木欖幼苗對淹水-鹽脅迫的生理響應
2019-01-17 02:45:16黃麗譚芳林林捷樂通潮游惠明歐陽亞群
防護林科技 2018年12期
黃麗,譚芳林,林捷,樂通潮,游惠明,歐陽亞群
(福建省林業科學研究院,福建省森林培育與林產品加工利用重點實驗室,福建 福州 350012)
紅樹林生長于熱帶亞熱帶的潮間帶,具有重要的生態功能和經濟價值[1]。20世紀50年代,紅樹林在我國仍有較大面積的分布,約占25萬 hm2。但是,紅樹林灘涂的轉換性開發利用,如開墾灘涂以改造成魚塘等造成紅樹林分布大面積減少,紅樹林資源不斷枯竭[2]。目前,不僅要保護現有的紅樹林來維持其生態功能和經濟價值,還需要加強紅樹林的營造工作。紅樹林宜林地選擇主要受如下因子的限制:溫度、潮位、水體鹽度、底泥和潮水流速[1]。潮位是影響紅樹造林成活的關鍵。潮位高低直接決定紅樹植物周期性淹水的時間,潮位越低,紅樹植物周期性淹水的時間就越長。調查表明,低潮位灘涂上的紅樹植物的存活率低于中、高潮位[3]。水體鹽度影響紅樹植物幼苗生長的好壞,對幼苗的存活具有重要意義[4]。因此,開展紅樹植物對鹽度及淹水環境的適應性研究以確定最佳的水體鹽度和淹水時間,將為紅樹植物的造林提供重要的理論依據。前人關于淹水脅迫或鹽脅迫對紅樹植物幼苗生長影響的研究大多數是單一脅迫[5-8],而關于兩者的交互作用影響的研究為數不多[9,10]。木欖(Bruguieragymnorrhiza)是我國東南沿海一種重要的紅樹植物,也是紅樹造林的重要樹種。研究木欖幼苗對不同淹水-鹽脅迫的生理適應特性,將進一步揭示紅樹植物在淹水脅迫、鹽脅迫以及兩種脅迫交互作用下的抗逆性機制,為紅樹人工栽培提供一定的理論依據。
1 材料和方法
1.1 試驗材料和設計
木欖胚軸采自漳江口紅樹林國家級自然保護區。選取無病蟲害、生長狀況相近的木欖胚軸種植于洗凈的河沙上。試驗在福建省林業科學研究院潮汐模擬實驗室中進行。水體鹽度設置為0‰、5‰、10‰、15‰、20‰和30‰6個水平;在不同鹽度水平下,每個半日潮苗木被完全水淹的時間設置為0、2、4、6和8 h5個水平,共30個處理(表1),每個處理設置3個重復,每個重復30株苗。培養周期為3個月,每周更換1次鹽水,每天補充適量的自來水以保持鹽度和水位高度。培養結束后隨機選取木欖幼苗中上部健康成熟的葉片進行各項生理指標的測定。

表1 試驗組合設計表
1.2 試驗方法
質膜透性應用電導法測定[11];葉片丙二醛( MDA)含量應用硫代巴比妥酸法測定[12];超氧化物歧化酶( SOD )活性應用氮藍四唑( NBT )比色法測定,一個SOD酶活性單位U定義為抑制NBT光化還原的50%所需要的酶量[12];過氧化物酶( POD )活性應用愈創木酚法測定,以每分鐘A470變化0.01為一個POD酶活單位U[12];可溶性總糖含量應用蒽酮比色法測定[12];可溶性蛋白質含量應用Bradford法測定[13]。
