普通小麥品種陜農(nóng)33矮稈突變體的矮化效應(yīng)分析
2018-10-09 02:33:56趙秋實(shí)李倩倩王超杰蔣宏寶耿皆飛劉錄祥張小燕謝彥周王成社
麥類作物學(xué)報(bào) 2018年9期
趙秋實(shí),李倩倩,王超杰,蔣宏寶,耿皆飛,楊 媛,劉錄祥,張小燕,謝彥周,王成社
(1.西北農(nóng)林科技大學(xué)農(nóng)學(xué)院,旱區(qū)作物逆境生物學(xué)國家重點(diǎn)實(shí)驗(yàn)室,陜西楊凌 712100;2.中國農(nóng)業(yè)科學(xué)院作物科學(xué)研究所,北京 100081)
小麥?zhǔn)鞘澜缟献钪匾募Z食作物之一,分布廣泛,消費(fèi)量大,在我國是僅次于玉米和水稻的第三大糧食作物[1]。株高是影響小麥產(chǎn)量的關(guān)鍵因素之一,已成為小麥高產(chǎn)育種所需改良的重要目標(biāo)性狀。矮化育種是現(xiàn)代小麥高產(chǎn)育種的一個重要途徑,20世紀(jì)60年代的“綠色革命”正是基于矮化育種使小麥的產(chǎn)量得到了很大的提高[2-3]。由于矮稈小麥在高產(chǎn)、灌溉和抗倒伏方面的優(yōu)勢,在大部分地區(qū)尤其是在半干旱地區(qū),其已逐步取代普通的高稈小麥[4-5]。目前在育種中應(yīng)用最廣泛的矮稈基因有 Rht-B1b(Rht1)、 Rht-D1b(Rht2)、 Rht8和 Rht9;其中, Rht-B1b和 Rht-D1b來源于農(nóng)林10, Rht8和 Rht9來源于Akakomugi[6-9]。目前小麥育種中矮稈基因的來源比較單一。在我國小麥品種中, Rht-B1b(Rht1)基因的平均分布頻率為24.3%, Rht-D1b(Rht2)基因的平均分布頻率更高,為46.9%[10];我國小麥中約42.3%的品種含有 Rht8基因[11];楊芳萍等[12]對世界小麥主產(chǎn)區(qū)及Guedira等[13]、孫樹貴[14]等對美國小麥的矮稈基因來源進(jìn)行分析,也發(fā)現(xiàn)當(dāng)前矮稈基因的利用較為單一的情況。因此,挖掘創(chuàng)制新的矮源,研究新的矮稈突變體,有助于矮稈基因在小麥遺傳育種上的應(yīng)用并促進(jìn)小麥產(chǎn)量的提高。
矮稈突變體作為研究矮稈基因和獲取新矮源的基礎(chǔ)材料已用于水稻[15]、玉米[16]、小麥[17]、大豆[18]、大麥[19]、棉花[20]等作物的遺傳育種,以對水稻矮稈突變體的研究最為深入和廣泛。目前對于小麥矮稈突變體的研究很多。研究發(fā)現(xiàn),航天誘變的小麥矮稈突變體植株矮化的重要原因是倒一節(jié)節(jié)間縱向細(xì)胞數(shù)目的減少和長度的縮短[21];小麥矮稈突變體矮128屬于赤霉素(GA)不敏感型突變體,其矮稈性狀受一對隱性基因控制[22];經(jīng)EMS(甲基磺酸乙酯)誘變得到的小麥矮稈突變體NM9對GA不敏感,其莖稈GA含量極顯著高于野生型,從而抑制植株伸長[23];輻射誘變得到的小麥矮稈突變體的矮稈基因無不良基因的多效性,可以作為矮源在育種中利用[24]。雖然目前對小麥矮稈突變體的研究取得了一定的進(jìn)展,但是目前還是不能滿足育種的需要。
本試驗(yàn)選用經(jīng)NaN3(疊氮化鈉)處理小麥優(yōu)質(zhì)強(qiáng)筋品種陜農(nóng)33得到的13個性狀穩(wěn)定的矮稈突變體為材料[25],進(jìn)行農(nóng)藝性狀、赤霉素敏感性、內(nèi)源激素含量、莖稈細(xì)胞學(xué)和矮稈基因檢測和分析,以期初步了解這些矮稈突變體的矮化原因,為其在遺傳育種中的應(yīng)用奠定基礎(chǔ)。
