白酒窖泥微生物多樣性研究方法及進展
2018-11-09 05:37:46郭壯趙慧君雷敏沈馨蔡宏宇張振東
食品研究與開發 2018年22期
郭壯,趙慧君,雷敏,沈馨,蔡宏宇,張振東
(湖北文理學院化學工程與食品科學學院鄂西北傳統發酵食品研究所,湖北襄陽441053)
白酒是中國特有的一種蒸餾酒,由于白酒的釀造工藝、生產環境以及酒曲制作方式的不同,形成了濃香型、醬香型、清香型、米香型等主要香型白酒,還由此衍生出兼香型等多種類型[1]。濃香型白酒以糧谷為主要原料,經傳統固態法發酵,蒸餾,陳釀,勾兌而成,以己酸乙酯為主體復合香,是我國銷量最大的白酒類型[2]。濃香型白酒使用泥窖發酵,窖池的優劣對濃香型白酒生產至關重要,以優質老窖養老糟才能生產好酒。上千年的馴化與自然選擇,孕育了知名酒廠酒窖中獨特的微生物區系,它們相互共生,相互競爭,通過復雜的相互作用關系,形成了一個穩定的生態群體。窖泥是白酒固體發酵過程中微生物的重要來源,是影響白酒的口感與品質的重要因素[3-4]。因此,對窖泥中微生物的研究意義重要性不言而喻。
近幾十年來,科技不斷進步,窖泥中的微生物研究技術手段也在不斷推陳出新。首先是不依賴于聚合酶鏈式反應(polymerase chain reaction,PCR)的傳統的培養方法、磷脂脂肪酸(phospholipid fatty acid,PLFA)以及熒光原位雜交(fluorescence in situ hybridization,FISH)等方法。然后是基于PCR的宏基因組研究策略如克隆文庫,變形梯度凝膠電泳(denaturing gradient gel electrophoresis,DGGE)以及目前研究的主流-第二代測序技術,也是目前窖泥微生物研究的主要方法。本文將對濃香型白酒窖泥微生物研究中常用的技術方法進行介紹,并介紹它們在白酒窖泥微生物研究中的應用進展。
1 傳統純培養方法及其在窖泥微生物研究中的應用
純培養技術是最早應用于微生物群落結構的研究手段,主要是使用不同營養成分的培養基對環境樣品中微生物進行分離,然后根據微生物的菌落形態、生理生化來確定微生物的分類情況。后來引入了分子生物學方法后,對所分離的純培養原核微生物的16S rDNA[5-6]基因和真菌微生物的26S rDNA或者轉錄間隔區(internal transcribed spacer,ITS)[7]間區序列對微生物進行分類,以此獲取微生物的群落結構信息。
以純培養方法對窖泥中的微生物研究由來已久。但是由于自然環境中僅有0.1%~10%微生物可以被培養分離,采用傳統的純培養方法容易導致目標環境中大量微生物無法培養而造成對樣品中微生物多樣性的低估。以岳元媛等的研究為例:采用厭氧培養方法對窖泥中的細菌進行了篩選分離,共得到8個屬的細菌,分別是芽孢桿菌屬、假單胞菌屬、梭菌屬等,絕大多數為兼性厭氧細菌,梭菌屬細菌僅占1%左右[7],這與采用未培養技術得到的結果大相徑庭。
盡管傳統純培養技術具有許多局限性,但是該技術到現在仍在被廣泛使用。這主要是由于該技術是研究特定種類的微生物生理生化特征,并將一些微生物應用于工業化的重要方法。科研人員也對傳統純培養技術進行了改進[8-9]:模擬自然環境,在分離海洋細菌時,使用海水配制培養基;改善培養條件,在培養厭氧細菌時,使用厭氧工作站;使用共培養技術,將共棲的兩種或者數種微生物一起培養等等,但是仍然達不到未培養方法對樣品中微生物多樣性研究的效果,因此發展起來了多種不依賴于傳統培養技術的方法。
