通過定點突變提高納豆激酶的酶活及熱穩定性
2018-10-17 11:16:20趙菡周麗周哲敏
食品與發酵工業 2018年9期
趙菡,周麗,周哲敏
(江南大學 生物工程學院,工業生物技術教育部重點實驗室,江蘇 無錫,214122)
枯草桿菌蛋白酶是絲氨酸蛋白酶的第二大家族,已經作為模式酶被廣泛研究[1],研究內容主要包括改造其催化活性[2-3]、底物結合特異性[4]以及穩定性[5]等。納豆激酶(Nattokinase,NK,EC3.4.21.62)是由日本傳統食品納豆在發酵過程中產生的一種枯草桿菌蛋白酶,其編碼基因為aprN[6],具有保守的催化活性中心(Asp32, His64 和 Ser221)。納豆激酶不僅可以通過激活纖溶酶原將其轉變為纖溶酶[7],間接溶解纖維蛋白,還可以直接作用于交聯的纖維蛋白[8],因而具有很強的溶栓功效。相較于市售的溶栓藥物(鏈激酶、尿激酶以及蛇毒纖溶酶等),納豆激酶還有半衰期長、安全、副作用小以及成本低等優點,因此將其開發為治療心腦血管疾病的口服藥物,具有廣闊的應用前景[9]。然而野生型納豆激酶的酶活較低,熱穩定性較差,尤其當溫度超過60 ℃時,酶分子會迅速失活,不利于其在工藝生產高溫環節保證酶活,因此不能滿足大規模生產[10]。本文選用一種高效的方法以提高納豆激酶的活性以及熱穩定性。
在蛋白質中,翻譯后的脫酰胺過程將天冬酰胺(Asn)和谷氨酰胺(Gln)轉化為帶負電的天冬氨酸(Asp)和谷氨酸(Glu),可能改變蛋白質的電荷網絡進而影響酶的活性、最適pH值和穩定性等[11]。因此,可通過替換酶分子中的Asn和Gln改造酶分子的性質。如KATO將蛋清溶菌酶中2個Asn突變為Asp后,最適pH值向酸性偏移了0.5個單位[12];JAKOB通過改造蛋白酶BgAP表面的Asn和Gln,使其在pH值8.5下的熱穩定性提高了2.5 ℃[13]。研究發現,蛋白質表面氨基酸的改造可以改變酶的性質。例如,KHURANA將表面的疏水氨基酸突變為親水氨基酸提高了脂肪酶的穩定性[14]MICHAEL通過替換酶分子表面的76位和218位氨基酸提高了枯草蛋白酶BPN’的穩定性[15]。此外,表面電荷的改造也可提高酶的活性[16]和穩定性等。如,JAOUADI通過優化絲氨酸蛋白酶表面電荷與電荷之間的相互作用提高了其熱穩定性[17]。然而,許多研究集中于改造酶活性中心附近的氨基酸,而遠離活性中心的位點也可能影響酶的性質,如,MIYAZAKI采用定向進化方法篩選得到熱穩定性及酶活提高的突變體位于遠離酶活性中心的loop環上[18]。
綜合以上研究,為了高效選擇突變位點,設定以下選點原則:選擇表面的Asn和Gln;為非保守殘基;遠離酶的活性中心。最終選擇8個位點,分別將其突變為帶負電的Asp和Glu,通過篩選得到酶活和熱穩定性提高的突變株,為納豆激酶的工業應用奠定了基礎。
1 材料與方法
1.1 菌株,質粒以及培養條件
EscherichiacoliJM109以及E.coliBL21 (DE3)分別用于質粒構建以及蛋白表達,由作者實驗室保藏。質粒pET-24a用作表達載體,將納豆激酶的成熟肽與前導肽基因序列克隆至pET-24a載體上(于限制性酶切位點XbaI 和BamHI之間),得到重組表達載體pET-24a-pro-NK(實驗室已有)[10]。LB培養基(10 g/L胰蛋白胨,5 g/L酵母提取物,10 g/L NaCl)用于質粒的構建與培養。自誘導培養基ZYM-5052用于納豆激酶的發酵表達[19]。培養基中卡那霉素的終質量濃度為50 μg/ mL。
1.2 主要材料與儀器
1 mL離心式蛋白純化空柱、含鎳樹脂:生工生物工程(上海)股份有限公司;對硝基苯胺(pNA)、四肽合成底物(N-succinyl-L-Ala-L-Ala-L-Pro-L-Phe-p-nitroanilide,suc-AAPF-pNA) :Sigma公司;PCR儀:BIO-RAD公司;UV-1800PC型紫外可見分光光度計:上海美譜達有限公司;pH計:梅特勒-托利多儀器(上海)有限公司;多功能酶標儀:Bioteck公司(美國)。
1.3 實驗方法
1.3.1 基因操作
以質粒pET-24a-pro-NK為模板采用定點突變(重疊延伸)方法構建突變體。所有突變體相關引物如表1所示。