1 材料與方法
1.1 材 料
主要試驗(yàn)材料為小麥優(yōu)良品種陜農(nóng)33及陜農(nóng)33經(jīng)NaN3誘變得到的13個性狀穩(wěn)定的矮稈突變體,中國春和新麥18作為部分試驗(yàn)對照材料;13個矮稈突變體以田間編號命名為D20、I49、J79、G50、D2、G34、A62、E48-1、A96、B19、F27、I25、H96。所有材料種植于西北農(nóng)林科技大學(xué)北校區(qū)試驗(yàn)田,株距10 cm,行長1 m,行寬30 cm。
1.2 DNA提取
在小麥三葉期,用CTAB法[26]提取基因組DNA,并用1%的瓊脂糖凝膠電泳檢測是否存在DNA降解和RNA污染。
1.3 農(nóng)藝性狀調(diào)查
在小麥成熟期分別調(diào)查陜農(nóng)33及其矮稈突變體的株高、穗長及各莖節(jié)長度、穗數(shù)、穗粒數(shù)、每穗小穗數(shù)、千粒重等性狀,并對野生型和矮稈突變體株高對比拍照。
1.4 細(xì)胞學(xué)觀察
在小麥抽穗后取穗下節(jié)作為材料,參考《植物制片技術(shù)》[27]中的方法,并稍作修改,在穗下節(jié)的中部取5 mm制作組織切片,經(jīng)過固定、初染、沖洗、脫水、透明、浸蠟、包埋、切片、脫蠟、復(fù)水、復(fù)染、透明、封固等過程制作石蠟切片,切片在32 ℃左右烘干,經(jīng)番紅固綠染色,在ZEISS(Imager M2)顯微鏡下觀察,用測微尺測量細(xì)胞長度和稈壁厚度,記錄100 μm2內(nèi)的細(xì)胞數(shù)目,并進(jìn)行統(tǒng)計(jì)分析。
1.5 對外源GA3反應(yīng)的測定
外源GA3苗期處理參照Gale[28]和Worland[29]等的方法,并稍作改動。分別將種子放在濕潤的培養(yǎng)基中,在4 ℃環(huán)境下萌發(fā)。兩天后,將發(fā)芽一致的種子種植到花盆中。隨后將花盆全部放在20 ℃、全黑暗的生長箱中,每天定量澆灌不同濃度的GA3。當(dāng)植株進(jìn)入三葉期,開始測量植株的第一片葉長度和胚芽鞘長度,每個濃度測10株。試驗(yàn)中每個材料設(shè)4個GA3濃度處理(0、20、50和100 mg·L-1),每個材料每次處理10株小麥苗。最后,將所得結(jié)果用SPSS 19.0(http:www.spss.com)進(jìn)行單因素方差分析,判別植株的GA反應(yīng)型。
1.6 內(nèi)源激素含量測定
取小麥抽穗期穗下節(jié)中部,分別用赤霉素(GA3)、細(xì)胞分裂素(CTK)、生長素(IAA)、油菜素內(nèi)酯(BR)激素試劑盒測定陜農(nóng)33及矮稈突變體的內(nèi)源激素含量(試劑盒由江蘇科晶生物科技有限公司提供)。
1.7 分子標(biāo)記檢測矮稈基因 Rht-B1b和 Rht-D1b
Rht-B1b和 Rht-D1b引物序列按照Ellis等[30]提供的序列設(shè)計(jì)。PCR引物序列由上海生工生物技術(shù)有限公司合成。其引物序列如下:
BF:5′-GGTAGGGAGGCGAGAGGCGAG-3′;
DF:5′-CGCGCAATTATTGGCCAGAGATAG-3′;
DF2:5′-GGCAAGCAAAAGCTTCGCG-3′;
MR1:5′-CATCCCCCATGGCCATCTCGAGCTA-3′;
WR1:5′-CATCCCCATGGCCATCTCGAGCTG-3′;
