羊瘙癢因子263K感染倉鼠腦組織中蘭尼堿受體2表達變化的研究
2018-11-14 11:34:28李堯黎建樂肖康馬月高麗萍王晶甄禎郭燕軍石琦
中華實驗和臨床病毒學雜志 2018年5期
關鍵詞:研究
李堯 黎建樂 肖康 馬月 高麗萍 王晶 甄禎 郭燕軍 石琦
100050北京,首都醫科大學附屬北京友誼醫院神經內科(李堯、黎建樂、甄禎、郭燕軍);102206北京,中國疾病預防控制中心病毒病預防控制所朊病毒室(肖康、馬月、高麗萍、王晶、石琦)
朊病毒病是一種可侵襲人及動物神經系統的退行性疾病,致死率100%,目前尚無有效治療措施。根據目前流行的朊病毒病發病機制學說,神經系統中正常存在的朊蛋白(prion cellular protein,PrPc)錯誤折疊后形成具蛋白酶抗性的異常朊蛋白(scrapie isoform of prion protein,PrPSc),在神經系統沉積并導致神經元退行性變[1]。人類的朊病毒病包括人克雅氏病、庫魯病、致死性家族失眠癥等。動物朊病毒病包括羊瘙癢病(scrapie)、牛的海綿狀腦病(bovine pongiform encephalopathy,BSE)及麋鹿的慢性消耗性疾病(chronic wasting disease,CWD)等。朊病毒病典型的中樞神經系統病理改變包括淀粉樣斑塊沉積、神經元細胞丟失、膠質細胞增生和海綿樣空泡變性。
鈣離子是一種重要的細胞內第二信使。鈣離子穩態對于維持細胞的正常形態和功能具有重要意義[2]。細胞胞漿內的鈣離子主要來源于兩個方面,分別為胞外鈣離子內流和胞內內質網或肌漿網內的鈣庫。蘭尼堿受體(ryanodine receptors,RyRs)作為一種廣泛存在于細胞內質網和肌漿網的鈣離子通道,他的開放可導致內質網或肌漿網中的鈣離子進入胞漿內,參與維持細胞內鈣離子穩態的平衡。
RyR2是蘭尼堿受體家族的代表成員,不僅維持細胞內鈣離子穩定,并在神經細胞發育和凋亡中起重大作用。研究發現,RyR2參與到阿爾茲海默病等變性疾病的發病過程,與異常蛋白如Aβ淀粉樣蛋白的沉積關系密切。但對于RyR2與朊病毒病相互關系的研究甚少,RyR2在朊病毒感染人及哺乳動物中的表達變化尚未明確。本課題組在前期研究中應用2D-DIGE差異蛋白質組學研究方法發現,在遺傳性朊病毒病腦組織中,RyR2表達明顯上調,提示RyR2可能與朊病毒感染相關,為探索RyR2與朊病毒病之間的相互關系提供理論線索[3]。
