miR-222促進家兔骨骼肌衛星細胞增殖
2018-10-19 10:04:22劉卜瑋毛旭東梁小虎鄭天豪何洪炳賴松家
西南農業學報 2018年9期
關鍵詞:檢測
劉卜瑋,楊 雪,2,毛旭東,梁小虎,鄭天豪,何洪炳,賴松家 *
(1. 四川農業大學畜禽遺傳資源發掘與創新利用四川省重點實驗室,四川 成都 611130; 2. 成都市農林科學院,四川 成都 611130)
【研究意義】高等動物中,骨骼肌衛星細胞(skeletal muscle satellite cells,SMSCs)附著在肌纖維的底膜上,正常情況下是靜止的單核細胞,當肌肉受到損傷后,能通過增殖和分化與原有的肌纖維融合形成新的肌纖維,對肌肉進行修復。SMSCs的增殖和分化受復雜的基因網絡多重調控,例如成肌調節因子(MRFs)家族的成肌分化因子D(MyoD)、MyoG、Myf4、Myf5和Myf6;它們和肌細胞增強因子2(MEF2)家族成員共同激活和肌肉特異性的基因轉錄,其中MyoG對骨骼肌衛星細胞分化的作用尤為突出,是分化的關鍵調控因子[1-2]。
MicroRNAs(miRNAs)是真核生物中長度18~24 bp的一類短鏈非編碼RNA,通過與靶基因的3端UTR區互補配對結合,不完全靶向可抑制該mRNA發揮作用,完全靶向則會降解該mRNA,從而實現在轉錄后水平抑制靶基因的表達[3]。【前人研究進展】miRNAs在肌肉的生長發育過程中也起著重要作用,如miR-1,miR-133a和miR-206是橫紋肌特異的miRNAs,在成肌細胞增殖分化過程中,miR-1、miR-133a和miR-206表達量升高,miR-1通過靶向組蛋白去乙酰化酶4(HDAC4)從而促進MEF2的表達,而MEF2又可以促進miR-1的表達,形成一個正反饋環,從而促進成肌細胞的分化和抑制成肌細胞的增殖[4]。而miR-133a和miR-206分別通過抑制血清應答因子(SRFs)和成肌抑制素基因(myostatin)的表達促進SMSCs增殖[5-6]。miR-222參與調控多種生物學過程,例如,在小鼠的心臟中過表達miR-222,可以通過激活mTOR信號通路抑制心肌的凋亡,同時抑制P27的表達從而誘導心臟心力衰竭[7];在平滑肌中,miR-222可以通過靶向PTEN/Akt通路中的關鍵因子抑制血管平滑肌自噬[8];miR-222能通過靶向P21和P27促進氣管平滑肌的增殖[9-10]。此外,更多的研究主要集中在miR-222對腫瘤的調節作用,研究表明miR-222能夠促進黑色素瘤[11]、宮頸癌[12]、結直腸癌[13]等腫瘤的增殖或轉移。有研究表明,在小鼠的成肌細胞系(C2C12細胞系)中,miR-222過表達可以促進成肌細胞的增殖和抑制成肌細胞的分化[14]。然而,目前沒有研究報道miR-222是否參與調控家兔骨骼肌衛星細胞的增殖。
我國是養兔大國,也是兔肉出口大國。家兔是一種十分重要的經濟動物,它既可以食用,也可以作為工業原料、觀賞動物、醫用模型等,因此家兔具有很大的研究價值。【本研究切入點】本試驗采用剛出生新西蘭兔骨骼肌衛星細胞為實驗材料,研究miR-222對SMSCs增殖的影響。【擬解決的關鍵問題】結果表明miR-222過表達促進SMSCs的增殖,miR-222抑制表達抑制SMSCs的增殖。
1 材料與方法
1.1 組織的采集和RNA的提取
所有的實驗動物均由四川農業大學科研兔場提供,分別采集6只(公母各3只)84日齡新西蘭兔的心、肝、脾、肺、腎、回腸和腿肌(半腱肌)組織樣各1 g左右,迅速放入液氮中保存。將樣品在液氮中研磨后,用Trizol法(TaKaRa,寶生物)提取出各樣本的總RNA備用。
1.2 RNA的抽提與反轉錄
總RNA和miRNAs的純度采用OD260nm/OD280nm和OD260nm/OD230nm比值測定,其完整性通過1.5 %瓊脂糖凝膠電泳評估。參照反轉錄試劑盒(Takara,Mir-XTM miRNA First-Strand Synthesis Kit 貨號:1604342A和Takara, PrimeScriptTM RT reagent Kit with gDNA Eraer 貨號AK4002)進行miRNA和mRNA的反轉錄,得到各自的cDNA,保存于-20 ℃冰箱。