MR2:5′-CCCCATGGCCATCTCGAGCTGCAT-3′;
WR2:5′-GGCCATCTCGAGCTGCAC-3′。
Rht1基因位點(diǎn)野生型( Rht-B1a)和突變型 ( Rht-B1b)的特異性引物中,引物BF與WR1用于檢測Rht-B1a基因,BF與MR1用于檢測 Rht-B1b基因;Rht2基因位點(diǎn)野生型(Rht-D1a)和突變型( Rht-D1b)的特異性引物中,引物DF2與WR2用于檢測Rht-D1a基因,DF與MR2用于檢測 Rht-D1b基因。
Rht1檢測的PCR反應(yīng)條件:95 ℃,10 min;94 ℃變性30 s、65 ℃退火30 s、72 ℃延伸30 s,32個循環(huán);最后72 ℃延伸5 min;Rht2檢測的PCR反應(yīng)條件:95 ℃,10 min;94 ℃變性30 s、64 ℃退火30 s、72 ℃延伸10 s,43個循環(huán);最后72 ℃延伸 5 min。Rht1基因和Rht2基因的擴(kuò)增產(chǎn)物以2%的瓊脂糖凝膠電泳檢測,緩沖體系均為1×TAE溶液,100 V 電壓下電泳,溴化乙錠染色,凝膠成像儀下觀察并照相。
2 結(jié)果與分析
2.1 矮稈突變體的主要農(nóng)藝特征
野生型陜農(nóng)33株高為79.5 cm,矮稈突變體的株高為43.4~66.1 cm,這些矮稈突變體株高較陜農(nóng)33均有極顯著的降低(圖1和表1)。13個矮稈突變體千粒重較陜農(nóng)33均減少,其中11個材料的變化達(dá)到極顯著水平。有6個矮稈突變體的單株產(chǎn)量較陜農(nóng)33增加,其中D20增產(chǎn)極顯著;7個材料的單株產(chǎn)量較陜農(nóng)33減少,其中4個材料極顯著減產(chǎn)(表1)。矮稈突變體的其他農(nóng)藝性狀較陜農(nóng)33有降也有升,表明矮稈突變連鎖了一些不良農(nóng)藝性狀突變。在這些突變材料中D20、I49、J79和A96農(nóng)藝性狀綜合表現(xiàn)相對較好,可作為育種中間材料。
2.2 矮稈突變體的節(jié)間長度
小麥株高的降低通常是由于節(jié)間的縮短或者是節(jié)間數(shù)的減少導(dǎo)致。通過測定分析,與野生型陜農(nóng)33相比,這些矮稈材料的節(jié)間數(shù)沒有發(fā)生變化,加上穗下節(jié)一共都是5個節(jié)間;矮稈突變體株高的降低主要是由于各個節(jié)間的縮短所致,其中穗下節(jié)和第四節(jié)間縮短較為明顯。13個矮稈突變體的各節(jié)間均較野生型陜農(nóng)33縮短,且大部分變化顯著或極顯著,降稈效應(yīng)較大(表2)。
2.3 矮稈突變體的穗下節(jié)間細(xì)胞學(xué)顯微觀察結(jié)果
小麥株高的變化通常伴隨著莖稈細(xì)胞數(shù)目或者大小的變化。對穗下節(jié)進(jìn)行石蠟切片觀察發(fā)現(xiàn),矮稈突變體較陜農(nóng)33穗下節(jié)維管束有增多的趨勢(圖2a、圖2c、圖2e);9個矮稈突變體稈壁厚度比野生型陜農(nóng)33有所減少,其余4個矮稈突變體稈壁厚度有所增加;矮稈突變體的穗下節(jié)細(xì)胞長度較野生型陜農(nóng)33極顯著縮短,同時穗下節(jié)單位面積內(nèi)的細(xì)胞數(shù)目增加,表明其細(xì)胞變小(圖2、表3)。由此可以推斷,突變體株高的降低是由于莖稈細(xì)胞長度的減少和細(xì)胞變小共同造成的,而細(xì)胞長度的減少是主要因素。

SN33:陜農(nóng)33。SN33:Shaannong 33.