表1 本研究所用引物序列Table 1 Oligonucleotide primers used in this study
PCR得到的基因產物于DpnI酶中37℃消化5 h以降解模板,然后轉化至E.coliJM109感受態細胞中并提取質粒,基因序列驗證正確后,轉化至E.coliBL 21(DE3)中,成功獲得陽性轉化子。
1.3.2 目的蛋白的表達純化
挑取單個轉化子接種于含卡那霉素的5 mL LB培養基中,37 ℃、200 r/min振蕩過夜培養。以2%的接種量將種子培養液轉接至100 mL自誘導培養基ZYM-5052中,37 ℃、200 r/min振蕩培養。當OD600值達到2左右時,溫度降至18 ℃,培養20 h,離心收集菌體,用10 mL結合緩沖液(100 mmol/L Tris-HCl pH 8.0,500 mmol/L NaCl,20 mmol/L咪唑)重懸菌體,并于冰上超聲破碎,收集破碎上清液,于4 ℃經親和層析鎳柱離心柱(將含鎳樹脂裝入1 mL離心空柱中)純化目的蛋白,采用含50~100 mmol/L咪唑洗脫緩沖液進行梯度洗脫,純酶液于100 mmol/L Tris-HCl pH 8.0緩沖液中過夜透析。采用Bradford法測定蛋白濃度,并將野生型以及突變體純酶液調節至相同的蛋白濃度。
1.3.3 酶活的測定[3]
在100 mmol/L Tris-HCl pH 8.0緩沖液(含0.1 mmol/L CaCl2)中,以人工合成的suc-AAPF-pNA為底物,以對硝基苯胺(pNA)為標準,在終體積為0.2 mL,25 ℃下反應3 min,根據405 nm 的吸收值測定生成pNA濃度。一個酶活力單位定義為:在25 ℃,pH值8.0,底物濃度為0.4 mmol/L時,1 min內,將1 μmol suc-AAPF-pNA轉化為1 μmolpNA所需的酶量。
1.3.4 最適溫度和最適pH值的測定
取適量純酶液溶解于100 mmol/L Tris-HCl(pH 8.0)緩沖液中,根據以上酶活測定方法在30、40、50、60、70 ℃下測定其活性,將最高的酶活力定義為100%。在不同pH值緩沖液條件下測定野生型酶和突變體酶的活性,將最高的酶活力定義為100%,pH值3.0~5.0采用50 mmol/L檸檬酸緩沖液,pH值6.0~8.0為50 mmol/L磷酸鹽緩沖液,pH值9.0~11.0為50 mmol/L碳酸鈉緩沖液(至少3次獨立實驗的平均值)。
1.3.5 熱穩定性和pH值穩定性的測定
取適量野生型和突變體純酶液于55 ℃中分別孵育(10~60 min),測定各個純酶液的半衰期(t1/2),將未處理的原始酶活定義為100%,并以相對酶活對時間作圖。取適量野生型和突變體純酶液在4 ℃下,不同的緩沖液(pH 3.0~5.0,50 mmol/L檸檬酸緩沖液,pH 6.0~8.0,50 mmol/L磷酸鹽緩沖液,pH 9.0~11.0,50 mmol/L碳酸鈉緩沖液)中孵育4 h,測定剩余酶活,將最高的酶活力定義為100%,并以相對酶活對pH值作圖(至少3次獨立實驗的平均值)。
1.3.6 動力學常數的測定
以人工合成的四肽suc-AAPF-pNA為底物,底物濃度范圍為0.05~1 mmol/L,在終體積為0.2 mL,25 ℃下反應3 min,測量suc-AAPF-pNA的初始反應速率,通過軟件GraphPad Prism 5擬合得到Km與Vmax值,根據kcat=Vmax/ [Enzyme],最后求得kcat及催化效率常數kcat/Km。
2 結果與分析
2.1 突變點的選擇
為提高納豆激酶的活性及穩定性,本文依照以下3個原則選擇突變點:利用GETAREA軟件篩選表面暴露程度>50%的Asn和Gln;位點遠離酶的活性中心(以減少對酶活的負面影響);為非保守氨基酸(圖1)。最終,構建了8個突變體N43D、Q59E、Q103E、N109D、 N118D、Q185E、Q206E、N218D(圖2)。

