免疫補體調(diào)節(jié)蛋白C1抑制物對巨噬細胞極化的作用①
2019-01-03 06:10:36吳世舜余文博劉夢元張海謀劉東旭
中國免疫學(xué)雜志 2018年12期
關(guān)鍵詞:功能
吳世舜 余文博 劉夢元 張海謀 熊 君 劉東旭
(湖北大學(xué)生命科學(xué)學(xué)院,武漢 430062)
在體內(nèi),炎癥引起組織損傷可迅速導(dǎo)致?lián)p傷區(qū)巨噬細胞數(shù)量增加,在很大程度上,主要依賴于單核-巨噬細胞系的極化,這種變化主要是由于血中單核細胞浸潤,成為靜息巨噬細胞。這些靜息巨噬細胞在細胞因子的作用下,轉(zhuǎn)變?yōu)椴煌偷臉O化巨噬細胞[1]。在體外,巨噬細胞集落刺激因子(CSF)或粒細胞-巨噬細胞集落刺激因子(GM-CSF)均可促使單核細胞分化為靜息巨噬細胞,而干擾素-γ(IFN-γ)和白細胞介素4(IL-4)+IL-13可誘導(dǎo)靜息巨噬細胞分別轉(zhuǎn)化為經(jīng)典活化的巨噬細胞(M1)和替代激活的巨噬細胞(M2)[2,3]。巨噬細胞呈現(xiàn)明顯的差異性極化能力,可隨具有不同功能表型之間動態(tài)變化而相互轉(zhuǎn)換[4]。人巨噬細胞表型的表達CD14、CD163、CD206是區(qū)分M1和M2的重要標記物[5],也是巨噬細胞呈現(xiàn)明顯極化能力的標志。M1產(chǎn)生大量的具有促炎性特性的細胞因子如腫瘤壞死因子-α(TNF-α)、IL-6和趨化因子(C-C motif)配體17(CCL17)及相關(guān)的酶如誘導(dǎo)型一氧化氮合酶(iNOS)。而M2則產(chǎn)生大量的具有抗炎作用物質(zhì)如15脂氧合酶(ALOX15)、IL-10和CCL18和精氨酸酶1(Arg1)[4,6,7]。
補體調(diào)節(jié)蛋白C1抑制劑(C1INH)是補體系統(tǒng)中經(jīng)典途徑唯一蛋白酶抑制劑,C1INH 存在于人體血漿中。巨噬細胞也是參與執(zhí)行補體系統(tǒng)中途徑效應(yīng)功能的關(guān)鍵因素。補體系統(tǒng)的成分對巨噬細胞極化起調(diào)節(jié)作用。已有研究表明,補體蛋白C1q可以直接巨噬細胞極化[8]。補體成分C1q、C3b和C5a與巨噬細胞上的受體相互作用,導(dǎo)致炎癥反應(yīng)和細胞因子的產(chǎn)生。C3a、C5a和C5b-9調(diào)節(jié)巨噬細胞趨向炎性M1表型,促炎細胞因子的釋放,而C3b、C1q調(diào)節(jié)巨噬細胞趨向抗炎性M2表型,抗炎細胞因子的產(chǎn)生,這些變化可能與細胞信號傳導(dǎo)作用有關(guān)[9]。既往研究已經(jīng)證實:C1INH具有抗炎的新功能,抑制巨噬細胞激活和炎性細胞因子釋放[10-12]。
本研究將分析C1INH對極化的巨噬細胞表型標志物CD14、CD163和CD206作用,研究C1INH對極化巨噬細胞產(chǎn)生促炎因子和抗炎因子的作用,同時,探索C1INH對極化巨噬細胞殺菌活性和吞噬功能的作用。進一步分析C1INH對CD14結(jié)合Toll樣受體4(TLR4)的影響。從而闡明C1INH對巨噬細胞極化的調(diào)節(jié)作用。