目前采用傳統純培養技術對窖泥中的微生物研究,多集中在梭菌屬。最初對白酒窖泥中的微生物研究發現,在這些復雜多樣的微生物中,有一種呈鼓槌狀的產芽孢桿菌(梭菌),能以乙醇、乙酸經丁酸合成己酸,這類微生物被稱為己酸梭菌。1942年,該菌被命名為Clostridium kluyveri[10]。自從1964年己酸乙酯作為白酒中的香氣成分被發現以來,己酸菌在白酒發酵過程中的作用逐漸被人們認識到,優質的己酸菌被發掘出來,如內蒙30、黑輕80等等,黑輕80己酸產量為約380 mg/100 mL[11]。學者對典型的濃香型白酒如瀘州老窖[12],五糧液酒業[13],安徽金種子酒業[14]江蘇湯溝酒業[15]等窖池窖泥中己酸菌進行了大量的研究工作,收集到了不少品質優良,己酸產量高的菌株,最高己酸產量可達4.36 g/L。己酸細菌是窖泥微生物的研究中最重要的發現之一,其含量是評估窖泥質量的基礎。優質的己酸菌被用于退化窖池的養護[16-17],還被用于生產人工窖泥[18]。使用己酸菌制作的人工窖泥在短期內就可生產出優質酒,打破了非50年老窖不能生產名酒的說法。
2 PLFA技術及其在窖泥微生物研究中的應用
瀘州老窖是國內濃香型白酒的典型代表,保存有入選世界吉尼斯紀錄,擁有400多年窖齡,國內最古老的白酒窖池。對窖泥微生物多樣性報道最多的當屬對瀘州老窖酒業的窖泥研究,研究方法集中在了PLFA,克隆文庫,聚合酶鏈式反應-變性梯度凝膠電泳(poly merase chain reaction-denaturing gradient gel electrophoresis,PCR-DGGE)及第二代測序技術。
磷脂脂肪酸是生物細胞膜的主要成分,僅存在于活細胞膜中,當微生物死亡后,脂肪酸就會被代謝掉。細胞中包含脂肪酸的脂類物質主要有碳水化合物、脂性醇、磷脂、糖脂和中性脂等[19]。通常這些脂類物質在同一種微生物中是穩定的,并且操作難度與試驗條件要求較低,因此PLFA指紋圖譜技術被開發出來,在對窖泥、土壤、食品等多種環境的微生物的研究中得到應用[19-21]。Tunlid在1985年首次利用磷脂脂肪酸技術對油菜根際微生物的群落結構進行了研究,此后該技術逐漸被引入到其他的微生物研究領域。窖泥是一種特殊的土壤,因此在窖泥微生物的研究中也引入了磷脂脂肪酸技術。劉琨毅等[21]對窖泥微生物的分析表明,厭氧革蘭氏陽性菌和真菌是窖泥中的優勢菌群,不同窖齡窖泥中的微生物PLFA含量不同,300年窖齡中PLFA含量大于5年與100年窖齡[22]。但是另一研究報道了瀘州老窖 20、50、100、200、300 年窖泥中革蘭氏陰性菌、革蘭氏陽性菌與需氧菌的比例并無顯著差異[23]。
總體上來說,PLFA技術不依賴于分離和培養的技術,更為快速、簡便,操作難度更低,但是磷脂脂肪酸分析方法不能對微生物在種或菌株水平上加以區分[25],所以常與其他分析技術,如DGGE或傳統培養方法相結合,用于的窖泥微生物研究。
3 克隆文庫技術及其在窖泥微生物研究中的應用
