利用崗46B/A232重組自交系群體分析葉綠素含量相關QTL
2018-12-11 03:29:16李永洪李傳旭劉成元向箭宇
西南農業學報 2018年11期
李永洪,李傳旭,劉成元,何 珊,向箭宇,謝 戎*
(1.四川省農業科學院水稻高粱研究所/農業部西南水稻生物學與遺傳育種重點實驗室/國家水稻改良中心瀘州分中心,四川 德陽 618000;2.華中農業大學生命科學技術學院,湖北武漢 430070)
【研究意義】水稻葉片是光合作用最重要的器官,其發育形態和葉綠素含量直接決定了產量的高低,水稻產量的90% ~95%來自于葉片的光合作用產物[1]。【前人研究進展】前人研究表明,葉綠素是植物吸收、傳遞和轉換光能最主要的色素,在一定范圍內,葉綠素含量和光合速率呈正相關關系,而光合速率的高低又直接決定了產量的高低[2-5]。由于氮素是葉綠素的主要組成部分,氮肥不足,葉綠素含量低,造成葉片短小,從而導致減產。但過多的施用氮肥導致氮素利用率較低,不僅加大農業生產成本,也帶來嚴重的環境問題[6-8]。因此,在不影響產量和品質的前提下控制化學肥料的施用量,提高肥料的利用效率,是現代農業的必然要求。葉片葉綠素含量的差異,反應了不同基因型對氮素的吸收狀況與葉綠素合成效率的差異,也有利于與氮素吸收相關基因的發掘。實際生產和研究工作中,常用葉綠素含量的高低來衡量植株營養水平,光合作用能力和逆境脅迫狀況[9-10]。與水稻許多農藝性狀一樣,水稻葉片葉綠素含量也是數量性狀。近年來,不同研究者利用不同遺傳群體對水稻葉片葉綠素含量QTL進行大量的定位研究,為闡明葉綠素的分子遺傳機制和高光效育種奠定了基礎。目前定位出的葉綠素含量相關 QTL較多,但被克隆的卻較少。qLSCHL4是一個控制葉形和葉綠素含量的主效QTL(qLSCHL4),圖位克隆和表達分析表明qLSCHL4是NAL1的等位基因,不僅控制葉形和葉綠素含量,還影響每穗粒數,屬一因多效[11]。研究發現Ghd7是自然變異控制葉綠素含量的主效QTL,Ghd7負向調節葉綠素含量[12]。