箭頭所指為突變位點圖1 NK與其他枯草桿菌蛋白酶的序列比對Fig.1 Sequence alignment of NK with other subtilisins

圖b由圖a旋轉一定角度得到;D32,H64,S221為催化活性中心位點圖2 突變位點在納豆激酶三維結構中的分布Fig.2 The distribution of mutants in NK three-dimensional structure
2.2 單點突變的初步篩選
為初步篩選熱穩定性提高的突變體,取野生型與突變體粗酶液于65 ℃處理30 min,測定剩余酶活,如圖3-a所示,N218D的熱穩定性有所提高,相較于WT其殘余酶活提高了15%左右。經過初篩,Q59E酶活顯著高于WT(圖3-b)。

a-熱穩定性篩選;b-酶活篩選圖3 單突變體初步篩選Fig.3 Preliminary screening of single mutants
2.3 組合突變與酶學性質表征
為進一步提高納豆激酶的綜合性質,將初步篩選得到的酶活提高突變體Q59E以及熱穩定性提高的突變體N218D進行組合,得到雙突變體Q59E-N218D。
為了準確表征各個突變體酶,將WT以及3個突變體(Q59E、N218D和Q59E-N218D)進行表達純化,取純酶液測定其酶學性質。如圖4-a和圖4-b,各個突變體酶的最適溫度和最適pH值并未發生偏移。如圖5-a,突變體Q59E的熱穩定性較差,而突變體N218D和Q59E-N218D的熱穩定性明顯提高。其中,N218D在55 ℃下的t1/2約為28 min(表2),相較于WT(約12 min)提高了1.3倍。組合突變體的熱穩定性進一步提高,其t1/2提高至33 min左右,為WT的2.75倍。通過對比野生型和突變體的pH值穩定性(圖5-b),發現Q59E和N218D的堿性穩定性較差,然而雙突變Q59E-N218D 在pH 5~7更為穩定,即耐酸性較WT有所提高。