1 材料與方法
1.1實驗材料、試劑 Ficoll PaqueTMPLUS單核細胞分離液購自immunbioTM;M-CSF、GM-CSF、IFN-γ,IL-4,IL-13購自R&D;C1INH購自Berinert,ZLB Behring;Anti-CD14、anti-CD163,anti-CD206抗體,R&D;磷酸化的RAF-1、MEK-1和p42/p44ERK抗體購自NEB;RNA Mini Kit和PCR Kit購自Qiagen;96孔平底板購自BD Biosciences;免疫反應(yīng)-增強化學(xué)發(fā)光檢測系統(tǒng)檢測購自Pierce Biotechnology;pHrodoTMGreen E.coli BioParticlesTMConjugate和LPS購自ThermoFisher。
1.2實驗方法
1.2.1細胞分離和誘導(dǎo) 健康成人外周血液樣本的研究得到湖北大學(xué)生命科學(xué)學(xué)院審查委員會的批準,所有捐助者都知情同意。應(yīng)用17-1140-02 Ficoll PaqueTMPLUS 單核細胞分離液分離人外周血單個核細胞,分離所得的細胞中95%±5%為單核細胞,分離所得的細胞存活細胞率>90%。細胞培養(yǎng)濃度在1×106個細胞/ml,細胞在完全培養(yǎng)基含10%胎牛血清,100 U/ml青霉素/鏈霉素,加入50 ng/ml M-CSF或50 ng/ml GM-CSF,培養(yǎng)3 d后,再添加到含有生長因子的新鮮培養(yǎng)基中。培養(yǎng)7 d后,細胞用1×PBS含Ca2+和Mg2+和新鮮的完全RPMI洗2次,再加入完全培養(yǎng)基含有100 ng/ml IFN-γ(M2)或10 ng/ml IL-4+10 ng/ml IL-13(M2),培養(yǎng)48 h。在預(yù)實驗中,設(shè)立C1INH濃度為0、50、100、150、200 μg/ml,同時也設(shè)立C1INH作用時間0、12 h、24 h、48 h。因此,最后選定C1INH終濃度和作用時間為150 μg/ml和48 h。在上述實驗組中,加入150 μg/ml C1INH、24 h作用時間,設(shè)立陰性對照組。通過檢測和鑒定,M-CSF或GM-CSF誘導(dǎo)的巨噬細胞的純度分別為95%和96%。
1.2.2流式細胞儀檢測 在培養(yǎng)的細胞加入抗體(CD14、CD163,CD206)進行孵育,洗滌后再進入二抗。細胞用PBS含Ca2+和Mg2+和新鮮的完全培養(yǎng)基洗2次,細胞固定后,進行流式細胞儀檢測(BD),應(yīng)用CellQuest軟件分析結(jié)果。