一個在低氮條件下影響葉綠素含量的主效QTLqFCC7L,可能與水稻耐低氮脅迫能力有關,并將其定位在第7染色體長臂124.5 kb的區間內,對其候選基因分析表明,小肽轉運蛋白基因OsPRT4(LOC-Os07g41250)最有可能與qFCC7L有關,雙親在候選基因上存在序列變異和基因表達量的差異,OsPRT4主要在葉片和根中表達,這也與推測其具有吸收和轉運氮素的能力相符合[13]。【本研究切入點】本研究小組發掘的秈稻種質A232,中抗多種水稻害蟲[14]。利用該材料進行抗蟲轉育研究是本研究團隊近年來的主要研究內容之一。本研究擬利用“崗46B/A232”重組自交系群體,進行不同年度及不同葉位葉片葉綠素含量的QTLs分析,【擬解決的關鍵問題】以期為水稻高光效育種提供新的基因資源,為抗蟲種質A232的雜交轉育及組配出高產優質抗蟲品種提供分子標記及指導依據。
1 材料與方法
1.1 供試材料
以崗46B為母本,A232為父本進行雜交,按“一粒傳”法構建重組自交系[15],本研究以重組自交系群體的173個家系和雙親作為試驗材料。
1.2 田間試驗及葉綠素測定
田間試驗分別于2016,2017年在四川省農業科學院水稻高粱研究所瀘縣福集鎮試驗基地進行。2年均于當年3月20日播種所有試驗材料,秧齡25 d左右移栽。每個家系種植5行,每行10株,株行距為20.0 cm×26.7 cm。分別于分蘗期(5月30日)和抽穗期(7月15日)用便攜式葉綠素儀(SPAD-502.Monito,Japan)測定兩親本及各家系主莖的倒1、2和3葉葉片的SPAD值。測量時,取葉片中部位置重復測定3次,每個家系測定10片葉,取平均值用于換算該家系葉片葉綠素含量。計算分蘗期倒1、2和3葉葉綠素含量(分別簡稱 CTFL、CTSL和CTTL),抽穗期倒1、2和3葉葉綠素含量(分別簡稱CHFL、CHSL和CHTL),葉綠素儀測定的SPAD值與葉綠素含量之間的換算公式為:Y=0.1285X-0.5656,式中,X為葉綠素儀測定的SPAD值,Y為葉綠素含量(mg·dm-2)[16]。
1.3 遺傳圖譜構建和QTL分析