1.3 實時螢光定量PCR(RT-qPCR)
miR-222的定量用U6(microRNA反轉錄試劑盒自帶)作為處理數據的內參基因,miR-222引物序列5’-agctacatctggctactgggt-3’,每個樣本3個重復孔;RT-qPCR定量程序:預變性95 ℃ 30 s;40個循環:95 ℃ 5 s,60 ℃ 25 s,熔解曲線:95 ℃ 10 s,65 ℃ 5 s,95 ℃ 5 s。mRNAs的定量體系:5 μl SYBR green I,0.4 μl上游引物,0.4 μl下游引物3.2 μl的去RNA,1 μl cDNA。本試驗所用引物均由成都擎科生物有限公司合成。
1.4 功能富集分析
使用TargetScan(http://www.targetscan.org/vert_71/)、miRBase(http://www.mirbase.org/)和miRWalk(http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/generetsys-self.html)對miR-222進行靶基因預測,選擇3個網站的預測交集作為miR-222潛在的靶基因,然后使用DAVID 6.8工具(https://david.ncifcrf.gov/)進行靶基因功能富集分析。
1.5 原代細胞培養
新生1 d的新西蘭兔脫頸處死后,取后腿半腱肌放入PBS中,仔細去除肌肉上面筋膜等組織,將肌肉塊剪碎后放入0.1 %的膠原蛋白Ⅰ中37 ℃水浴消化2 h,每10 min搖晃混勻1次;離心后用胰蛋白酶消化10 min,用生長培養基(含10 % FBS,1 %雙抗的DMEM高糖培養基)(GM)終止消化并用40 μm細胞篩網過濾,將濾液1000 r/min離心5 min,再將細胞重懸接種在培養瓶中。
1.6 miR-222過表達和抑制表達效率檢測
miR-222 mimics、miR-222 NC、miR-222 inhibitor和inhibitor NC由上海生工生物工程股份有限公司合成,序列如表1。
表1miR-222mimics、miR-222NC、miR-222inhibitor和inhibitorNC序列信息
Table 1 The sequence information of miR-222 mimics, miR-222 NC, miR-222 inhibitor and inhibitor
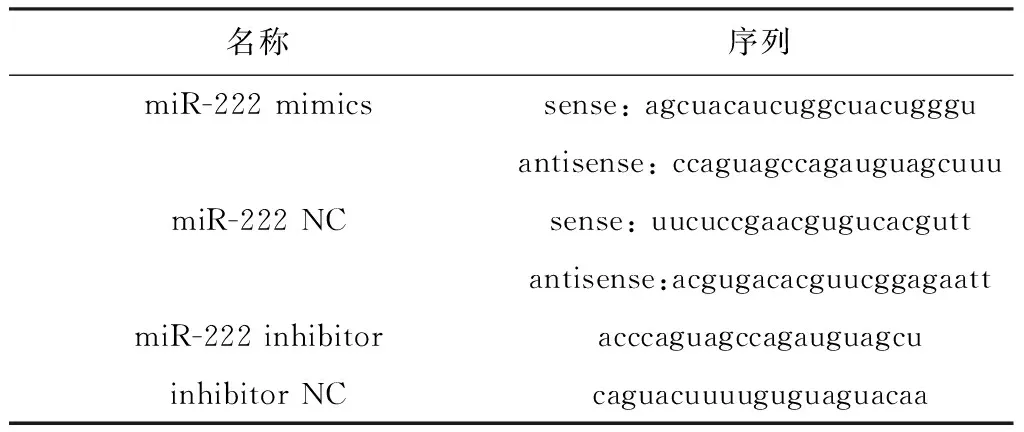
名稱序列miR-222 mimicssense: agcuacaucuggcuacuggguantisense: ccaguagccagauguagcuuumiR-222 NCsense: uucuccgaacgugucacguttantisense:acgugacacguucggagaattmiR-222 inhibitoracccaguagccagauguagcuinhibitor NCcaguacuuuuguguaguacaa
將SMSCs培養在6孔板中,當細胞密度達到80 %左右時,根據lipo3000(Thermo,貨號:L3000015)說明書將miR-222 mimics(濃度:50 nM)、miR-222 NC(濃度:50 nM)、miR-222 inhibitor(濃度:100 nM)和inhibitor NC(濃度:100 nM)進行轉染,每組3個重復孔,24 h后換液,48 h后,提取RNA進行反轉錄和定量分析miR-222的表達量,確定過表達和抑制表達效率。
1.7 細胞增殖檢測
CCK-8增殖檢測:將SMSCs按照1×104個/孔的密度接種進96孔板,待細胞密度達到80 %左右,根據lipo3000說明書將miR-222 mimics(濃度:50 nM)、miR-222 NC(濃度:50 nM)、miR-222 inhibitor(濃度:100 nM)和inhibitor NC(濃度:100 nM)進行轉染,每組8個重復,24 h后換成100 μl GM培養基,之后每48 h換液;檢測轉染后第24、48、74、96和120小時的細胞增殖情況,簡述如下,每個孔提前2 h加入10 μl CCK-8試劑(同仁,貨號ck04)放入37 ℃培養箱中孵育2 h,在450 mm處測定出吸光度,重復3次。