表1 矮稈突變體和陜農(nóng)33主要農(nóng)藝性狀的比較Table 1 Comparison of main agronomic traits of dwarf mutants and Shaannong 33
2.4 矮稈突變體的對外源GA3的反應(yīng)
根據(jù)赤霉素對幼苗形態(tài)的影響,可將矮稈材料劃分為赤霉素敏感型和赤霉素不敏感型。與不施GA3處理(0 mg·L-1)相比,經(jīng)不同濃度的GA3處理后,陜農(nóng)33和G50、D2、G34、A62、E48-1、A96、B19、H96的胚芽鞘長度均沒有發(fā)生顯著變化(表4),應(yīng)屬于赤霉素不敏感型;I49、J79、D20、F27、I25的胚芽鞘長度在個別的GA3濃度下有顯著變化,應(yīng)屬于赤霉素弱敏感型(表4)。中國春(赤霉素敏感型)的胚芽鞘長度隨GA3濃度的升高而增加,新麥18(赤霉素不敏感型)則變化不明顯(表4)。
2.5 矮稈突變體的內(nèi)源激素含量差異
與陜農(nóng)33相比,12個矮稈突變體穗下節(jié)的GA含量增加,其中7個突變體的變化達(dá)到極顯著水平,只有1個有微弱減少(圖3);矮稈突變體CTK含量均降低,其中有12個材料的變化極顯著降低(圖4);10個矮稈突變體的IAA含量有所升高,3個矮稈材料有所降低(圖5);矮稈突變體的BR含量變化不一,其中5個矮稈材料有所升高,8個矮稈材料有所下降(圖6)。這說明矮稈突變體株高的降低伴隨著內(nèi)源激素的變化。結(jié)合外源激素測定結(jié)果分析,這些矮稈材料的矮化可能與內(nèi)源激素代謝的變化密切相關(guān)。

表2 矮稈突變體和陜農(nóng)33的節(jié)間長度Table 2 Internode length of dwarf mutants and Shaannong 33 cm

表3 矮稈突變體和陜農(nóng)33莖稈特征的統(tǒng)計(jì)分析Table 3 Statistical analyses of stem characteristics of dwarf mutants and Shaannong 33

a、b:陜農(nóng)33穗下節(jié)橫切、縱切;c、d:矮稈突變體材料D20穗下節(jié)橫切、縱切;e、f:矮稈突變體材料I49穗下節(jié)橫切、縱切;SV:小維管束;BV:大維管束。