采用在兩親本間存在差異且帶型清晰的130對SSR引物對RIL群體株系進行基因型分析,構建SSR標記遺傳連鎖圖譜[15]。利用 ICIM(QTL Ici-Mapping)軟件對所考查的葉片葉綠素含量性狀進行QTLs定位分析,判斷QTL存在的閥值LOD≥2.50。QTL命名方法參照McCouch等提出的方法[17]。
2 結果與分析
2.1 RIL群體葉綠素含量與產量性狀間的關系
從表1可見,各葉綠素含量性狀間呈極顯著正相關,與單穗重直接相關系數不大,抽穗期倒3葉葉綠素含量與谷粒厚兩年中均呈極顯著正相關關系,分蘗期倒2葉葉綠素含量與谷粒厚兩年中分別呈極顯著和顯著正相關。其余各葉綠素含量性狀與各粒形性間的相關性在2017年均不顯著。而在2016年中,分蘗期倒2葉葉綠素含量與千粒重達顯著正相關,分蘗期倒3葉葉綠素含量與谷粒厚達極顯著正相關,抽穗期倒1葉葉綠素含量與谷粒長和谷粒長寬比達顯著正相關;抽穗期倒2葉葉綠含量與谷粒長呈顯著正相關,與谷粒厚和千粒重呈極顯著正相關。
2.2 RIL群體及其親本的葉綠素含量性狀的表型變異
由表2可知,在分蘗期和抽穗期2個生長階段,2年中雙親頂部三片葉的葉綠素含量存在明顯差異,崗46B高于A232。在RILs群體中,不同家系間葉綠素含量變異范圍較大,上三葉葉綠素含量均存在超親分離,偏度和峰度分析表明,2年上三葉葉綠素含量均表現近正態的連續分布,表明葉片葉綠素含量為多基因控制的數量遺傳,可以進行QTL定位分析。
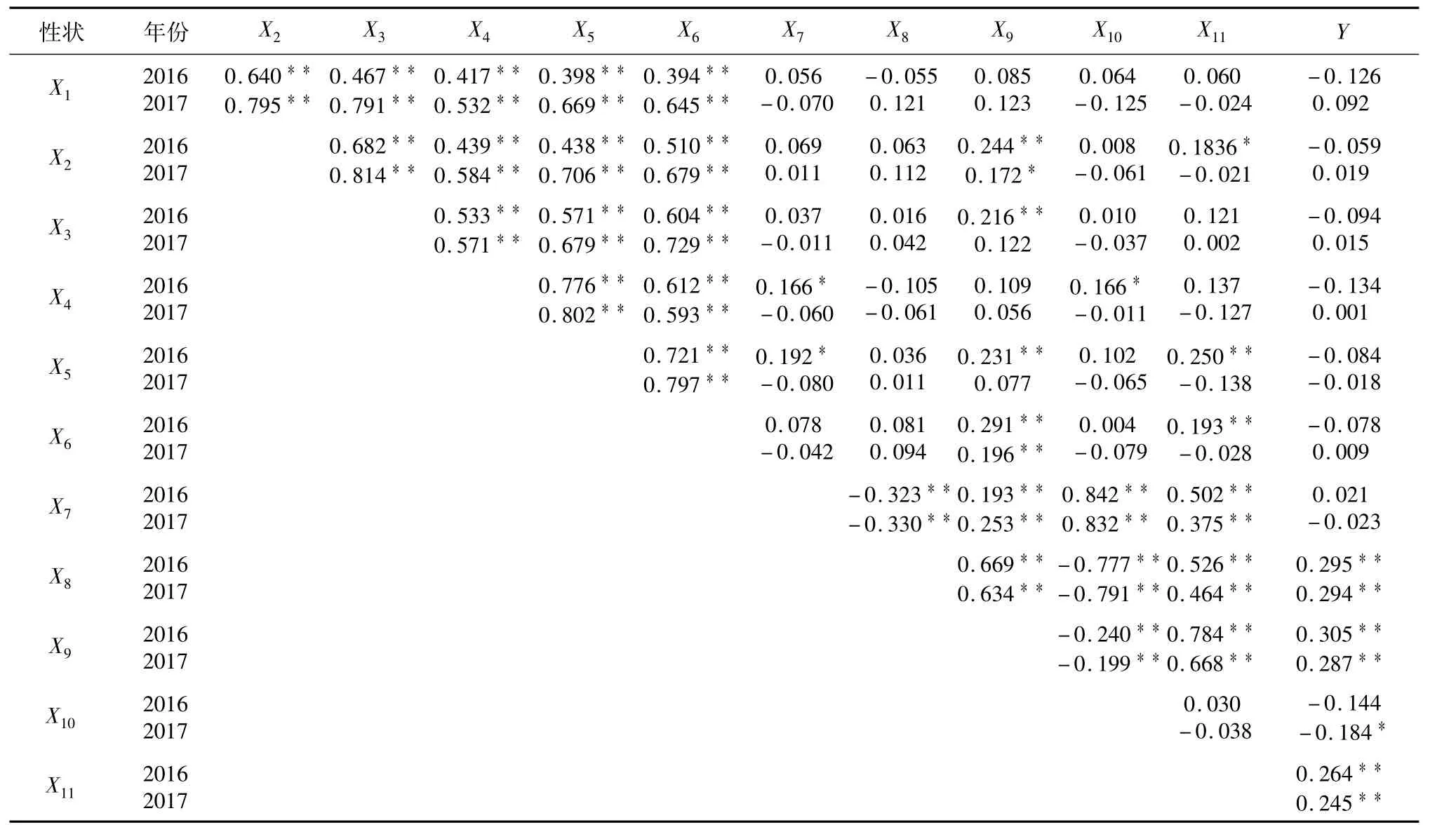
表1 各葉綠素含量性狀與粒形及產量性狀的相關分析Table 1 Correlation coefficients among chlorophyll contents and grain shape traits and grain yield in the Gang46B/A232 RIL population
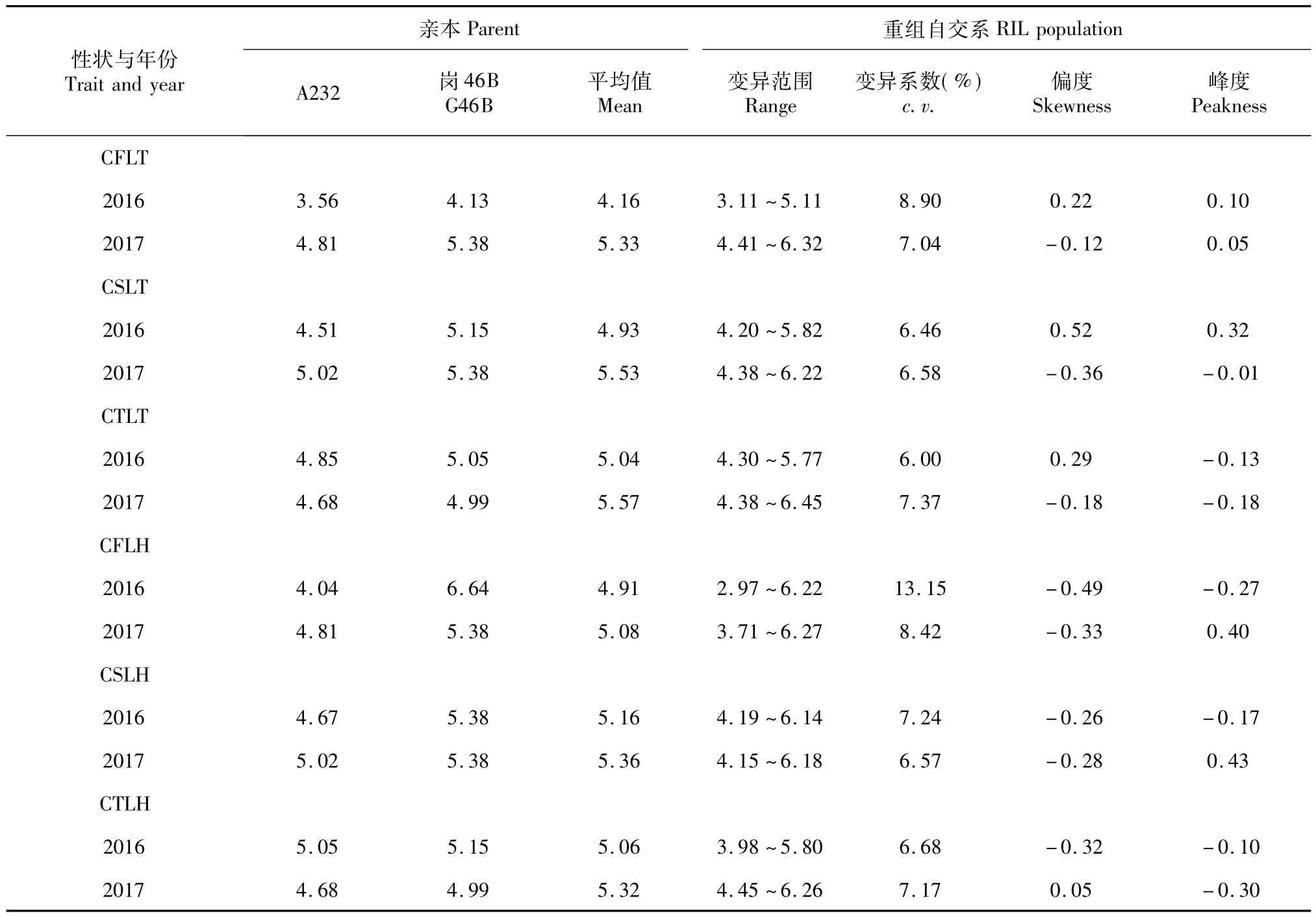
表2 親本與重組自交系群體各時期的葉綠素含量表現與分布Table 2 Phenotypic variation and distribution of chlorophyll content in the RIL population and its parents at different stages