本研究利用羊瘙癢因子倉鼠敏感株263 K感染的倉鼠為動物模型,通過免疫印跡方法探索RyR2在263 K感染倉鼠腦組織中表達水平的變化。通過免疫組織化學染色,明確了RyR2在朊病毒感染病變組織中的表達分布,并初步分析了RyR2與朊蛋白PrP的共定位關系,為進一步探討和揭示RyR2在朊病毒病發病過程中的作用和分子機制提供實驗依據和基礎。
1 材料與方法
1.1 腦組織勻漿制備 羊瘙癢因子263 K感染的黃金倉鼠終末期(感染后第80 d)腦組織樣品保存于-70℃冰箱,使用時分別用裂解液(10 mmol/L NaCI,10 mmol/L EDTA,0.5%NP 40,0.5%Sodium deoxycholate,10 mmol/L,Tris,pH7.4)制成10%(wt/vol)腦勻漿,2 000 g離心10 min去除組織碎片,取上清,置于-70℃備用。
1.2 Western blot檢測 取15μg腦組織勻漿樣本經SDS上樣緩沖液處理后進行8%SDS-PAGE,電轉至NC膜上,檢測RyR2蛋白的電轉移使用槽式濕轉,將轉移槽置于冰浴中,以200 V穩壓電轉3.5 h。檢測β-actin使用半干式電轉移,以160 mA穩流電轉1.5 h。轉膜成功后以5%脫脂奶封閉1 h,一抗孵育,4℃過夜后以PBS浸泡5 min×3次,以二抗室溫孵育2 h,ECL顯色。反應條帶用軟件Image J測定灰度值進行定量分析。所用抗體及稀釋比例如下:RyR2兔源多克隆一抗(Abcam,1∶5 000稀釋)、β-actin 鼠源單克隆一抗(Santa,1∶5 000 稀釋);二抗為HRP標記的抗鼠及抗兔IgG抗體(Santa,1∶5 000稀釋)。
1.3 免疫組織化學染色 腦組織切片經常規脫蠟至水后,復合消化酶處理2 min,PBS浸泡2 min,微波抗原修復30 min,復溫后PBS浸泡2 min,3%過氧化氫處理15 min,PBS浸泡3 min×3次,5%BSA室溫封閉1 h,RyR2一抗孵育4℃過夜,PBS浸泡5 min×3次;加入二抗,37℃孵育0.5 h,PBS洗5 min,3次,DAB顯色,蘇木素復染、透明及封片,顯微鏡下拍攝照片。