內(nèi)源性γ-干擾素對斑馬魚血管發(fā)育的影響
2018-09-05 03:28:46蔣偉青王永剛
安徽醫(yī)科大學(xué)學(xué)報(bào) 2018年9期
蔣偉青,袁 欣,劉 晨,任 瀟,王永剛
一個成熟的血管網(wǎng)絡(luò)結(jié)構(gòu)的形成需要經(jīng)過復(fù)雜的過程,包括血管發(fā)生、血管生成和其后伴隨的血管形成、血管穩(wěn)定、血管分支、血管修剪和血管分化[1]。已經(jīng)有許多研究報(bào)道了免疫系統(tǒng)在血管生成中的重要作用。而關(guān)于γ-干擾素(interferon-γ,IFN-γ)對血管生成的作用,國內(nèi)外報(bào)道有著不同的結(jié)果。該研究擬以斑馬魚為模式生物,在體研究內(nèi)源性IFN-γ對血管發(fā)育的影響。
1 材料與方法
1.1實(shí)驗(yàn)動物該實(shí)驗(yàn)所用的斑馬魚品系為Tg(Flk1:eGFP),用于標(biāo)記斑馬魚內(nèi)皮細(xì)胞。每個實(shí)驗(yàn)組使用6條斑馬魚幼魚。斑馬魚胚胎養(yǎng)殖于10% Hanks溶液,放置于14 h/10 h明暗交替的28 ℃恒溫培養(yǎng)箱。
1.2胚胎顯微注射顯微注射是指在顯微鏡下,利用玻璃微電極將外源物質(zhì)注射入胚胎細(xì)胞的技術(shù)。玻璃微電極由外徑為1.0 mm、內(nèi)徑為0.58 mm的玻璃毛細(xì)管經(jīng)微電極拉制儀拉制而成。在注射前,將房間溫度調(diào)至20 ℃,以延緩受精卵分裂。在電極內(nèi)灌入要注射的反義嗎啉環(huán)寡聚核苷酸(morpholino oligomer,MO)后,將電極安裝在電極夾持器上并45°傾斜固定于顯微操作器上,鏡下將電極尖端修剪至適合口徑,使每次注射約1 nl。將待注射的物質(zhì)注射入一細(xì)胞期受精卵的動物極中。MO購自美國Gene Tools公司。該實(shí)驗(yàn)所用MO序列如下,對照MO:5’-CCTCTTACCTCAGTTACAATTTATA-3’;crfb17 MO:5’-CGATGAGACACAGATCAATATCCGCAT-3’。利用電極將MO(2 ng)注入1-細(xì)胞期胚胎動物極,另注入等濃度的對照MO作為對照。
1.3斑馬魚活體共聚焦成像用于共聚焦成像的斑馬魚于胚胎受精后天數(shù)(day post fertilization,dpf)為1 dpf時去卵膜,并在Hanks溶液中加入0.003%的 N-苯基硫脲(1-phenyl-2-thiourea)防止其色素形成。成像前,將斑馬魚包埋于濃度為0.013 mg/ml的低熔點(diǎn)膠中。斑馬魚活體共聚焦成像使用Olympus Fluoview 1000激光共聚焦顯微鏡。成像中使用XLumplfl 20×(W/IR NA:0.95) 物鏡。光路、光電倍增管接收光譜范圍、pinhole大小及分光鏡等參數(shù),采用Fluoview 1000軟件染料列表中給出的最優(yōu)參數(shù)。Z軸成像間距取3 μm。
1.4斑馬魚中腦血管三維結(jié)構(gòu)定量分析利用共聚焦顯微鏡對Tg(Flk1:eGFP)的斑馬魚胚胎進(jìn)行XYZ軸成像后,利用該實(shí)驗(yàn)室編寫的腦部血管三維結(jié)構(gòu)定量分析軟件對其中腦網(wǎng)絡(luò)血管特征進(jìn)行分析,包括中腦血管密度、中腦血管節(jié)段數(shù)和中腦血管密度。具體步驟如下:① 原始圖像處理:利用Image J軟件對原始圖片進(jìn)行區(qū)域選擇,范圍區(qū)域需包含所有中腦血管,范圍大小需與模板腦圖像的大小一致,選定區(qū)域后將圖像剪裁,隨后將原始圖像處理為從Z軸由深至淺排列的8位tiff圖;② 圖片等方向性插值:利用實(shí)驗(yàn)室自主編寫的軟件,在菜單欄property setting輸入原始圖片的XYZ軸分辨率,通過 equal insert將處理過得8位tiff圖進(jìn)行等方向性插值處理,并保存;③ 提取中腦血管:通過3D V/B match功能,將血管圖與模板腦圖片進(jìn)行Z軸方向重合,根據(jù)模板腦大小去除中腦以外的血管;④ 圖像分割和去燥:通過segment batch功能對灰度圖片進(jìn)行批量血管分割,分割閾值根據(jù)圖像的灰度值進(jìn)行人工設(shè)定,通過remove noise功能去除部分噪聲點(diǎn);⑤ 圖像三維填充:通過fill 3D功能對分割后血管中的空隙進(jìn)行填充;⑥ 