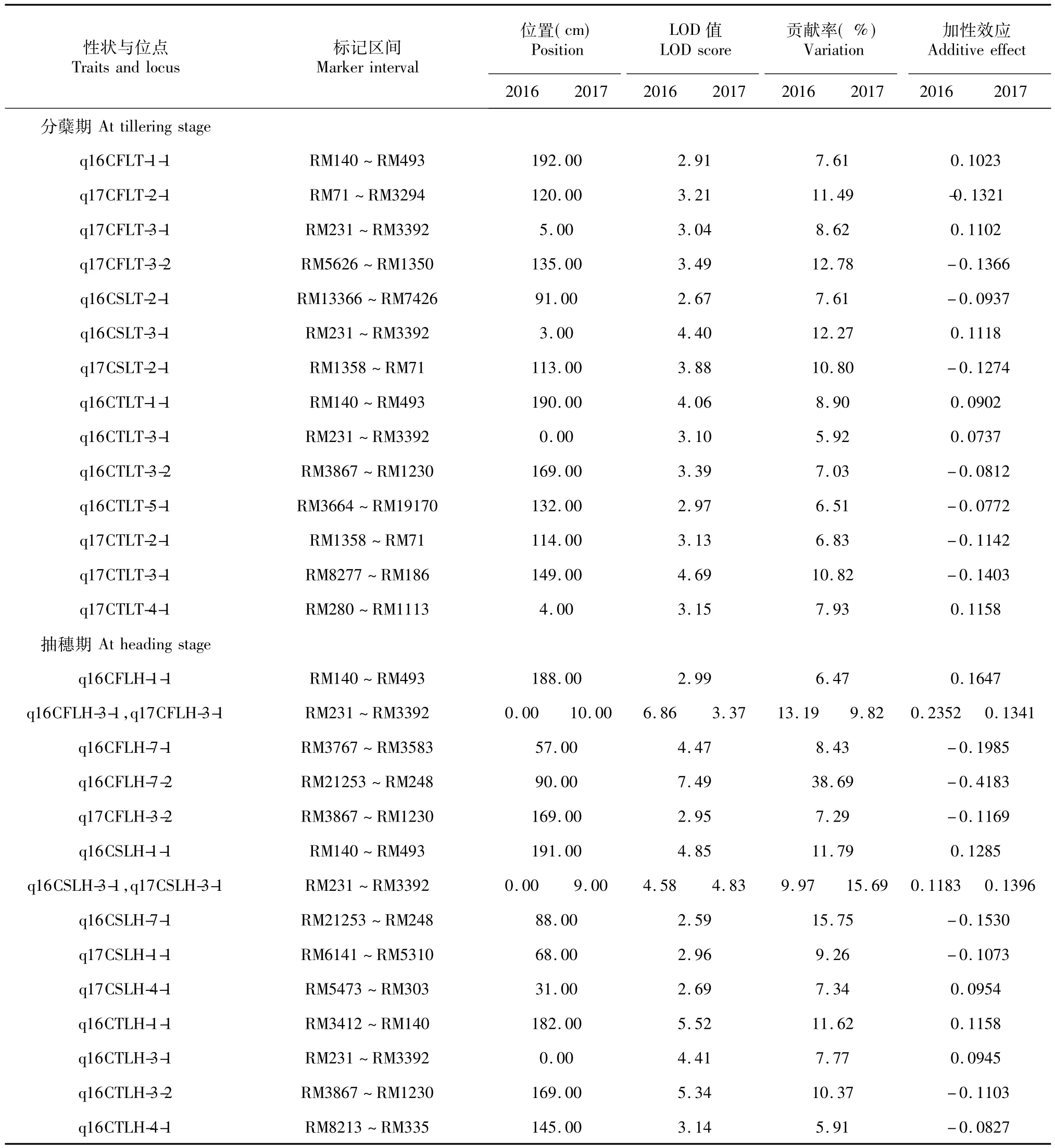
表3 RIL群體葉片葉綠素含量的QTL位置與遺傳效應Table 3 Identification of QTL for chlorophyll content of leaf in RIL population and their genetic parameters estimated
2.3 利用復合區間作圖法對葉綠素含量的QTL檢測
利用完備區間作圖法對崗46B/A232的RIL群體進行葉綠素含量QTL分析,2016年共檢測到18個 QTL,分布于第1、2、3、4、5 和7 染色體,LOD 值介于2..59 ~7.49,單個 QTL 分別解釋相應性狀變異的5.91% ~38.69%。對2017年分蘗期和抽穗期調查的頂部3片葉的葉綠素含量進行分析,共檢測到12個 QTL,分別位于第1、2、3和4染色體上,LOD值介于2.95~4.83,分別解釋葉片葉綠素含量變異的6.83% ~15.69%(表3)。
2016年控制分蘗期和抽穗期倒1、2和3葉葉綠素含量的QTL在第3染色體RM231-RM3392區間上,而根據2017年表型值在此區間上只有抽穗期的倒1葉和倒2葉葉綠素含量QTL被檢測到。2016年檢測到在分蘗期和抽穗期控制倒3葉葉綠素含量QTLq16CTLT-3-1和q16CTLH-3-2,位于第3染色體RM3867~RM1230區間上,但根據2017年葉綠素表型值,在此區間上檢測到的是控制抽穗期倒1葉葉綠含量的QTLq17CFLH-3-2。根據2016年表型值,在第1染色體RM140-RM493區間上檢出的有控制分蘗期倒1葉葉綠素含量QTLq16CFLT-1-1、分蘗期倒3葉葉綠素含量QTLq16CTLT-1-1、抽穗期倒1葉葉綠素含量QTLq16CFLH-1-1和抽穗期倒2葉葉綠素含量QTLq16CSLH-1-1,但根據2017表型數據,未能在此區間檢測葉綠素含量相關性狀QTL。