克隆文庫是20世紀末在對微生物多樣性的研究中被開發出來。通過構建克降文庫分析微生物群落時,首先要獲得樣品總DNA,以提取到的總DNA為模板PCR擴增微生物的16S rDNA基因,然后將得到的擴增產物與載體連接,轉化感受態細胞,通過藍白斑篩選,挑取陽性克隆進行測序,根據測序結果判斷樣品中的微生物信息[25-27]。這一方法最早在1991年被Giovannoni等用來分析浮游細菌多樣性[28]。由于克隆文庫法針對原核微生物的16S rDNA全長,且避開了傳統的純培養方法,能將一些對營養要求苛刻的微生物鑒定出來,因此能更全面的反映樣品中的微生物多樣性;由于采用該方法能幾乎獲取微生物的16S rDNA全序列,所以結果也更加準確,被用于白酒窖泥微生物的研究。
2013年,劉森等[25]采集了四川一濃香型白酒公司20年窖池窖泥樣品,從窖池上層窖泥中檢測到Clostridium、Lactobacillus兩個菌屬,中層檢測到5個菌屬,分別是 Lactobacillus、Serratia、Clostridium、Bacillus、Caloramator,窖池下層檢測到4個菌屬,分別是Lactobacillus、Clostridium、Bacillus、Caloramator,發現窖池不同位置的微生物分布顯著不同。在所有樣品中,Clostridium與Lactobacillus菌屬在各層樣品中均占20%以上,為優勢菌屬。2014年[28],同樣采用克隆文庫法對安徽金種子酒業的濃香型白酒窖泥微生物進行了解析,發現退化窖泥中優勢菌門為Firmicutes與Bacteroidetes,而老窖泥中Firmicutes與Chloroflexi為優勢菌門。盡管通過克隆文庫法對窖泥的研究取得了一些成果,但是由于該法操作繁瑣,且耗時耗力,因此已逐漸被DGGE與二代測序技術取代很少出現在近幾年的研究報道中。
4 PCR-DGGE技術及其在窖泥微生物研究中的應用
DGGE技術能用于環境中細菌、藍細菌、古菌、微微型真核生物、真生物和病毒群落的生物多樣性的分析,目前主要研究對象是原核微生物及真菌微生物。原理是在聚丙烯酰胺凝膠基礎上,加入了變性劑(尿素和甲酰胺)使聚丙烯酰胺凝膠從上到下呈現從小到大的變性梯度,PCR產物沿著化學梯度有不同解鏈行為,在凝膠的不同位置上停止遷移而分離開來[29]。PCR-DGGE分析微生物多樣性的試驗流程為:首先是樣品總DNA的提取;然后以總DNA為模板進行PCR擴增;制作相應梯度的聚丙烯酰胺凝膠,然后在樣品孔添加樣品的PCR產物進行聚丙烯酰胺凝膠電泳;對聚丙烯酰胺膠進行染色,挑取條帶回收,克隆與測序,對聚丙烯酰胺膠進行分析[29-30]。
細菌16S rRNA基因含有多個可變區與保守區[5](見圖1)。
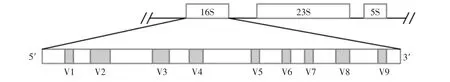
圖1 細菌16S rRNA基因結構[5]Fig.1 The schema of ribosome complex and 16S rRNA gene
由于DGGE對大于500 bp的DNA片段分離度較差,通常使用16S rRNA基因中的一個或者兩個區段。由于PCR-DGGE技術無需培養,檢測快速,成本低,目前仍然與第二代測序技術成為當前白酒窖泥中微生物研究的主要技術。自從1993年,Muyzer[30]第一次將PCR-DGGE技術引入到微生物多樣性的研究以后,該技術就在與微生物相關的各個領域得到廣泛應用,在白酒窖泥微生物的研究也引入了PCR-DGGE技術(見表1),且成為白酒窖泥研究的主要研究方法。