利用Image Pro Plus 6.0軟件對免疫組織化學圖片測定灰度值,進行定量分析。所用抗體及稀釋比例如下:RyR2兔源一抗(Abcam,1∶200稀釋);二抗為生物素標記羊抗兔IgG試劑盒(中衫金橋)。
1.4 免疫熒光雙標染色 腦組織切片處理同常規免疫組化方法,5%BSA室溫封閉1 h后,加入RyR2特異性兔源多克隆抗體與PrP特異性鼠源單克隆抗體3F4的混合一抗,4℃孵育過夜后,PBS浸泡5 min×5次;加入 AlexaFlour 546標記的羊抗兔與AlexaFlour 488標記的羊抗鼠混合二抗,37℃孵育2 h,PBS浸泡5 min×5次,室溫避光下DAPI染色20 min,PBS浸泡5 min×5次。抗熒光衰減封片劑封片,Operetta CLS高內涵細胞成像分析系統下觀察。所用抗體及稀釋比例如下:RyR2兔源一抗(Abcam,1∶200 稀釋)、3F4 鼠源一抗(Santa,1∶250 稀釋);AlexaFlour 546標記的羊抗兔二抗(Invitrogen,1∶200稀釋)、AlexaFlour 488標記的羊抗鼠二抗(Invitrogen,1∶200 稀釋)。
1.5 統計學方法 實驗數據應用SPSS13.0軟件進行統計分析,兩組間差異比較采用t檢驗,P<0.05為差異有統計學意義。
2 結果
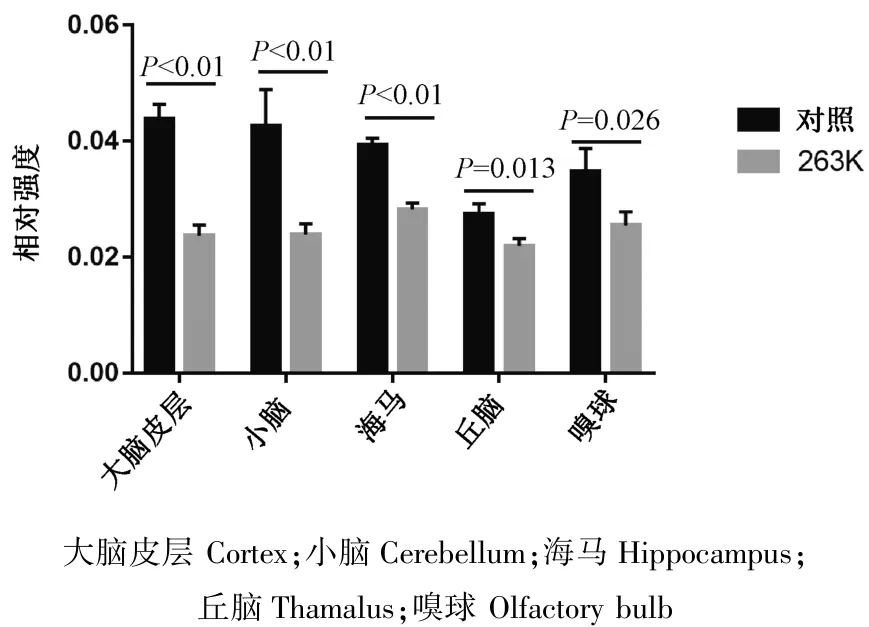
2.1 RyR2在朊病毒感染腦組織各腦區的表達為了探究RyR2在朊病毒感染腦組織中的分布和定位,本研究選取了正常倉鼠及羊瘙癢因子263 K感染倉鼠終末期(感染后第80 d)腦組織切片進行免疫組織化學染色。結果顯示,RyR2在正常及263 K感染倉鼠腦組織中均有分布,兩者分布區域大致相同,主要分布定位于小腦浦肯野細胞層及大腦皮層,在海馬、下丘腦和嗅球也有分布。其中在正常倉鼠的各腦區中,與感染倉鼠相比,可以發現RyR2染色增強、陽性細胞數增多(圖1)。利用Image Pro Plus 6.0軟件對圖片測定灰度值,進行熒光強度定量分析,結果顯示,263 K感染倉鼠各腦區RyR2表達均明顯降低,差異有統計學意義(圖2)。