在第2染色體 RM1358~RM71區間,只有2017年的表型值在此區間上檢出q17CSLT-2-1和q17CTLT-2-1 2個葉綠素含量相關QTL。在抽穗期,2016年表型值檢出4個控制倒三葉葉綠素含量QTL,但根據2017的表型值卻未能檢測到控制倒三葉葉綠素含量的QTL。
3 討論
3.1 葉綠素含量與產量
葉綠素是水稻進行光合作用的物質基礎,改善葉片葉綠素含量可以提高光合效率,進而提高水稻產量和品質。本研究簡單相關分析表明,各葉綠素含量相關性狀與單穗重并無顯著相關關系,但其與產量密切相關的粒形性狀和千粒重呈現出各種復雜相關關系,有的相關性達顯著或極顯著水平。可見,不同葉位葉綠素含量與水稻產量有著密切的關系[18-19],并且可能影響稻谷的品質。
3.2 本研究定位的葉綠素含量QTL與前人定位結果的比較
本研究檢測到的控制分蘗期倒1葉葉綠素含量QTLq17CFLT-3-2(RM5625~RM1350)與孫小霞等檢測到的葉綠素含量QTLqChl-3c(RM565~RM168),姜樹坤等檢測到的分蘗期葉綠素含量QTLqCT3a(PM380~RM426)染色體位置區間部分重疊[20-21]。
胡頌平等定位的葉綠素含量QTLqCC3c(RM22~RM231),范淑秀等定位的葉綠素含量QTLqChla-3b、qChlb-3b、qChlt-3b 和 qChlxc-3b(RM231 ~RM22)與本研究定位到的葉綠素含量相關QTL區間(RM231~RM3392)具有相近或相同的染色體位置[22-23]。本研究中,該區間兩年共有8次被檢出,說明該位點上控制葉片葉綠素含量的基因受環境影響較小,表達穩定。
孫小霞等檢測到的qChl-4(RM470~RM451),胡頌平等定位的qCC4b(RM255~RM349),Jiang等定位的 qCH4,qCM4b和qCTH4(RM255~RM559)及Zhang等定位的qLSCHL4,阿加拉鐵等檢測到的qCHL4-1(RM280~RM1113)、qCHL4-2(RM255~RM280)與本研究檢測到的葉綠素含量QTLq17CTLT-4-1(RM280~RM113)和q17CSLH-4-1(RM5473~RM303)具有相同的染色體位置或位置部分重疊[11,20,22,24-25]。