圖像三維重建和中腦血管精細(xì)提取:通過3D viewer binary images的points功能對填充處理過的圖像進(jìn)行三維重建顯示;在三維顯示界面下,將未去除干凈的中腦以外的血管去除,并以原始圖像為參照,將因信號過弱而導(dǎo)致斷裂的血管進(jìn)行小球填充,使其恢復(fù)連續(xù)性及原來的網(wǎng)絡(luò)拓?fù)浣Y(jié)構(gòu);⑦ 血管網(wǎng)絡(luò)骨架中心線:將所有血管端點(diǎn)進(jìn)行標(biāo)記并保存,通過centerline generation功能對血管網(wǎng)絡(luò)骨架中心線進(jìn)行提取,并根據(jù)原始圖像對存在錯誤的骨架中心線進(jìn)行刪除;⑧ 網(wǎng)絡(luò)特征提取:分別通過feature analysis下的brain和vessel功能提取腦體積和血管體積參數(shù),通過Feature Analysis下Centerline中l(wèi)ength的功能提取血管長度和血管節(jié)段數(shù)參數(shù)。
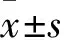
2 結(jié)果
2.1IFN-γ活動對斑馬魚腦血管發(fā)育的影響利用反義核苷酸技術(shù),于斑馬魚胚胎1-細(xì)胞期分別將crfb17 MO和對照MO顯微注射入胚胎動物極,下調(diào)斑馬魚IFN-γ受體crfb17的表達(dá),從而抑制內(nèi)源性IFN-γ的作用。由于斑馬魚腦血管發(fā)育時間窗為1.5~7 dpf,并在3 dpf時達(dá)到發(fā)育高峰[2],將3 dpf設(shè)為觀察起始點(diǎn)。又由于crfb17 MO組的胚胎在6~7 dpf時狀態(tài)普遍不佳,出現(xiàn)心包嚴(yán)重水腫、脊柱彎曲等癥狀,將5 dpf設(shè)為觀察結(jié)束點(diǎn)。分別在3 dpf和5 dpf這兩個時間點(diǎn)對幼魚腦血管進(jìn)行活體三維熒光共聚焦成像(圖1),結(jié)果顯示crfb17 MO組相較對照MO組,腦血管發(fā)育受到了明顯的抑制。
2.2中腦血管三維結(jié)構(gòu)定量分析利用軟件,對3 dpf時不同組別幼魚的中腦血管三維結(jié)構(gòu)進(jìn)行定量分析,crfb17 MO組中腦血管總長度(5.7±0.8) mm、血管節(jié)段數(shù)(36.5±6.3)、血管密度(1.65%±0.22%)都較對照MO組[(9.2±1.3) mm、(62.5±5.1)、(2.75%±0.18%)]顯著降低(t=5.606、7.804、9.409,P<0.001,進(jìn)一步證實(shí)了下調(diào)內(nèi)源性IFN-γ活動后,斑馬魚腦血管生成受到了抑制。見圖2。

圖1 crfb17 MO對斑馬魚腦血管發(fā)育的影響 ×26
2.3crfb17MO對斑馬魚軀干血管發(fā)育的影響由于中樞血管與外周血管發(fā)育的調(diào)控因素存在較大差異,為了探索IFN-γ對外周血管發(fā)育的影響,我們對斑馬魚胚胎進(jìn)行相同的操作,并對軀干血管發(fā)育進(jìn)行觀察。斑馬魚軀干血管發(fā)育時間窗大致為胚胎受精后24~30 h(hours post fertilization,hpf)[2],選取介于24~30 hpf的時間點(diǎn),即28 hpf(此時斑馬魚幼魚的自發(fā)抖動較24 hpf時明顯減少,利于成像)作為觀察軀干血管發(fā)育的起始點(diǎn),并于軀干血管發(fā)育結(jié)束后,即在31 hpf,再次進(jìn)行成像,作為觀察軀干血管發(fā)育的結(jié)束點(diǎn)。在正常斑馬魚軀干血管發(fā)育過程中,節(jié)間血管向背側(cè)生長,并通過背側(cè)縱向吻合血管(dorsal longitudinal anastomotic vessel,DLAV)相連接。圖3顯示,在28 hpf時,對照MO組已有少量DLAV連接相鄰血管,至31 hpf時,所有DLAV已經(jīng)長成并連接成管。而在crfb17 MO組中,從28 hpf至31 hpf,DLAV都沒有發(fā)育完全,說明下調(diào)crfb17后,斑馬魚軀干血管的發(fā)育受到了明顯抑制。
3 討論
本研究顯示,下調(diào)斑馬魚IFN-γ受體crfb17后,斑馬魚腦血管和軀干血管的發(fā)育受到明顯抑制;對腦血管進(jìn)一步的定量分析表明,3 dpf時crfb17 MO組中腦血管總長度、血管節(jié)段數(shù)、血管密度都較對照MO組顯著降低。