2.2 朊病毒感染倉鼠模型中RyR2表達降低 為了探究在朊病毒感染動物模型中RyR2表達的變化,本研究選取正常倉鼠及羊瘙癢因子263 K感染終末期倉鼠(感染后第80 d)為研究對象,利用Western blot檢測 RyR2表達含量的變化。以 βactin為內參進行相對定量,使用Image J對目的蛋白條帶進行灰度定量分析,結果利用t檢驗分析,結果顯示,在263 K感染倉鼠腦組織中,RyR2的表達均較正常對照組明顯下降,差異有統計學意義(P=0.037)(圖3)。
2.3 RyR2與朊蛋白存在共定位關系 為了探究RyR2與朊蛋白(PrP)是否存在共定位關系,本研究選取了正常倉鼠及羊瘙癢因子263 K感染倉鼠終末期(感染后第80 d)腦組織切片進行免疫熒光雙標染色。結果提示,在正常及263 K感染倉鼠腦組織切片的大腦皮層中,均發現了RyR2與PrP蛋白存在共定位關系(圖4)。
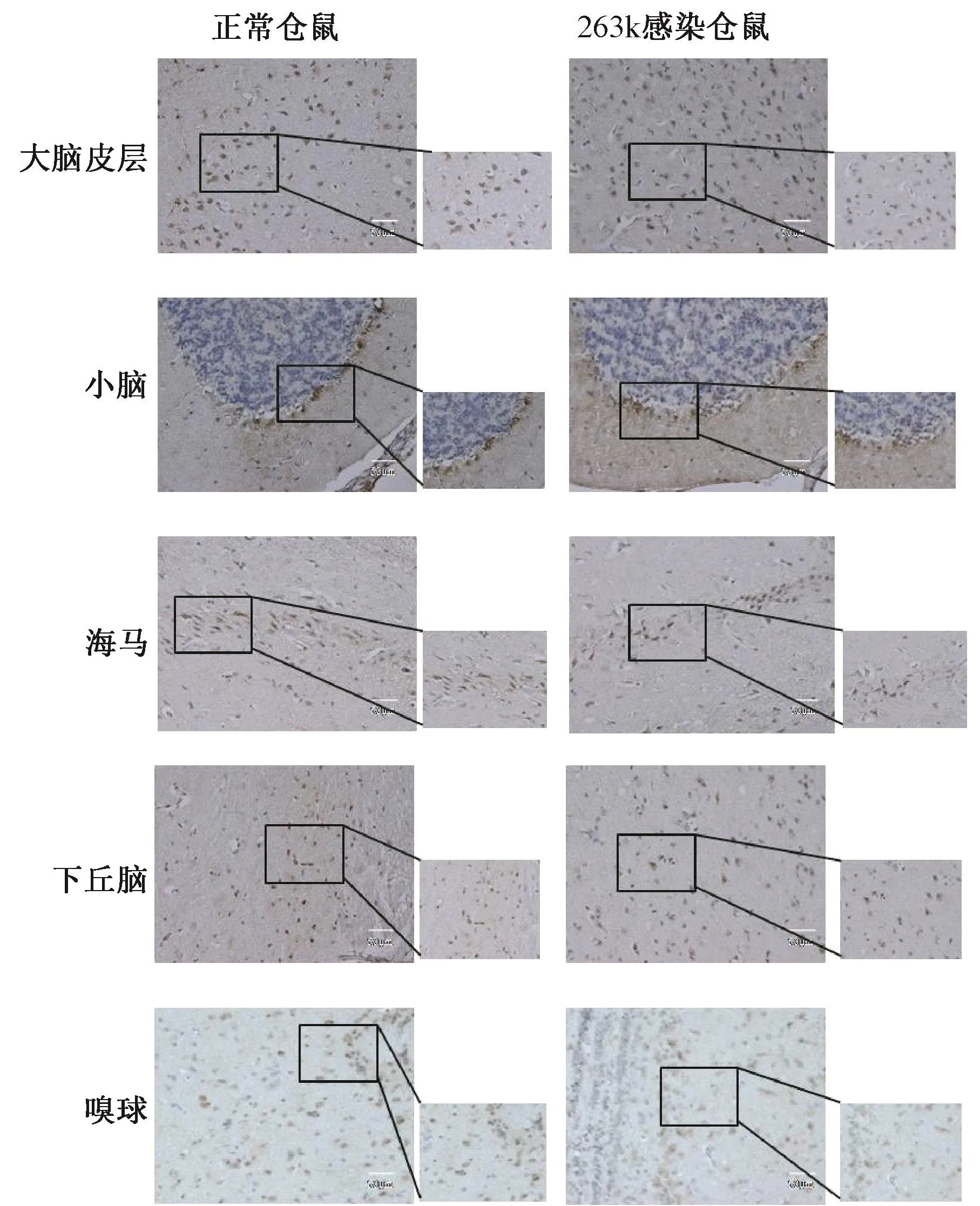
圖1 免疫組織化學檢測朊病毒263 K毒株感染倉鼠及正常倉鼠腦組織中RyR2表達(×400)Fig.1 Immunohistochemical detection of RyR2 in scrapie-263 K infected and normal hamster brain tissue(×400)
3 討論
RyR家族共有3種亞型:RyR1-3。在外周組織中,RyR1主要分布在骨骼肌;RYR2主要分布在心臟平滑肌;RyR3主要分布在一些上皮細胞、胃組織細胞及全身其他器官。而在中樞神經系統當中,3種亞型均有表達,其中 RyR2的表達最為豐富。RyR2主要分布于神經元細胞的內質網中,由同源四聚體組成,每個單體相對分子質量約為550×103。目前已有多種物質被證實與RyR2的活性有關,如FKBP、鈣離子、鈣調蛋白(CaM)、蛋白激酶 A(PKA)等[4]。
RyR2在神經元的樹突、胞體、突觸中均有分布[5]。在此前的文獻報道中發現,哺乳動物中,RyR2主要分布在大腦皮層和小腦浦肯野細胞層,并在海馬、丘腦等部位也有表達。除此之外,在嗅球、視神經、腦干等均發現RyR2的分布[6]。本研究利用免疫組織化學的方法探索RyR2在倉鼠腦組織中的分布,發現在正常和感染倉鼠的腦組織中,在大腦皮層、小腦、海馬、嗅球、下丘腦均發現RyR2的表達,這與此前文獻的報道基本符合。
