 NCX1(A)、NCX2(B)和 NCX3(C)在鰓中不同時間點應對不同鈣濃度的表達規律Fig.4 Expressions of NCX1(A),NCX2(B)and NCX3(C)in gill of Anguilla marmorata in response to different Ca2+concentrations at different time points注:不同時間點和不同鈣離子濃度的差異顯著性用不同字母表示。同一時間點不同處理組之間的差異性用不同符號*(高鈣組與對照組具有顯著性差異)和#(缺鈣組與對照組具有顯著性差異)表示 (P<0.05,two-way ANOVA)Note:Significant differences between Ca2+ concentrations and different time points are identified by different letters;and different symbols*and#indicate significant differences between groups at the same time point(P<0.05,two-way ANOVA)
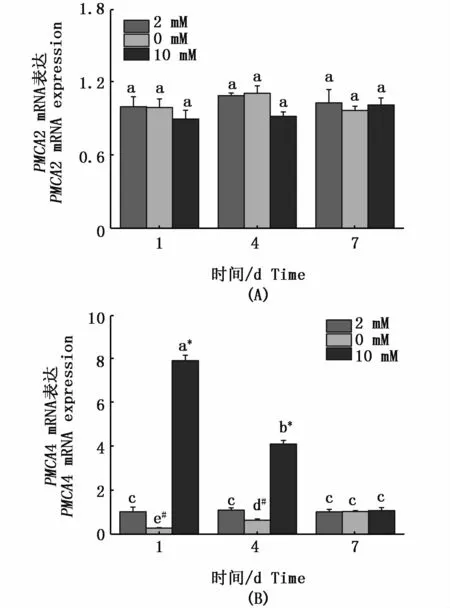
圖5 花鰻鱺PMCA2(A)和PMCA4(B)在鰓中不同時間點應對不同鈣濃度的表達規律Fig.5 Expressions of PMCA2(A)and PMCA4(B)in gill of Anguilla marmorata in response to different Ca2+concentrations at different time points注:不同時間點和不同鈣離子濃度的差異顯著性用不同字母表示。同一時間點不同處理組之間的差異性用不同符號*(高鈣組與對照組具有顯著性差異)和#(缺鈣組與對照組具有顯著性差異)表示(P<0.05,two-way ANOVA)Note:Significant differences between Ca2+ concentrations and different time points are identified by different letters;and different symbols*and#indicate significant differences between groups at the same time point(P<0.05,two-way ANOVA)
目前國內對于魚類滲透壓的研究主要集中在鹽度脅迫,王丹等[22]研究了如花鰻鱺AQP3基因在不同鹽度下表達規律。而鈣脅迫對魚類血清滲透壓的影響報道不多。褐牙鲆(Paralichthys olivaceus)低鈣條件下血清滲透壓在6 h時顯著上升,而12 h后下降至正常水平并維持穩定[23]。在本研究中也出現了穩態現象,缺鈣環境下血清滲透壓和鈣離子濃度起初較低,但隨著時間的增加顯著地上升,在第7天3種環境下的鈣離子濃度達到穩態。同樣在高鈣環境脅迫1 d后,血清滲透壓和血鈣濃度逐漸下降到穩定狀態,此結果表明,隨著周圍環境鈣濃度的增加,花鰻鱺主動或被動地減少了鈣離子的吸收。CARRIER等[24]認為,水中Ca2+濃度的變化可增強鰓上皮細胞對水和離子的通透性。血細胞對魚體自身生理狀態的變化及外界環境因子刺激非常敏感,其血液指標能反映魚類的正常生理狀態[25]。筆者測定了各項血液指標(白細胞數目、紅細胞數目、葡萄糖濃度、血紅蛋白濃度和血小板數目),處理組相較對照組各項指標隨時間呈先升高后下降的趨勢,在第7天各項指標呈穩定狀態。以上結果均表明花鰻鱺體內存在一種鈣離子調節機制以維持機體內的鈣離子穩態。