2014年,Ding等[34]使用DGGE法對瀘州老窖不同窖齡的人工窖泥中微生物進行了研究:共檢測到2個細菌菌門,分別是Proteobacteria和Firmicutes,檢測到9個菌科。樣品中的主要微生物類群是Clostridiaceae,Lactobacillaceae。Clostridiaceae占到總微生物的46.2%;而Lachnospiraceae微生物僅在新窖泥中出現,而在3年與4年窖齡窖泥中消失,而Ruminococcaceae相反,在4年窖齡窖泥中出現,而在新窖泥中消失。2013年,Zheng等[23]對瀘州老窖20年窖齡與300年窖齡窖泥的分析表明,20年窖泥主要類群為Lysinibacillus與Clostridium;300年窖泥主要類群為Lysinibacillus,Soilbacillus,Clostridium,不同窖齡窖泥中微生物存在顯著差異。2015年,胡曉龍等[35]還專門針對窖泥中己酸梭菌16S rRNA基因序列的V4和V5可變區,設計了DGGE檢測引物對(SJ-F和SJ-R),能特異性解析窖泥中梭菌菌群多樣性,能靈敏且準確的對窖泥梭菌菌群組成及多樣性進行研究。
雖然目前PCR-DGGE具有無需培養,且檢測快速,成本低等一系列優勢,是白酒窖泥微生物群落結構的主要研究方法,但仍具有一些缺陷[48]:僅能檢出樣品中含量超過1%的微生物,對一些含量低但可能具有重要功能微生物不能有效地檢測;DGGE圖譜中條帶信息量低,可能會低估窖泥中優勢微生物的多樣性;PCR產物片段長度大于500 bp的序列不適用于該法,因此在以后可能會被高通量測序技術取代。
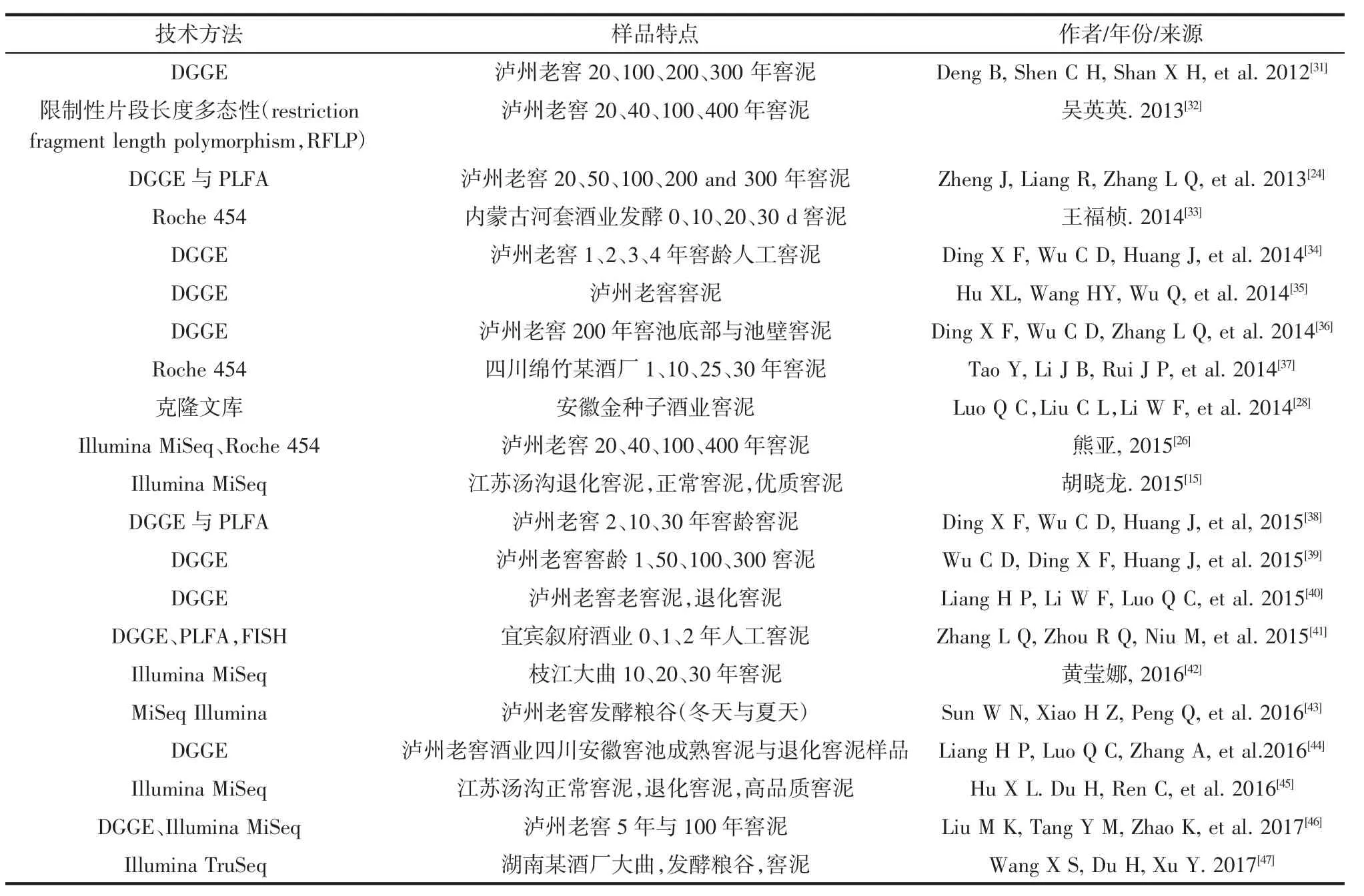
表1 白酒窖泥中微生物的研究方法Table 1 Research technique for study in microbiology in pit mud
5 第二代測序技術及其在窖泥微生物研究中的應用
1977年Sanger發明了雙末端終止測序法,即第一代測序方法。測序技術不斷革新,科研人員在一代測序技術的基礎上,開發出的二代測序技術,即高通量測序法逐漸成為目前窖泥中微生物研究的重要手段,在傳統發酵食品微生物中得到廣泛的應用[49]。高通量測序技術的應用從2006年至今呈現明顯的上升趨勢。目前常用的二代測序平臺主要是以Illumina公司的Solexa平臺、ABI公司的SOLiD平臺和Roche公司的454平臺為代表[50-51]。二代測序測序的基本操作流程主要包括克隆文庫制備、DNA片段固定、DNA片段單分子擴增、測序反應、光學圖像采集與處理及序列拼接及組裝[50],具有:通量高(能得到0.035 Gb~1 800 Gb的數據信息);準確率高;成本低;試驗周期短;相比于傳統可培養及其它分子手段,能檢測到環境樣品中稀有種屬,即能全面客觀地揭示目標環境中微生物群落信息,同時獲得定性及相對定量信息等優勢[50-51],因此被廣泛的應用于窖泥及其他樣品中微生物的多樣性研究。
2017年,Liu等[46],同時采用DGGE與Illumina Mi-Seq二代測序技術分別對瀘州老窖5年與100年窖泥中的真菌微生物進行了研究。DGGE法僅檢測到了樣品中12個菌屬,而MiSeq法檢測到檢測到4個菌門(Ascomycota,Zygomycota,Basidiomycota,Chytridiomy -cota)與111個菌屬,所獲取的窖泥中的微生物信息量遠大于DGGE法:瀘州老窖5年與100年樣品中檢測到 Rhizopus,Aspergillus,Phoma等 19 個核心菌屬,5 年窖齡窖泥優勢菌屬為 Rhizopus,Phoma,Trichosporoni;100年窖齡優勢菌屬為 Aspergillus,Candida。按照Yilmaz等的觀點,自然界中絕大多數或者99%的微生物在實驗室難以富集培養生長[52],而DGGE由于技術本身的缺陷不能對低于1%的微生物類群進行表征,因此高通量測序法具有天然的優勢。