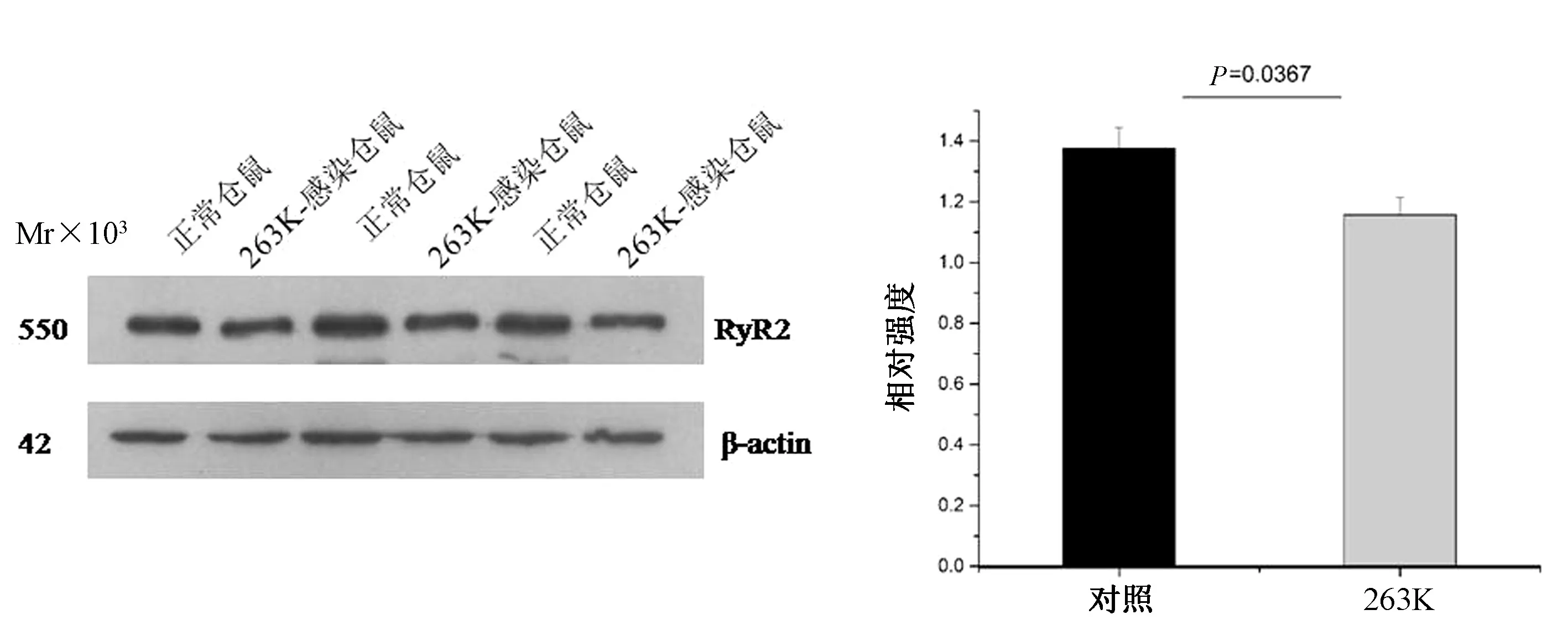
圖3 朊病毒263 K毒株感染倉鼠腦組織中RyR2表達含量與正常倉鼠比較Fig.3 RyR2 expression in the brain of scrapie-263 K infected was decreased compared with normal hamster
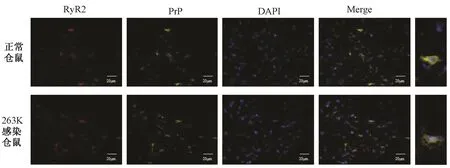
圖4 免疫組織熒光檢測顯示,感染倉鼠終末期腦組織中RyR2與PrP存在共定位關系(×400)Fig.4 Co-localization between RyR2 and PrP in scrapie-263 K infected and normal hamster brain tissue(×400)
RyR2參與到記憶以及學習的形成過程,可以將不穩定的記憶整合成強大、持久的記憶痕跡。受過水迷宮訓練的大鼠,可以發現其海馬中RyR2表達增高[7]。給小鼠腦室內注射RyR2拮抗劑,可以發現小鼠的記憶能力較對照組下降[8]。其次,RyR家族還與藥物成癮、社交活動、情緒變化等癥狀表現關系密切[9-11],這些都體現了RyR2在高級皮層功能方面的重要作用。因此,在正常生理狀態情況下,大腦中RyR2的存在對于認知功能的形成有重要作用,RyR2的表達減少或者功能抑制會導致記憶力的損害。而快速進展的認知功能下降正是人類朊病毒病最常見的癥狀之一。本研究發現,RyR2的表達在朊病毒感染動物模型中明顯下降,提示其可能與認知功能的快速下降有關。
除此之外,RyR2與神經變性疾病也有密切的聯系。大部分研究都認為在阿爾茲海默病中,RyR2的表達較正常上升。Bruno等發現,在輕度認知障礙和阿爾茲海默病(Alzheimer disease,AD)患者的額葉腦組織中,都發現了RyR2的mRNA表達量的升高[12],提示RyR2可能在認知功能障礙的早期即參與到疾病發展的過程中。