a and b:Transverse and longitudinal sections of the peduncle of Shaannong 33; c and d:Transverse and longitudinal sections of the peduncle of dwarf mutant D20; e and f:Transverse and longitudinal sections of the peduncle of dwarf mutant I49; SV:Small vessels; BV:Large vessels.
2.6 矮稈突變體的矮稈基因檢測結(jié)果
目前已發(fā)現(xiàn)并命名的矮稈基因大概有27個,本試驗(yàn)初步對在育種中應(yīng)用范圍較廣的2個矮稈基因 Rht-B1b(Rht1)和 Rht-D1b(Rht2)進(jìn)行檢測,以初步分析陜農(nóng)33和矮稈突變體是否含有這兩個在育種中應(yīng)用較廣的矮稈基因;利用Ellis[30]等設(shè)計(jì)的引物序列,檢測 Rht-B1b基因。如果小麥品種中具有Rht-B1a基因,用引物BF和WR1可以擴(kuò)增出一條237 bp的片段;如果具有 Rht-B1b基因,用引物BF和MR1也可以擴(kuò)增出一條237 bp的片段,因此同一品種的兩對引物PCR產(chǎn)物可以互補(bǔ)驗(yàn)證(圖7)。用引物DF2與WR2檢測Rht-D1a基因,DF與MR2檢測 Rht-D1b基因。如果小麥品種中具有Rht-D1a基因,可以擴(kuò)增出一條264 bp的片段;如果具有 Rht-D1b基因,則可以擴(kuò)增出一條254 bp的片段(圖8);在PCR反應(yīng)和凝膠電泳中,加入無 Rht-B1b和 Rht-D1b基因的CS作為對照。
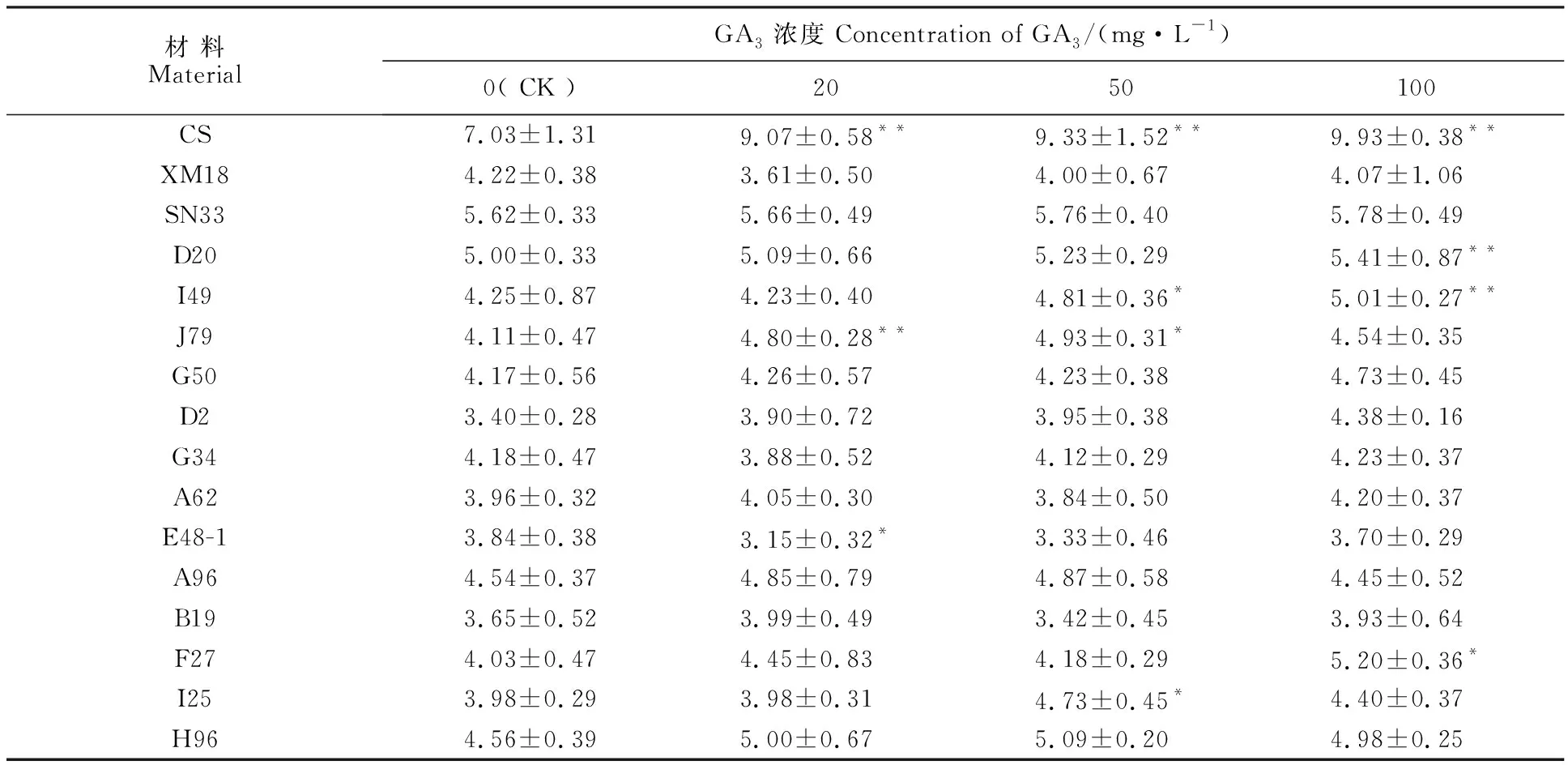
表4 外源赤霉素(GA3)對矮稈突變體和陜農(nóng)33幼苗胚芽鞘長度的影響Table 4 Effects of gibberellin acid(GA3) on the coleoptile length of dwarf mutants and Shaannong 33 at seedling stage

*和**分別表示矮稈材料和陜農(nóng)33相比較在0.05和0.01水平上差異顯著。SN33:陜農(nóng)33。圖4~圖6同。* and ** mean significant difference between dwarf mutants and Shaanong 33 at 0.05 and 0.01 levels, respectively. SN33:Shaannong 33. The same in figure 4-figure 6.