1.3 數據統計與分析
應用 Excel 2010對數據進行圖表制作。應用 DPS 15.10軟件進行方差分析。
2 結果與分析
2.1 淹水-鹽脅迫對木欖幼苗葉片膜脂過氧化作用的影響
如圖1A所示,在低鹽(鹽度≤15‰)處理下,隨著淹水時間的增加,MDA含量基本呈現先增后降的趨勢。而在高鹽(鹽度為20‰~30‰)脅迫下,MDA含量隨淹水時間的增加,先降低后增加。另外,在同一個淹水脅迫處理下,隨著水體鹽度的增加,MDA含量基本均呈現先降后增的趨勢,在8 h淹水-20‰鹽脅迫下的MDA含量最大。同時,質膜透性的變化趨勢與MDA含量的一致(圖1B),在8 h淹水-20‰鹽脅迫下的質膜透性最大,高鹽脅迫下質膜透性均普遍高于低鹽脅迫。


圖1 淹水-鹽脅迫對木欖幼苗葉片膜透性的影響
對MDA含量和質膜透性進行方差分析,結果顯示木欖幼苗葉片MDA含量、質膜透性對淹水、鹽度及其交互作用影響具有極顯著性差異(P<0.01)。而且水體鹽度對木欖幼苗生長的影響比淹水時間更大(表2)。

表2 木欖幼苗在淹水-鹽處理下的MDA含量及質膜透性的方差分析
2.2 淹水-鹽脅迫對木欖幼苗葉片抗氧化酶活性的影響
如圖2A所示,當水體鹽度介于10‰和15‰之間時,隨著淹水時間的增加,POD活性呈現先降后升的趨勢,且均在淹水時間為8 h時POD活性最高。當水體鹽度<30‰,且淹水時間>4 h時,POD活性與淹水時間呈正相關關系。當水體鹽度位于10‰~20%區間時,短時水淹(2~4 h)使POD活性降低,長時水淹(6-8 h)則使POD活性增強。
如圖2B所示,在2 h淹水處理下,水體鹽度由5‰增加到10‰時,SOD活性呈明顯的增強趨勢,增長率為193.61%。長時淹水環境下,且水體鹽度為5‰~20‰時,SOD活性均明顯增強,然而在30‰鹽度下反而降低。在10‰和20‰鹽脅迫條件下,隨著淹水時間的增加,SOD活性均呈現先增后降再增的趨勢,且在 2 h淹水環境下SOD活性最大。

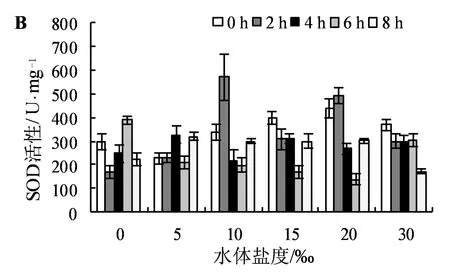
圖2 淹水-鹽脅迫對木欖幼苗葉片POD、SOD活性的影響
由表3可以看出,淹水時間和水體鹽度分別對POD活性和SOD活性產生極顯著影響(P< 0.01);淹水-鹽度交互作用對POD活性和SOD活性也具有極顯著影響(P< 0.01)。
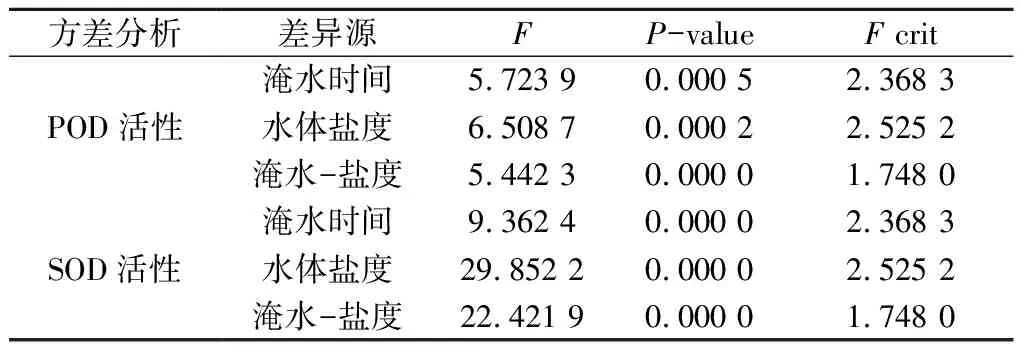
表3 木欖幼苗在淹水-鹽脅迫處理下的POD、SOD活性的方差分析