a-最適溫度曲線;b-最適pH曲線圖4 最適反應條件的測定Fig.4 Determination of the optimum reaction conditions
2.4 動力學常數和比酶活的測定
為了研究催化活性,本文測定了突變體酶的動力學常數以及比酶活值(如表3)。突變體N218D 熱穩定性提高,可能是由于其結構剛性增強,然而代價是與底物的親和能力下降以及kcat的降低,導致催化效率較WT下降了約40%。相反,突變體Q59EKm值降低,說明與底物的親和能力增強,同時kcat提高了37%,因而催化效率顯著提高,約為WT的1.60倍。另外,Q59E的比酶活提高,為WT的1.54倍,與動力學參數相一致。然而,Q59E熱穩定性較差,說明酶活與熱穩定性之間存在一定的平衡,酶的催化活性更需要柔性的空間構象,而酶的熱穩定性則要求酶結構的剛性以抵御外界壓力的破壞。因此將Q59E引入N218D,雙突變體Q59E-N218D催化效率比單突變體N218D增強,同時熱穩定性明顯提高,與何孝天的研究相比[10](其提高納豆激酶熱穩定性的同時伴隨著催化活性的大幅度下降),本研究提供了綜合性質更好的突變株,為納豆激酶的工業應用奠定了基礎。所以組合突變可以一定程度上綜合單點突變的優勢獲得最佳平衡,從而得到更為有效的突變酶。
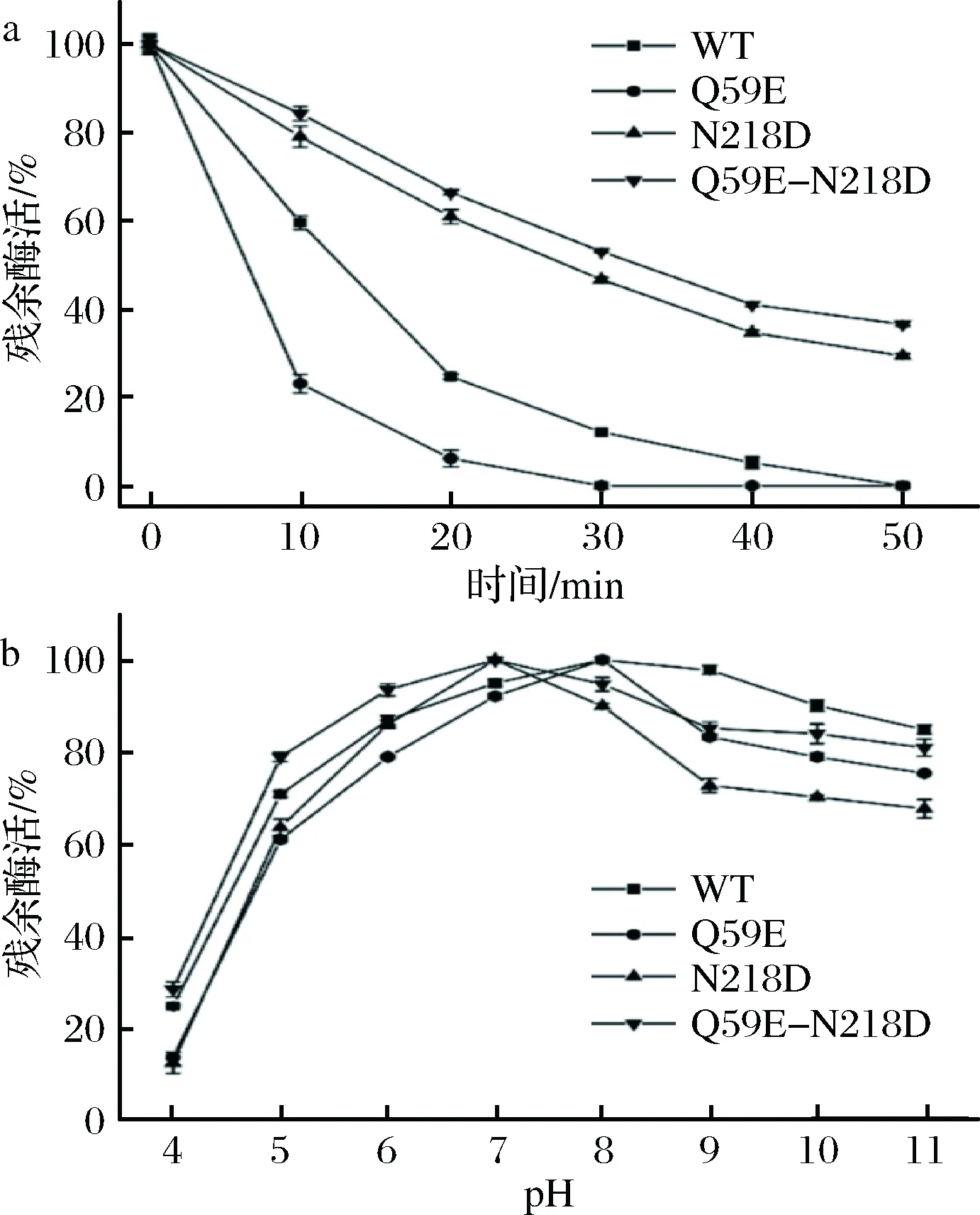
a-原始酶與突變體的熱穩定性(在55℃下殘余酶活隨時間變化)曲線;b-原始酶與突變體在不同pH下的穩定性曲線圖5 熱穩定性和pH穩定性的測定Fig.5 Determination of the thermal stability and pH stability
表2 野生型與突變體酶的半衰期值Table 2 Half-life of wild-type NK and mutants

酶半衰期(t1/2)/minwild-type11.90±0.59Q59E4.81±0.14N218D28.17±0.35Q59E-N218D32.38±0.98

表3 野生型酶與突變體酶的動力學參數以及比酶活Table 3 Kinetic parameters and specific activities of wild-type NK and mutants
3 討論
本文通過定點突變構建重組納豆激酶突變體,經過篩選得到催化活性顯著提高的突變體Q59E以及熱穩定性提高突變體N218D。組合突變Q59E-N218D的熱穩定性進一步提高,由野生型酶的12 min提高至33 min,且其比酶活達到與原始酶相似水平。
本文酶活提高突變體Q59E,在蛋白質表面引入了負電荷,可能通過遠距離靜電相互作用影響酶活性中心區域的結構,從而提高了納豆激酶的催化活性。因此,本文采用篩選突變體的方法可作為一種改善酶性質的有效策略,為其他酶的改造提供參考。