姜樹坤等定位的 qCH7(RM478~RM429)、qCM7a(RM214)和 qCM7b(RM18~RM429)區間,范淑秀等定位的 qChla-7、qChlb-7、qChlt-7和qChlxc-7(RM18~RM118)區間,Ye等定位的在低氮條件影響頂三葉葉綠素含量的主效QTLqFCC7L均位于本研究檢測到的q16CFLH-7-2(RM21253~RM248) 區間上[13,21,23]。本研究中,該位點只在2016年表型值中被檢測到,且其遺傳效應值較大,該位點的表達是否受低氮條件的影響,需進一步確認。
3.3 葉綠素含量QTL的多樣性及表達的穩定性
定位用的遺傳群體、分子標記種類及QTL作圖方法的不同,以及性狀值受環境條件影響等因素,前人對水稻葉片葉綠素含量QTL定位研究結果差異較大。目前定位出的葉綠素含量相關QTL較多,但被克隆的卻較少。有研究表明,不同生長階段和不同葉位葉綠素QTL的表達表現出明顯特異性,如QTLqFCC7L在始穗期低氮條件影響頂三葉葉綠素含量,在抽穗期低氮條件下也控制倒一和倒三葉葉綠素含量[13]。本研究定位在第3染色體RM5625~RM1350區段和RM231~RM3392區段,第4染色體RM280~RM1113區段和RM5473~RM303區段,第7染色體RM21253~RM248區段的葉綠素含量QTL位點與前人研究結果一致[11,13,20-25],說明這些區間上的QTLs受環境影響較小,在水稻不同生長階段能穩定表達,有利于提高葉片葉綠素含量,增強光合作用,從而能提高水稻產量和品質。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
今日農業(2021年14期)2021-10-14 08:35:40
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
文苑(2020年6期)2020-06-22 08:41:52