酒窖窖泥環境比較特別,含有較高的乙醇量,較低的pH值及較低的氧氣含量,窖泥中微生物群落結構復雜多樣,對瀘州老窖30年與300年窖齡的窖泥中細菌微生物進行了研究[53]:300年窖泥中古菌主要屬于Euryarchaeota,占 8.8%~16.6%;30年窖泥中 Euryarchaeota僅占0.6%,含量遠低于300年窖泥。在30年窖泥中,Firmicutes是優勢菌門,占到92%以上,主要是乳酸菌Lactobacillus,占到90%以上;而300年窖泥中,Firmicutes也作為優勢菌門,占到43%以上,主要是乳酸菌Lactobacillus,占到22%~41%,遠遠低于30年窖泥。而Clostridiales在300年窖泥中占到21%~40%,Clostridium 占到 2.6%~4.9%;Clostridiales在300年窖泥中占到約2.5%,Clostridium僅占到0.35%。可以看出,新老窖池窖泥中微生物群落結構存在顯著差異。Hu等[45]對江蘇湯溝濃香型白酒優質窖泥、普通窖泥與退化窖泥中細菌微生物進行了分析,發現在退化窖泥有中僅有 2個優勢菌屬:Lactobacillus,Ruminococcus;常規窖泥中有 Lactobacillus,Caloramator,Clostridium等12個優勢菌屬;而優質窖泥中有Lactobacillus,Caloramator,Clostridium等 15個優勢菌屬。隨著窖泥質量的增加,檢測到Lactobacillus含量顯著減少,而 Clostridia、Bacteroidia、Methanobacteria及 Methanomicrobia 4個菌綱的核心屬含量明顯增加,表明這4個綱的微生物含量的增加及Lactobacillus含量減少有利于提高窖泥的質量。分析后認為,高乳酸含量、低NH4+、pH值、有機磷含量是導致窖泥退化的外因。
另外胡曉龍等[15]通過網絡分析方法分析了微生物群落結構的相關性,從正相關網絡圖譜中獲得了13個hubs,這些hubs與其他微生物間有著很強的相關聯系,在這些微生物之間也存在協同作用,有利于窖泥環境中的碳、氮及硫元素循環及窖泥風味物質形成。從負相關網絡圖譜中共發現3個hubs包括Lactobacillus、Pediococcus和Streptococcus 3個屬是典型的乳酸菌菌屬,它們與多種微生物呈負相關關系。這也說明乳酸菌與其它窖泥微生物類群存在著一定的拮抗作用,能抑制其它微生物類群的生長繁殖。窖泥微生物類群之間存在著復雜相互作用關系,認識和解析窖泥中的微生物信息,將有助于人們采取措施有目的干預窖泥中的特定微生物,來提升白酒酒質與白酒生產效率。
雖然通過第二代測序方法比DGGE法獲取的信息量更多,對窖泥中微生物的解析更為精細,但事實上二代測序技術并非無懈可擊。由于測序讀長所限大部分研究只能以16S rRNA可變區部分區段為擴增、測序靶點,致使對微生物分析鑒定只能停留在“屬”水平,甚至“科”水平,不能對環境中微生物群落結構進行精準描述。