已經(jīng)有許多研究報(bào)道了免疫系統(tǒng)在血管生成中的重要作用。促炎性細(xì)胞因子如白介素6、白介素8和白介素1β等已被報(bào)道具有促進(jìn)血管生成的作用[3]。然而,IFN-γ一直被認(rèn)為是抑制血管生成的細(xì)胞因子[4-5]。IFN-γ對血管內(nèi)皮作用的研究主要基于腫瘤背景,IFN-γ不僅通過作用于腫瘤基質(zhì)細(xì)胞[6-7],而且直接作用于血管內(nèi)皮細(xì)胞[8-9],達(dá)到抑制血管生長從而抑制腫瘤生長的效果。然而,也有相當(dāng)一部分研究顯示,IFN-γ具有促血管生成的作用。Trompezinski et al[10]和Liu et al[11]分別發(fā)現(xiàn)IFN-γ能促進(jìn)人角質(zhì)細(xì)胞和視網(wǎng)膜色素上皮細(xì)胞釋放血管內(nèi)皮生長因子從而促進(jìn)血管生成。IFN-γ由免疫細(xì)胞產(chǎn)生,可以誘導(dǎo)巨噬細(xì)胞的活化,而Jetten et al[12]通過在小鼠腹腔血管生長基膜實(shí)驗(yàn)發(fā)現(xiàn),M2型巨噬細(xì)胞能夠增加內(nèi)皮細(xì)胞的數(shù)量和管狀結(jié)構(gòu)的形成。該研究顯示,抑制內(nèi)源性IFN-γ活動,斑馬魚的血管發(fā)育受到了明顯抑制,提示IFN-γ對斑馬魚血管發(fā)育的促進(jìn)作用。

圖2 3 dpf斑馬魚中腦血管三維結(jié)構(gòu)定量分析A:斑馬魚中腦血管三維骨架圖;B-D:斑馬魚中腦血管總長度、血管節(jié)段數(shù)、血管密度比較;與對照MO組相比:***P<0.001

圖3 crfb17 MO對斑馬魚軀干血管發(fā)育的影響 ×20紅色箭頭:指示對照MO組中已發(fā)育成形的DLAV
斑馬魚的基因組中含有十個編碼Ⅱ型細(xì)胞因子多肽的基因,其中兩個與哺乳動物IFN-γ最相關(guān)的基因?yàn)镮FN-γ1和IFN-γ2。其中IFN-γ1自斑馬魚胚胎0.5 hpf后就有持續(xù)的表達(dá)[13]。而ifng1-1和ifng1-2基因所表達(dá)蛋白的相應(yīng)受體皆為crfb17。IFN-γ在斑馬魚的早期發(fā)育中也起重要作用,IFN-γ已被報(bào)道參與斑馬魚胚胎早期膽道發(fā)育[14]和造血干細(xì)胞的形成[7]。
本研究顯示,通過抑制斑馬魚胚胎期內(nèi)源性IFN-γ的作用,斑馬魚的腦血管和軀干血管發(fā)育都受到了嚴(yán)重影響。此外,下調(diào)crfb17后,3 dpf斑馬魚腦血管發(fā)育受到抑制,而5 dpf時腦血管發(fā)育受抑制的現(xiàn)象更為明顯。由于3 dpf是斑馬魚腦血管發(fā)育的關(guān)鍵時期,胚胎腦血管發(fā)育在此時已受到抑制,另外,IFN-γ已被報(bào)道與斑馬魚胚胎早期胚胎發(fā)育中的重要作用[7,14],加上crfb17 MO的持續(xù)作用,導(dǎo)致5 dpf時腦血管發(fā)育受抑制的現(xiàn)象比3 dpf時更明顯,進(jìn)一步反映了內(nèi)源性IFN-γ在長時程的腦血管發(fā)育過程中持續(xù)的重要作用。雖然大部分體外實(shí)驗(yàn)表明,外源性給予IFN-γ能抑制內(nèi)皮細(xì)胞的增殖,也有報(bào)道證明,低劑量IFN-γ能誘導(dǎo)細(xì)胞增殖而高劑量則反之[15]。結(jié)合前述研究背景,提出假說,生理性IFN-γ對血管發(fā)育具有促進(jìn)作用,而在病理狀態(tài)下,炎性IFN-γ可通過抑制血管的生長而達(dá)到控制病灶發(fā)展的作用。
IFN-γ作為促炎細(xì)胞因子,具有抗病毒、免疫調(diào)節(jié)及抗腫瘤等特性。在腫瘤研究中IFN-γ起抑制血管生長的作用,而也有其他研究報(bào)道IFN-γ促血管生成的作用。本研究顯示,下調(diào)斑馬魚IFN-γ受體crfb17后,斑馬魚腦血管和軀干血管的發(fā)育受到明顯抑制,提示內(nèi)源性IFN-γ對斑馬魚血管發(fā)育中的促進(jìn)作用。該研究首次通過在體實(shí)驗(yàn)表明內(nèi)源性IFN-γ在血管發(fā)育階段對血管生成的促進(jìn)作用,期待后續(xù)進(jìn)一步的在體實(shí)驗(yàn)來證明內(nèi)源性IFN-γ對血管生長的調(diào)節(jié)作用。