組織表達結果顯示,花鰻鱺NCX和PMCA基因的各個亞型在腎臟中均呈現高表達,在腎臟中通過過濾吸收鈣離子致使腎組織表達量相對較高,這與腎臟作為動物吸收鈣離子的主要器官相吻合[16]。另外,動物主要通過食物來獲得鈣,其全身的鈣守恒主要通過腸的吸收和腎臟的重吸收來維持進行[26]。本實驗中一些鈣通道蛋白皮膚組織的表達量較高,如NCX1和NCX2,筆者推測魚類主要通過皮膚和鰓來增強鈣離子的吸收,由于皮膚和鰓與外界的水環境直接接觸且皮膚和鰓是離子交換的主要場所[26]。LIN等[27]研究顯示魚類從淡水中獲得鈣離子主要通過鰓和腸,而鰓是鈣離子吸收的主要場所。LAFONT等[16]通過對斑馬魚上皮鈣離子通道NCX1和PMCA2特異性表達的研究,結果顯示兩個基因在富含線粒體的鰓細胞中表達量最高。本研究中鰓組織表達量并不高,筆者推測是由于鈣離子的吸收機制發生在鰓中富含線粒體的細胞,其中鈉鈣交換體NCX與質膜鈣離子泵PMCA參與鈣離子的吸收,它們從質膜中排出鈣離子到血液中,故鈣離子在鰓中的濃度降低,所以鰓中的表達量并不高。綜上,本文以鰓為靶器官作為研究,探索外界鈣離子濃度變化時花鰻鱺NCX和PMCA哪些亞型的表達發生了變化,為魚類洄游中鈣穩態的研究提供了一定的基礎資料。
鰓組織中鈣離子轉運體的mRNA表達量反應了鰓中的鈣含量。在本研究中,隨著缺鈣環境時間的延長,NCX1亞型基因mRNA表達量逐漸減弱,高鈣環境下NCX1亞型基因mRNA表達量顯著增長,而其它NCX亞型在外界鈣濃度環境改變下表達量均未發生改變。有研究報道,控制鈣離 子 的 排 出 主 要 靠 NCX1 亞 型[28],LANGENBACHER等[11]認為在魚腸上皮細胞NCX1是細胞膜排出鈣離子的主要蛋白,高鈣環境下可以通過質膜鈣離子泵NCX1將Ca2+排出。由此,筆者推測:為了維持花鰻鱺機體內鈣離子穩態,在高鈣環境下,鰓組織中的NCX1表達量持續升高,促使鰓細胞將過多的鈣離子排出胞外;而在低鈣環境下,NCX1表達量持續降低,以減少鰓細胞中鈣離子的流失。而PMCA4亞型基因在缺鈣環境下表達量逐漸增強,高鈣環境下表達量逐漸降低,在第7天3種鈣濃度下出現鈣穩態狀態。推測低鈣環境可以誘發PMCA鈣運輸能力的增強,這在虹鱒、莫桑比克羅非魚、斑馬魚、金魚 (Carassius auratus)和香魚 (Plecoglossus altivelis)中都有報道[26-30]。HOENDEROP等[15]研究表明,PMCA4在鈣離子的排出機制中具有重要作用,本研究的時序表達驗證了PMCA4在低鈣環境下控制鈣離子的排出,高鈣環境下加快鈣離子的排出,證實了PMCA4具有鈣穩態調控器的作用。而在斑馬魚中,LARONT等[16]研究發現上皮鈣離子通道PMCA2在鰓中MR細胞選擇性表達,并參與跨膜鈣離子吸收,本研究的結果并沒有證實PMCA2在鈣離子的調控作用,筆者推測不同物種間PMCA不同亞型扮演的功能有所不同。在小龍蝦(Procambarus clarkia)鈣離子的輸送機制研究中,WHEATLY等[31]闡述了 NCX家族基因在鈣轉運過程扮演“馱馬”的功能,而PMCA在鈣離子轉運過程中并不扮演任何作用。
本研究顯示,在極性鈣濃度環境下,花鰻鱺鰓組織的NCX1和PMCA4在脅迫不同時段中呈現不同的表達變化模式,并且在各時段中,NCX1和PMCA4基因的表達水平均呈顯著的差異。進一步說明,NCX1和PMCA4在花鰻鱺鈣穩態調節過程中扮演了重要角色。
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學院學報(2017年2期)2017-04-20 08:44:31
中國環境監察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38