2.3 淹水-鹽脅迫對木欖幼苗葉片滲透調節物質的影響
如圖3A所示,當水體鹽度<15‰時,可溶性總糖含量隨著淹水時間的增加呈現先降低后增加的趨勢。只有鹽脅迫下,可溶性總糖含量與水體鹽度呈現正相關關系。當木欖幼苗受到鹽脅迫時,短時淹水脅迫下的可溶性總糖含量普遍低于長時淹水脅迫,隨著淹水時間的增加,可溶性總糖不斷累積,有利于細胞膜的滲透調節。當木欖幼苗受到淹水脅迫的同時,低鹽脅迫條件下的可溶性總糖含量普遍高于高鹽脅迫,然而高鹽脅迫造成可溶性總糖含量的降低,不利于滲透調節。由此可知,鹽脅迫對可溶性總糖含量的影響要高于淹水脅迫。
在相同鹽脅迫條件下,隨著淹水時間的增加,可溶性蛋白含量不斷下降,均低于未淹水處理(即淹水時間為0 h)。而在相同淹水脅迫下,隨著水體鹽度的增加,可溶性蛋白含量變化比較復雜;總體上看,在短時淹水處理下,可溶性蛋白含量隨著水體鹽度的增加先增加后下降,而在長時淹水處理下,鹽度越高其可溶性蛋白含量也越高。


圖3 淹水-鹽脅迫對木欖幼苗葉片可溶性總糖、
如表4所示,鹽脅迫、淹水脅迫及其淹水-鹽脅迫交互作用極顯著地影響木欖幼苗葉片內的可溶性總糖含量和可溶性蛋白質含量(P< 0.01)。
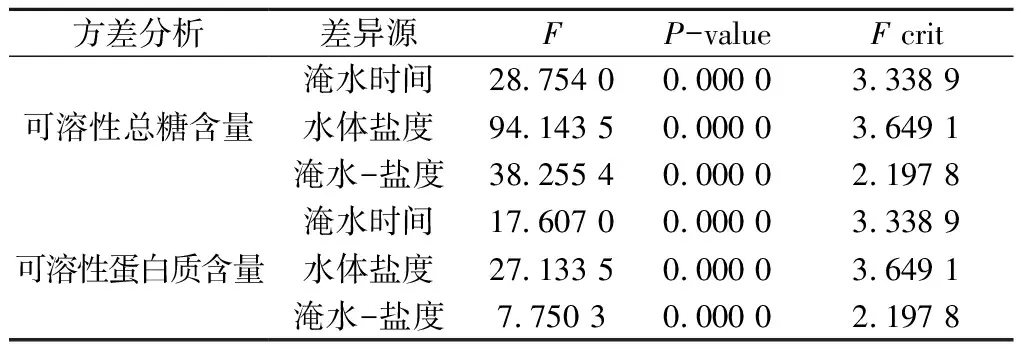
表4 木欖幼苗在淹水-鹽脅迫處理下的可溶性總糖、可溶性蛋白含量的方差分析
3 討論與結論
3.1 淹水-鹽脅迫下木欖幼苗葉片抗氧化作用
在脅迫環境下,植物可通過提高SOD和POD的活性以達到清除過多ROS的目的。高鹽脅迫下秋茄葉片中SOD和POD的活性顯著提高[14]。在水淹脅迫下,秋茄葉片中的POD和SOD活性隨著水淹時間的增加而明顯增強,木欖葉片的POD和SOD活性也顯著提高[15,16]。在淹水-鹽雙脅迫處理下,木欖幼苗葉片POD活性在適宜鹽度(< 15‰)下隨著淹水時間的增加,先降低后提高,而SOD活性均普遍降低;在4 h淹水-30‰鹽脅迫下,木欖幼苗葉片中POD活性最大,而4 h淹水-10‰鹽脅迫下,其SOD活性最大。這些結果說明木欖幼苗葉片內SOD對鹽度變化的敏感性高于POD,且能高鹽度抑制SOD活性,促進POD活性,而低鹽度促進SOD活性。這與前人研究結果不一致,這可能與試驗考慮的脅迫因素和試驗材料的不同有關,具體原因還需后續深入研究。
3.2 淹水-鹽脅迫下木欖幼苗葉片膜脂過氧化作用