EdU增殖檢測:按照CCK-8增殖檢測步驟進行轉染,轉染48 h后進行EdU增殖檢測;將培養基換成100 μl 50 μM 濃度的EdU(銳博生物,廣州)GM培養基,37 ℃培養箱中孵育2 h,其他操作步驟按照說明書進行。倒置螢光顯微鏡觀察并采集螢光圖片,每孔至少采集5組圖片。
1.8 數據分析
采用2-ΔΔct分析定量數據,使用imageJ 1.50e軟件對細胞進行計數和計算百分比,使用SPSS 22對所有數據進行單因素方差分析,統計結果用平均數±標準差(S.E.M)表示,P<0.05表示組間差異顯著,P<0.01表述組間差異極顯著。
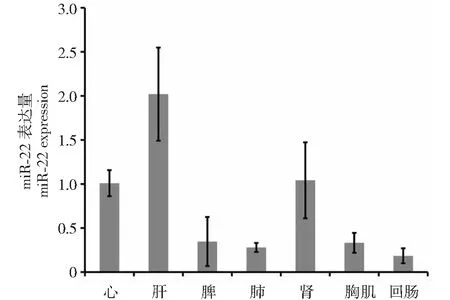
圖1 miR-222在新西蘭兔各組中表達量Fig.1 The expression of miR-222 in New Zealand rabbit different organizations
2 結果與分析
2.1 miR-222在各組織中的表達譜
對心、肝、脾、肺、腎、腿肌和回腸中miR-222進行了定量檢測,發現miR-222在肝臟中的表達量最高,回腸中表達量最低,在腿肌中中度表達。結果如圖1。
2.2 靶基因功能富集分析
使用TargetScan、miRBase和miRWalk 3個在線預測軟件進行靶基因預測,由于這3個數據庫上家兔的基因組不全,選擇在分類學上相近的小鼠基因組作為miRNA—mRNAs預測,篩選出了908個靶基因(3個預測結果取交集),使用DAVID 6.8對靶基因進行GO聚類分析和KEGG通路分析。這些靶基因參與轉錄、轉錄調控、細胞內信號轉導、神經系統發育和調節細胞凋亡和遷移等生物學過程,參與細胞膜、細胞質、神經元投射,細胞核等細胞構成,參與蛋白質結合、金屬離子結合、DNA結合、核苷酸結合,轉錄激活、轉錄因子激活和激酶激活等分子功能(圖2)。此外,miR-222的靶基因富集在以下幾個通路,包括癌癥通路、PI3K-Akt信號通路、MAPK信號通路、人類嗜T細胞病毒Ⅰ型感染、Ras信號通路、催產素信號通路、趨化因子信號通路和CAMP信號通路(表2),這些數據表明,miR-222在生物體內可能調控多種生物學功能。

圖2 GO分析預測miR-222的生物學功能Fig.2 Analysis of miR-222 biological function by GO terms

通路名稱相關基因P-ValuePathways in cancerE2F3, ADCY1, GNAI3, GNAI2, ADCY7, XIAP, LPAR2, GNG12, KIT, CXCL12, FOS, KRAS, CASP9, RASGRP1, GNG2, RUNX1, AXIN2, COL4A3, RET, BRAF, TGFBR1, RUNX1T1, SKP2, RAF1, IGF1, MAPK10, FZD5, STAT1, KITL, MAPK1, CBLB, 4930544G11RIK, MAPK90.0000PI3K-Akt signaling pathwayOSMR, LPAR2, GNG12, KIT, ITGB3, CDC37, KRAS, CASP9, GNG2, PRKAA2, MLST8, GHR, COL4A3, PKN2, YWHAB, IGF1, RAF1, KITL, IRS1, BCL2L11, MAPK1, YWHAG, CC-ND2, CHRM1, TSC2, RELN, PPP2R3C, EIF4E20.0005MAPK signaling pathwayPTPN7, CACNA2D1, NTF3, BRAF, TAOK1, TGFBR1, MAP2K4, PPP3R1, RAF1, MAPK10, CACNB4, GNG12, MAP3K7, FOS, MAPK1, KRAS, PAK2, MAP3K2, RASGRP1, MAPK9, ST-MN1, MAP3K13, NFATC30.0003HTLV-I