1.2.3RT-PCR 從細胞中提取總RNA,并通過測量其在260 nm處的吸光度來定量測定總RNA的濃度,PCR Kit進行qPCR分析,GAPDH作為內(nèi)部對照引物,具體操作過程按試劑說明書要求。反應(yīng)體系在95℃預(yù)變性30 s,40循環(huán)擴增反應(yīng)(95℃ 5 s,60℃ 34 s),熔解曲線分析,結(jié)果與內(nèi)參作對比。
1.2.4細胞殺菌試驗 按以前方法操作[11]。簡而言之,3×104細胞/孔接種于纖連蛋白涂覆的96孔平底板培養(yǎng)2 d。大腸桿菌生長對數(shù)中期(OD600=0.5)的RPMI1640培養(yǎng)基含有10 nmol/L HEPES,細胞數(shù)與等量的大腸桿菌數(shù)目在37℃下共培養(yǎng),并加入150 μg/ml C1INH,設(shè)立對照組。用冰冷的水和渦旋裂解細胞。在37℃下,將裂解物連續(xù)稀釋并涂于含5% FBS的瓊脂基片上,培養(yǎng)后計數(shù)活菌數(shù),測定存活細菌的百分比。
1.2.5細胞對菌的吞噬試驗 按說明書(生命技術(shù))方法操作,用FITC標記大腸桿菌,進行細胞吞噬試驗。FITC標記的細菌生長到對數(shù)中期(OD600=0.5)的RPMI1640培養(yǎng)基含有10 nmol/L HEPES。隨后,細胞與等量的大腸桿菌數(shù)目在37℃下共培養(yǎng),并加入150 μg/ml C1INH。細胞用PBS洗兩次去除非吞噬物質(zhì),刮掉細胞,然后通過流式細胞儀檢測細胞吞噬FITC標記物的情況。
1.2.6Western blot 離心分離細胞裂解液,在4℃下過夜,1 mg蛋白質(zhì)用合適的抗體和蛋白A/G加瓊脂糖孵育。蛋白用10%聚丙烯酰胺分離凝膠電泳分離,然后轉(zhuǎn)移到硝酸纖維素膜上。膜上蛋白質(zhì)與相應(yīng)抗體(磷酸化的RAF-1、MEK-1和p42/p44ERK抗體)結(jié)合,洗滌后,加入辣根過氧化物酶抗體。免疫復(fù)合物通過增強化學(xué)發(fā)光檢測系統(tǒng)檢測。

2 結(jié)果
2.1C1INH對M1和M2極化表型的作用 CD14是M1的表面標記物,CD163是M-CSF源性巨噬細胞分化表面標記物,CD206是IL-4誘導(dǎo)的M2表面標記物[5,13,14]。在靜態(tài)狀態(tài)下,C1INH抑制M-CSF源性巨噬細胞的CD14、CD163和GM-CSF源性巨噬細胞的CD206表面標記物的表達(圖1)。在極化狀態(tài)下,C1INH抑制M1的CD14、CD163和M2的CD206表面標記物的表達(圖2)。結(jié)果提示:C1INH對巨噬細胞極化表面標志物具有調(diào)節(jié)作用。
2.2C1INH對M1和M2的標記基因表達的影響 IFN-γ或IL-13誘導(dǎo)巨噬細胞極化不同的基因表達趨化因子和細胞因子是有差異的[5]。我們發(fā)現(xiàn)C1INH抑制M1的TNF-α和IL-6表達,而增加M2的15-脂氧合酶和IL-10表達(圖3A~D)。C1INH抑制M1 iNOSmRNA和提高M2 Arg1 mRNA表達(圖3E、F)。 結(jié)果提示:C1INH具有調(diào)節(jié)M1和M2的標記基因趨化因子和細胞因子分泌及相關(guān)酶表達。