水淹、鹽脅迫等逆境脅迫會對植物體內的活性氧(ROS)含量的動態平衡產生不利影響,造成ROS的大量累積,超過了閾值就會導致細胞中膜脂物質受到破壞,MDA含量將會升高。在未淹水條件下,木欖幼苗葉片中MDA含量最大,然而隨著淹水時間的增加,木欖幼苗葉片內MDA含量在總體上呈降低趨勢,這表明隨著淹水脅迫的加劇,木欖幼苗通過積極的抗氧化作用來減輕ROS對植物的傷害,同時淹水環境有利于木欖幼苗的生長。低鹽脅迫下 MDA含量明顯增加,隨著鹽脅迫的加劇,MDA含量顯著降低,這說明木欖幼苗在低鹽脅迫下開始響應鹽脅迫,抗氧化系統還未起作用,隨著鹽度的增加,抗氧化酶發揮作用,有效地降低膜脂過氧化作用,降低逆境對植物造成的傷害[4],這與抗氧化酶活性變化趨勢相一致。
在淹水、鹽度及其交互作用下,木欖幼苗葉片的質膜透性,隨著脅迫的增強,表現出先增加后降低,這與MDA含量的變化趨勢相一致性。木欖幼苗在短時間鹽脅迫下,葉片質膜透性未發生顯著變化,而在長時間鹽脅迫下,葉片質膜透性下降,這表明長時間鹽脅迫下木欖幼苗表現出對鹽脅迫更大的適應性[17]。這也說明膜脂過氧化不是鹽脅迫下木欖葉片膜損傷的主要原因。本試驗中,木欖幼苗因受到低鹽脅迫的傷害而導致質膜透性加強,隨著鹽脅迫的加劇,木欖幼苗通過積極的防護措施降低質膜透性,保證細胞不受到逆境傷害。當淹水-鹽脅迫超出木欖幼苗承受范圍時,質膜透性再次增強。
3.3 淹水-鹽脅迫下木欖幼苗葉片滲透調節作用
在逆境脅迫條件下,植物通過細胞內的滲透調節作用,適應不良環境條件,增強植物抗性[18]。可溶性總糖和可溶性蛋白是參與植物滲透調節作用的主要物質,在環境脅迫下,這兩類物質的大量積累,調節細胞內的滲透勢,維持滲透平衡,穩定蛋白質和細胞膜的正常功能[19,20]。木欖幼苗在鹽脅迫下,隨著水體鹽度的增加,可溶性總糖和可溶性蛋白含量先增加后降低。當水體鹽度從20‰增加到30‰時,可溶性總糖和可溶性蛋白含量均沒有明顯變化,說明高鹽脅迫范圍,細胞內的滲透調節作用不再發生改變。整體上,低鹽脅迫促進可溶性物質的積累,從而抵御不良環境;高鹽脅迫對植物細胞造成一定程度的傷害,從而導致可溶性物質的降低。在淹水環境下,短時淹水使細胞內可溶性總糖含量增加,而長時淹水可促進細胞內可溶性蛋白積累,這說明在淹水脅迫下,木欖幼苗可溶性總糖和可溶性蛋白起到的滲透調節作用并不一致,短時淹水主要是可溶性蛋白發揮作用,長時淹水主要是可溶性總糖在起作用。另外,在淹水-鹽雙脅迫下,可溶性物質的含量均明顯低于單一脅迫,這說明淹水-鹽雙脅迫對植物造成的傷害要高于單一脅迫。這與譚芳林等在淹水-鹽脅迫秋茄的結果相一致。
總體上,木欖幼苗對低鹽脅迫或短時脅迫產生一定的適應性,而對高鹽脅迫或長時脅迫適應性降低。淹水-鹽雙脅迫對木欖幼苗的傷害更嚴重,增加逆境對細胞結構及新陳代謝的破壞作用。通過本試驗分析,我們認為較適合木欖幼苗生長的淹水時間-水體鹽度組合為:2 h-15‰、2 h-20‰、4 h-15‰。但是,在實際種植過程中,除了淹水時間和鹽度對紅樹植物生長會造成影響外,還要多重考慮各類因素的影響,因而本研究結果僅能為木欖造林提供一定的建議。