Liang還發現,在AD模型小鼠中,接受RyR拮抗劑丹曲林處理的小鼠,其認知功能較對照組明顯改善,海馬區的老年斑形成也相對減少[14]。但也有少量研究得出了與之相悖的結果。Kelliher等[15]發現,在早期AD患者的腦組織中,RyR2的表達量明顯減少。Andrea等則發現,在Aβ處理誘導的海馬神經細胞株中,RyR2的表達減少[16]。在前期工作中發現RyR2在人遺傳性朊病毒病腦組織中表達明顯上調。而在263 K感染倉鼠動物模型中發現了相反的變化趨勢,這說明RyR2在人和其他哺乳動物中可能發揮著不一樣的功能;其次,這也說明仍需要更多的研究嘗試,從不同的角度驗證RyR2在朊病毒病中的表達變化。有趣的是,在心血管疾病中的研究領域中,不同學者對RyR2的研究也出現了這種互相矛盾的現象[16]。除此之外,在帕金森病、亨廷頓疾病、脊髓小腦共濟失調等遺傳相關的神經變性病中,也發現了RyR2表達或者鈣離子代謝紊亂的問題[17-18]。
雖然RyR2在不同神經變性疾病中的表達變化有所爭議,但普遍認為,RyR2介導了淀粉樣蛋白對神經元細胞的損傷過程。學者們猜測,可能是包括Aβ、PrP在內的淀粉樣蛋白通過刺激神經元細胞,造成內質網應激狀態,導致內質網RyR2功能的紊亂,大量鈣離子從內質網流入胞漿內,破壞神經細胞內鈣離子穩態,進而導致神經細胞的凋亡[15]。因此,在朊病毒感染倉鼠模型中,RyR2的表達出現了明顯的下降,其中的原因可能與PrPSc大量復制后影響RyR2的功能有關。除此之外,朊病毒病感染動物終末期的腦組織中,神經元的大量丟失也可能是導致RyR2表達含量下降的重要原因之一。
RyR2對于高級皮層功能的發育和完善有重要的作用,并介導參與了神經退行性病變的過程,RyR2的功能紊亂或者表達異常會影響神經細胞功能,甚至導致其凋亡。雖然目前已有學者探索RyR2作為靶點用于治療阿爾茲海默病,但考慮到RyR2在兩類疾病中這種截然相反的變化趨勢,提示應用RyR2拮抗劑治療朊病毒病的研究方向中,可能需要針對不同型別進行精準研究。這種差異對研究RyR2與朊病毒病的相互關系起到了重要的參考作用。
利益沖突 無
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