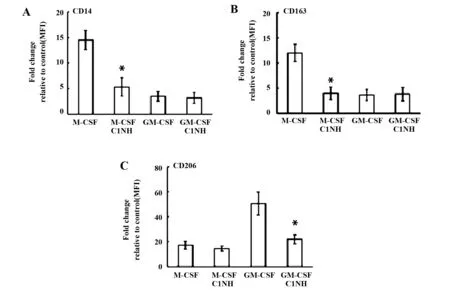
圖1 C1INH對靜息巨噬細胞(CD14、CD163和CD206)表型的影響Fig.1 Effect of C1INH on phenotype(CD14,CD163 and CD206)of resting macrophagesNote: FITC-conjugated antibodies for CD14 (A),CD163 (B),and CD206 (C) were used.Flow cytometric measurement of surface marker expression depicted as fold change of mean fluorescence intensity (MFI) relative to the control in box plots for macrophages (n=6).*.P<0.05.
2.3C1INH提高極化巨噬細胞殺菌活性和對菌的吞噬功能 M1具有很高的殺菌活性[4,6,7],而M2顯示吞噬功能比M1更大[15,16]。在實驗中重點研究C1INH對M1的殺菌活性和M2的吞噬功能的作用。實驗中發(fā)現(xiàn):在巨噬細胞對菌殺滅功能中,C1INH明顯的增強M1的殺菌活性功能(圖4A)。在分析巨噬細胞對菌的吞噬功能中,C1INH明顯的增加M2對菌吞噬功能(圖4B)。
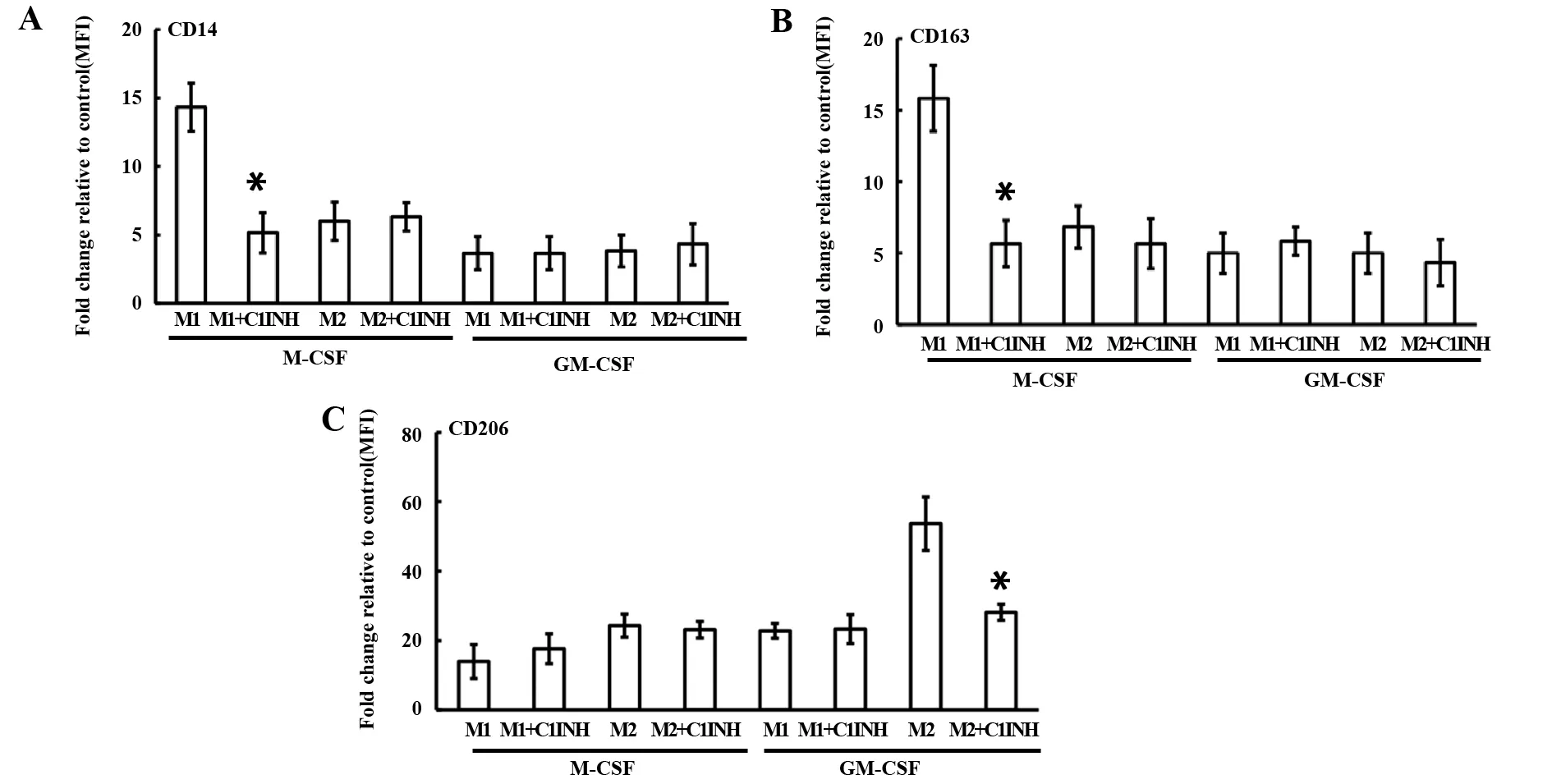
圖2 C1INH對極化的M1和M2(CD14、CD163和CD206)表型的影響Fig.2 C1INH regulates phenotype(CD14,CD163 and CD206)of M1 and M2 polarizationNote: FITC-conjugated antibodies for CD14 (A),CD163 (B),and CD206 (C) were used.Flow cytometric measurement of surface marker expression depicted as fold change of mean fluorescence intensity (MFI) relative to the control in box plots for M1 (n=6) and M2 (n=6).*.P<0.05.

圖3 C1INH對M1和M2標記基因表達的影響Fig.3 Effect of C1INH on expression of marker genes in M1 and M2Note: TNF-α(A),IL-6(B),ALOX15(C),IL-10(D),iNOS(E),and ARG1(F) were shown.Data are expressed as fold differences of polarized M1 (n=6) and M2 (n=6) with C1INH(n=6).*.P<0.05.