圖4 矮稈突變體和陜農(nóng)33穗下節(jié)內(nèi)源CTK含量

圖5 矮稈突變體和陜農(nóng)33穗下節(jié)內(nèi)源IAA含量

圖6 矮稈突變體和陜農(nóng)33穗下節(jié)內(nèi)源BR含量

M:2 000 bp DNA ladder;1:中國春;2:陜農(nóng)33;3:D20;4:I49;5:D2;6:I25;a: Rht-B1a 基因特異性引物的PCR擴(kuò)增;b: Rht-B1b 基因特異性引物的PCR擴(kuò)增。M:2000 bp DNA ladder;1:Chinese Spring;2:Shaannong 33;3:D20;4:I49;5:D2;6:I25;a:PCR amplification of Rht-B1a gene-specific primers;b:PCR amplification of Rht-B1b gene-specific primers.
經(jīng)PCR分析,矮稈突變體材料和陜農(nóng)33中均擴(kuò)增出254 bp片段(表5),說明其均含有 Rht-D1b基因;而在D2和I25中利用引物BF和MR1擴(kuò)增出237 bp片段(圖8),說明這兩個矮稈突變體材料含有 Rht-B1b基因。這表明矮稈突變體材料和陜農(nóng)33均含有矮稈基因 Rht-D1b,矮稈突變體材料的株高降低表明其體內(nèi)可能還含有其他致矮相關(guān)的基因。

M:DNA ladder 2000;1:中國春;2:陜農(nóng)33;3:D20;4:I49;5:D2;6:I25; a:Rht-D1a基因特異性引物的PCR擴(kuò)增,b: Rht-D1b 基因特異性引物的PCR擴(kuò)增。 M:DNA ladder 2000; 1:Chinese Spring; 2:Shaannong 33; 3:D20; 4:I49; 5:D2; 6:I25; a:PCR amplification with Rht-D1a gene-specific primers; b:PCR amplification with Rht-D1b gene-specific primers.
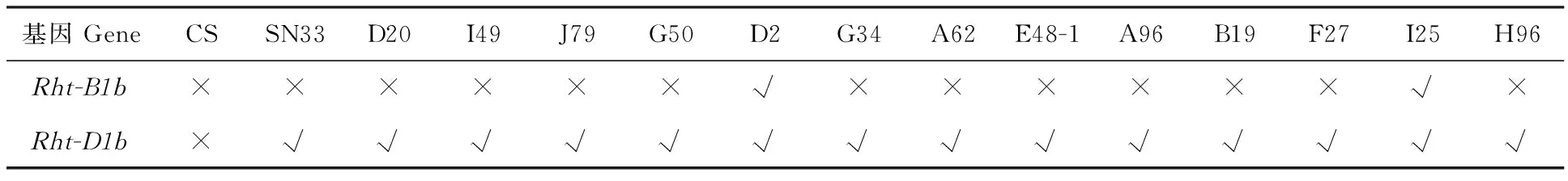
表5 矮稈基因檢測結(jié)果統(tǒng)計(jì)Table 5 Result of dwarf gene test
3 討 論
自第一次“綠色革命”以來,矮稈材料的利用使小麥的單產(chǎn)和總產(chǎn)得到了大幅提高,株高也成為小麥高產(chǎn)育種的重要目標(biāo)性狀。但目前應(yīng)用于小麥育種的矮稈基因的來源比較單一,因此需要進(jìn)一步創(chuàng)制和研究新的小麥矮源。
小麥的矮稈基因主要有赤霉素敏感型和赤霉素不敏感型兩種類型。研究表明,赤霉素敏感型的矮稈基因?qū)χ旮叩慕刀捘芰^弱,但是其對重要農(nóng)藝性狀的影響要小一些,適合于干旱和半干旱地區(qū)育種利用。赤霉素不敏感型矮稈基因降稈能力較強(qiáng),但同時也會影響到一些重要的農(nóng)藝性狀如胚芽鞘、籽粒大小等等[31-33]。本研究中的矮稈材料為赤霉素不敏感型或弱敏感型,降稈效果較明顯,但同時有一些不良性狀的連鎖(如千粒重降低、單株產(chǎn)量的降低等),這與前人研究結(jié)果一致。這些矮稈材料中綜合農(nóng)藝性狀相對較好的D20、I49、J79和A96可作為育種中間材料進(jìn)行利用。