infectionCRTC3, ADCY1, E2F3, XIAP, ADCY7, CRTC1, TGFBR1, MAP2K4, PPP3R1, NFYB, FZD5, CANX, ATF1, FOS, POLE4, KRAS, CCND2, ETS1, XBP1, ETS2, TRP53INP1, NFATC30.0024Ras signaling pathwayIGF1, RAF1, KIT, GNG12, MAPK10, KITL, RGL1, MAPK1, KRAS, PAK2, GAB2, TIAM1, ETS1, RASGRP1, ETS2, 4930544G11RIK, MAPK9, GNG2, RASA30.0032Oxytocin signaling pathwayADCY1, CACNA2D1, GNAI3, ADCY7, GNAI2, PPP3R1, RAF1, CACNB4, FOS, MAPK1, KRAS, CAMK4, 4930544G11RIK, GUCY1A3, CAMK2B, PRKAA2, NFATC3, CAMK1D0.0097Chemokine signaling pathwayADCY1, GNAI3, BRAF, ADCY7, GNAI2, PREX1, RAF1, GNG12, STAT1, CXCL12, CCL6, MAPK1, KRAS, TIAM1, 4930544G11RIK, CX3CR1, GNG2, WASL0.027cAMP signaling pathwayADCY1, GNAI3, BRAF, ADCY7, GNAI2, RAF1, ATP1A1, GRIA4, MAPK10, FOS, MAPK1, CAMK4, TIAM1, CHRM1, 4930544G11RIK, MAPK9, CAMK2B0.041
2.3 過表達和抑制表達效率
用miR-222、miR-222 mimics、miR-222 inhibitor和inhibitor NC分別轉染SMSCs,48 h后,監測miR-222的表達量。過表達miR-222后,miR-222表達量升高了610倍,mimics組極顯著高于mimics NC組(圖3,P<0.01);抑制表達miR-222后,表達量降低到約55 %,inhibitor組顯著低于inhibitor NC組(圖3,P<0.05)。結果顯示,過表達miR-222和抑制表達miR-222實驗成功。
2.4 miR-222促進SMSCs增殖
用CCK8試劑盒檢測SMSCs在24、48、72、96和120 h時的增殖情況。發現過表達miR-222后,在72、96和120 h時,mimic組的增殖速度顯著高于mimic NC組(圖4A,P≤0.05)。同時,miR-222 inhibitor組在72、96和120 h的增殖速度顯著低于NC組,如圖4B。結果表明,過表達miR-222可以促進家兔骨骼肌衛星細胞增殖,抑制表達miR-222可以抑制家兔骨骼肌衛星細胞增殖。

*P≤0.05,**P<0.01,下同*P≤0.05,**P<0.01, the same as below圖3 轉染效率檢測Fig.3 Detection of transfection efficiency
EdU試驗結果表明miR-222能促進SMSCs增殖,與miR-222 NC組相比,miR-222 mimics組EdU陽性細胞數量多36.4 %(P=0.008);與inhibitor NC組相比,inhibitor組EdU陽性細胞數量少了19.4 %(P=0.037)。
3 討 論