圖4 C1INH提高極化巨噬細胞的殺菌活性功能和對菌的吞噬功能Fig.4 C1INH improves bactericidal activity and phagocytosis of polarized macrophagesNote: A.Viable bacteria from M1 and M2 were detected and counted,and the percentage of surviving bacteria was determined;B.Phagocytosis tests were performed in M1 and M2 using FITC-labeled beads and then analyzed for the uptake of FITC-coupled beads by FACS.Data are expressed as fold differences of polarized M1 (n=6) and M2 (n=6) with C1INH (n=6),*.P<0.05.

圖5 C1INH抑制極化巨噬細胞的CD14-TLR4信號轉(zhuǎn)導(dǎo)Fig.5 C1INH inhibits CD14-TLR4 signal transduction in polarized macrophagesNote: Phosphorylation of RAF-1,MEK,p42/p44 ERK were assessed immunoblotting and an internal control.Representative immuno-blotting is shown for three experiments.
2.4C1INH抑制極化巨噬細胞CD14-TLR4介導(dǎo)的信號轉(zhuǎn)導(dǎo) CD14參與LPS誘導(dǎo)的TLR4信號通路,導(dǎo)致胞內(nèi)Raf-1/MEK/ERK激活[17,18]。實驗中發(fā)現(xiàn):通過Western blot證實C1INH抑制LPS誘導(dǎo)M1和M2的Raf-1/MEK/ERK (p44/42)的磷酸化的作用(圖5)。結(jié)果提示:C1INH具有抑制LPS誘導(dǎo)CD14-TLR4介導(dǎo)細胞信號通路激活的作用。
3 討論
C1INH是一個急性時相蛋白,是一種蛋白酶的抑制物,在患者致命感染性休克時,C1INH水平趨于正常的,但滅活的C1INH則增加,尤其感染性炎癥區(qū)C1INH水平可增加2.5倍。在C1INH缺乏,補體系統(tǒng)和相應(yīng)的激肽系統(tǒng)功能狀態(tài)就受影響[19-22]。已有研究證明,C1INH可減少細胞因子對補體系統(tǒng)激活的作用[23]。我們以前已證實:C1INH抑制LPS誘導(dǎo)的巨噬細胞系RAW264.7的TNF-α mRNA表達[10],C1INH抑制血液中或腹膜液中活細菌的生長,可通過增加中性粒細胞和巨噬細胞對細菌吞噬功能和殺細菌作用[11]。
單核細胞和巨噬細胞可表達補體受體CR1(CD35)、CR3(CD11b/CD18)和CR4(CD11c/CD18)。 補體成分可與巨噬細胞多種補體受體相互作用,導(dǎo)致細胞因子的產(chǎn)生和釋放,從而導(dǎo)致炎癥反應(yīng)[9]。IFN-γ誘導(dǎo)表型CD14的表達是M1的標記物,IL-4+IL-13誘導(dǎo)表型CD163的表達是M2的標記物[5]。在GM-CSF源性M2,CD206表型是增高的。CD206也可高表達在人單核細胞來源的樹突狀細胞上[24]。 在一些類似M2的細胞中,也可有表型CD206和CD163表達,呈現(xiàn)是一種抗炎表型的表達,但也能分泌一些促炎介質(zhì)[25]。但在某些疾病中,如哮喘患者,其M2的表型CD163的表達就比非哮喘正常巨噬細胞的表達較少[26]。
4 結(jié)論
在本研究中,C1INH對極化巨噬細胞的調(diào)節(jié)作用表現(xiàn)如下:①調(diào)節(jié)極化巨噬細胞表型標志物CD14、CD163、CD206再分布;②調(diào)節(jié)極化巨噬細胞趨化因子、細胞因子分泌和相關(guān)酶表達的改變;③增強極化巨噬細胞殺菌活性和對菌吞噬功能;④抑制CD14-TLR4介導(dǎo)信號通路途徑。因此,C1INH調(diào)節(jié)巨噬細胞極化,可能成為一種潛在的治療炎癥反應(yīng)新藥物。
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2020年1期)2020-02-20 13:23:44
經(jīng)濟技術(shù)協(xié)作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術(shù)與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學(xué)報(2016年1期)2017-01-20 08:21:20
中國中醫(yī)藥現(xiàn)代遠程教育(2014年22期)2014-03-01 04:32:55