紫外線[34]、EMS[35]、NaN3[25]等誘變技術(shù)都能使作物產(chǎn)生矮稈突變效應(yīng),且株高的致矮機(jī)理也不盡相同。水稻矮稈突變體MU101的節(jié)間數(shù)較野生型減少2個,各節(jié)間也有所縮短,細(xì)胞數(shù)目的減少是矮變的主要原因[15];而航天誘變的小麥矮稈突變體的矮化結(jié)果主要?dú)w因于各節(jié)間縮短、細(xì)胞長度的變小和細(xì)胞數(shù)目的減少[21]。本研究中,突變體的致矮原因與前人研究結(jié)果有所不同,矮化主要取決于各節(jié)間的縮短,而節(jié)間數(shù)目不變,其中穗下節(jié)和第四節(jié)間的縮短降低株高的效應(yīng)最大。細(xì)胞學(xué)觀察發(fā)現(xiàn),穗下節(jié)的細(xì)胞長度下降,體積變小,共同導(dǎo)致了矮稈突變體的矮化,其中矮稈材料的細(xì)胞長度有極顯著變短,是株高矮化的主要原因。
植物內(nèi)源激素通過調(diào)節(jié)或改變生理活動,對植物的生長發(fā)育進(jìn)程有著很重要的調(diào)控作用[36]。在已發(fā)現(xiàn)的植物激素中,赤霉素(GA)、細(xì)胞分裂素(CTK)、生長素(IAA)、脫落酸(ABA)和油菜素內(nèi)酯(BR)均參與了植物的株高建成[37-41]。研究表明,赤霉素敏感型矮稈突變體內(nèi)源赤霉素合成受阻,內(nèi)源赤霉素缺乏,導(dǎo)致植株變矮[42];而GA不敏感型矮稈突變體內(nèi)源GA含量變化不大,甚至?xí)@著高于野生型,GA的積累會抑制生長,導(dǎo)致株高降低[43]。小麥矮稈材料NM9對GA的不敏感,其莖稈GA含量較高,使植株矮化[23]。本研究中,13個矮稈材料經(jīng)鑒定為GA不敏感或弱敏感型,12個矮稈突變體莖稈GA含量較野生型陜農(nóng)33有所增加,只有1個微弱的降低,并且有9個顯著增加,這與前人研究結(jié)果一致,推測矮稈材料的GA轉(zhuǎn)導(dǎo)途徑受阻,導(dǎo)致GA積累,抑制了植株生長,使株高降低。CTK主要作用是促進(jìn)細(xì)胞分裂和細(xì)胞體積擴(kuò)大。本研究中矮稈材料的CTK含量均降低,其中有12個達(dá)到極顯著的降低,說明CTK的降低可能也影響了植株的生長發(fā)育。IAA和BR主要作用是促進(jìn)細(xì)胞的伸長。在小麥矮稈材料NM9中IAA含量與野生型相比無顯著差異[23],而本研究中矮稈材料IAA和BR含量相較于陜農(nóng)33有升有降,可能是由于每個矮稈突變體都是獨(dú)立的突變,它們的突變背景有所不同,在致矮突變的同時也引起了其他方面不同的突變,致使它們的內(nèi)源IAA和BR表達(dá)水平有所不同。
目前育種中應(yīng)用的矮稈基因來源相對比較單一,應(yīng)用最廣泛的有 Rht-B1b(Rht1)、 Rht-D1b(Rht2)、 Rht8和 Rht9;楊松杰等[10]研究發(fā)現(xiàn), Rht-B1b(Rht1)基因在我國小麥中的平均分布頻率為24.3%, Rht-D1b(Rht2)基因在我國小麥中的平均分布頻率為46.9%;本研究檢測分析發(fā)現(xiàn),13個矮稈突變體和陜農(nóng)33均含有 Rht-D1b基因,有兩個矮稈材料D2和I25含有 Rht-B1b基因,這兩個材料的株高也比較,但其他矮化材料,卻不含有這個基因,因此推測矮稈材料可能還含有突變的致矮基因,后期需要進(jìn)一步對這些矮稈材料進(jìn)行遺傳模式的分析和主效矮稈基因的定位,從而豐富小麥的矮源并使矮稈基因得到有效利用。