本試驗先分析了miR-222在新西蘭兔各個組織中的表達,發現miR-222在肝臟中表達量最高,在腿肌中中度表達。利用生物信息學預測了miR-222的靶基因,并對靶基因進行GO分析和KEGG pathway分析來推測miR-222的生物學功能,發現靶基因主要富集在轉錄因子激活、激酶激活、核苷酸結合等分子功能,細胞遷移、發育和凋亡等生物學過程,癌癥通路、PI3K-Akt信號通路、MAPK、Ras、cAMP等信號通路,這些通路對肌肉的發育起著重要的調控作用,例如:PI3K-Akt通路可以抑制肌肉特異性的泛素連接酶MuRF1和MAFbx的表達從而抑制肌肉萎縮[15],此外,比如胰島素樣生長因子1(insulin-like growth factor I,IGF-1)必須通過PI3K-Akt的磷酸化才能促進肌衛星細胞增殖[16];而MAPK信號通路可以通過Myf5誘導胚胎早期祖細胞的成肌分化,激活MyoD和MEF2家族中的成肌轉錄因子促進成肌細胞增殖和分化[17]。而且,miR-222在人類肌營養不良疾病以及MDX肌營養不良鼠模型的肌肉組織中的表達量都會升高[18-19],這些都暗示著miR-222可能對骨骼肌衛星細胞的增殖有調控作用。
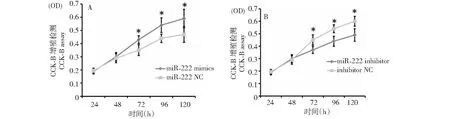
圖4 CCK-8實驗檢測miR-222對SMSCs增殖的影響Fig.4 The effect of miR-222 on the proliferation of SMSCs cells via CCK-8 assay
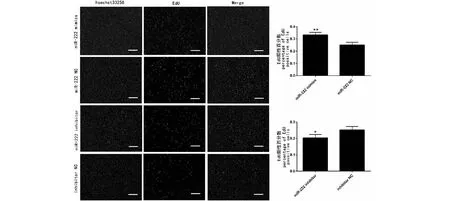
細胞核被hoechst標記成藍色,增殖的細胞被EdU標記成紅色,Merge為兩種染色疊加,EdU陽性細胞百分數計算公式為:(EdU陽性細胞數/細胞核數量)×100 %,比例尺為200 μmCell nuclei (blue) and EdU positive cells(red). The ratio of EdU-positive cells was calculated with (EdU-positive cells/Hoechst stained cells)×100 %. The scale bars stand 200 μm圖5 EdU試驗檢測miR-222對SMSCs增殖的影響Fig.5 The effect of miR-222 on the proliferation of SMSCs cells via EdU assay
使用miR-222 mimics、miR-222 inhibotor分別對SMSCs進行miR-222過表達和抑制內源性miR-222表達,CCK-8增殖檢測和EdU增殖實驗結果均表明,miR-222對SMSCs的增殖有促進作用。miR-222在骨骼肌中的研究較少,Cardinali等[14]研究發現,miR-222可以促進C2C12小鼠成肌細胞的增殖,與本研究結果一致。在外周動脈缺血后,miR-222能夠通過超氧化物歧化酶-2(superoxide dismutase‐2,SOD-2)抑制P57的轉錄后表達,從而促進四肢骨骼肌的再生[20]。在心肌細胞中,miR-222也能促進心肌細胞的增殖,在小鼠心臟內對miR-222的表達進行抑制,發現增殖的標記基因(HIPK1和HMBOX1)表達量降低,心肌細胞和心臟的增殖發育受到阻礙[21]。在平滑肌中,miR-222靶向P27促進哮喘病人氣管平滑肌的增殖。P27和P57是細胞周期抑制因子,通過抑制細胞周期蛋白依賴性激酶(cyclin-dependent kinase,CDKs)的表達從而抑制G1-S、G2-M之間的轉換,引起細胞增殖受到抑制[10],在骨骼肌中,P57和P27也可以同樣地抑制細胞的增殖,Chakkalakal等[22]研究發現,P27抑制小鼠骨骼肌的增重,敲出P27的骨骼肌衛星細胞可以促進細胞的增殖以及細胞損傷后的自我更新。Naito等[23]研究發現用RNAi技術抑制P57的表達,導致PAX7的表達量增加,從而促進骨骼肌衛星細胞的增殖。
4 結 論
本研究結果表明,miR-222在新西蘭兔各組織中差異表達,在肝臟中表達量最高,回腸中表達量最低,在肌肉中中度表達;用miR-222 mimics和inhibitor模擬miR-222在SMSCs中過表達和抑制表達,結果表明,miR-222對骨骼肌衛星細胞增殖有促進作用。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