6 第三代測序技術簡介
科研人員針對二代測序技術的不足,開發出第三代測序技術,包括Helicos Biosciences公司的單分子DNA 測序(true single molecular sequencing,tSMS)、Pacific Bioscience(PacBio)公司的單分子實時測序(single molecule real time sequencing,SMRT) 以 及 Oxford Nanopore的納米孔單分子技術[54-55]。以PacBio SMRT測序技術為例,PacBio公司的SMRT技術基于邊合成邊測序的思想,核心是零模式波導技術(zero-mode waveguide technology,ZMW),以SMRT芯片為載體進行測序反應。ZMWs直徑為100 nm,厚度為70 nm,剛好容納一個DNA聚合酶分子,從而觀察到DNA鏈合成過程。測序過程中,DNA聚合酶附著在ZMW孔底部,攜帶熒光標記的堿基,以單分子DNA為模板。在讀取模板過程中,DNA聚合酶能結合不同堿基會發出不同顏色的信號,從而判別堿基種類[56-57]。
Mosher等[56]在2014年,使用PacBio SMRT測序技術對環境樣品微生物多樣性進行研究,認為SMRT技術比Roche/454測序技術精度要高。對16S rDNA基因V4區的測序表明,SMRT技術的錯誤率為Roche 454與MiSeq等二代測序平臺的1/8[57]。使用SMRT技術對鐵皮石斛和丹參基因組的研究,充分說明了SMRT技術的優越性,讀長更長,且能免受DNA中高GC含量的影響,在對復雜基因組完整組裝分析時,體現出了巨大的優越性[58]。此外,SMRT技術能識別DNA的甲基化,應用到DNA甲基化研究中。在對傳統發酵食品如米酒曲[59],泡菜[60],乳制品與腸道微生物[61]的研究中也引入了SMRT三代測序技術,但尚未見到三代測序技術在白酒窖泥微生物研究中的任何報道。
7 宏基因組技術及其在窖泥微生物研究中的應用
宏基因組學也稱為環境基因組學,元基因組學,生態基因組學。Handelsman等[62]于1998年第一次將宏基因組(metagenome)定義為‘the genomes of the total microbiota found in nature’,即環境中全部微生物群的所有遺傳物質,以環境樣品中的微生物群體基因組為研究對象,以微生物多樣性,種群結構,進化關系,功能活性,相互協作關系及與環境之間關系為研究目的的研究方法。與傳統培養法及高通量測序技術相比,宏基因組測序分析有以下優點:無需培養微生物,能客觀全面的還原菌群結構;測序周期短;測序通量大、靈敏度高,能對樣品中總的微生物群落結構進行功能及代謝通路的分析[63-64]。
宏基因組技術的分析步驟為:首先提取環境樣品DNA,經克隆轉化、構建文庫,最后上機測序。通過生物信息學分析,將下機后的原數據去除污染和接頭序列、去除含N堿基序列、去除質量值小于20的序列、去除長度小于50 bp的短序列,獲得的高質量序列進行序列拼接。獲得的數據均進行兩方面的分析:一方面進行物種分類學注釋,分析種群分布,對不同生境下的種群進行比較分析;另一方面進行功能分析,首先將拼接好的Contigs進行開放閱讀框預測,之后利用數據庫進行功能注釋和代謝通路分析。常用的功能注釋數據庫有 COG、KEGG、Nt、Nr、GO、Swiss-Prot、SEED等。常用的代謝通路數據庫包括KEGG、RegulonDB、BioCyc、WikiPathwans、Reactome[64-66]等。
白酒固態發酵是一個復雜的過程,在窖泥中存在著大量難以培養的微生物,因此,通常對于菌群發酵機制及其相互作用知之甚少,而元基因組的方法不需要對菌株進行分離而直接測序,對窖泥微生物研究非常有效。2017年,Tao等[67]通過metagenomics法對綿竹市的濃香型白酒窖泥進行了研究,檢測到了脂肪酸鏈延伸途徑中的關鍵基因,重構了脂肪酸碳鏈延伸途徑,該途徑含有己酸合成途徑的關鍵酶,表明窖泥微生物具有以乙醇或丙酮酸為底物合成己酸的能力。另外獲取了窖泥微生物中甲烷合成途徑中編碼產氫酶與乙酸分解酶的基因,表明在甲烷菌與己酸菌之間存在氫轉移作用,重構了基于Clostridium、ClostridialclusterIV、Methanoculleus與 Methanosarcins種間氫轉移作用的己酸合成途徑。另外根據分析結果,Clostridialcluster IV,Caloramator,Clostridium,Sedimentibacter,Bacteroides與 Porphyromonas 6個菌屬構成了窖泥中己酸合成的活性微生物環境,Clostridialcluster IV與Clostridium能直接合成己酸。
8 展望
濃香型白酒采用泥窖發酵,在白酒糟醅發酵過程中,窖泥中的微生物逐漸進入到白酒糟醅中,窖泥來源的微生物約占到糟醅中總微生物的14%[47],多數屬于厭氧微生物。由于形成濃香型白酒主體香型物質己酸乙酯前體己酸的微生物多數屬于厭氧梭菌[15],窖泥微生物在白酒發生產中的重要性不言而喻。克隆文庫、DGGE、第二代測序技術等分子生物學技術的發展,使人們對窖泥微生物多樣性的認識上了一個新臺階,但并不能滿足人們對白酒窖泥中微生物研究的要求。白酒窖泥環境,具有低pH值,高乙醇含量,低氧等特殊的特點,經過長時間的富集與馴化,蘊含了復雜的微生物類群。為進一步認識白酒窖泥間微生物在代謝上的聯系,濃香型白酒以己酸乙酯為主體的復合香產生的微生物機制,對白酒香型物質產生具有重要貢獻的微生物之間的生態關系,對白酒香氣化學組分形成不利的微生物類群,新的科學技術的出現為白酒窖泥的這些科學問題的解決提供了可能。
鑒于第三代高通量測序技術的讀長可以覆蓋16S rRNA全長,測序精度高,不受高GC影響等一系列優點而越來越受到學者的青睞,同時宏基因組策略也在微生物群落代謝與功能中的研究中得到廣泛應用。預計將來第三代測序技術與宏基因組在白酒窖泥中的應用會越來越多,將兩者結合起來,將有助于人們加深對白酒固態發酵過程的認識,破解窖泥微生物群落中與白酒香型形成的基因代碼與代謝機制,解決人們在白酒生產中的問題,以便更高效率的生產高品質的白酒。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
兒童故事畫報(2019年5期)2019-05-26 14:26:14
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
Coco薇(2016年2期)2016-03-22 02:42:52
Coco薇(2015年1期)2015-08-13 02:47:34
小雪花·成長指南(2015年7期)2015-08-11 15:03:12
