DNA條形碼在西藏水系裂腹魚亞科魚類鑒定中的研究
2019-01-22 06:25:26周建設劉海平王萬良曾本和牟振波
淡水漁業 2019年1期
關鍵詞:物種
周建設,張 馳,劉海平,馬 波,王萬良,曾本和,牟振波
(1.西藏自治區農牧科學院水產科學研究所,拉薩 850032;2.中國水產科學研究院黑龍江水產研究所,哈爾濱 150000;3.河南農業大學牧醫工程學院,鄭州 450002)
裂腹魚亞科(Schizothoracinae)隸屬于鯉形目(Cypriniformes)、鯉科(Cyprinidae),全球約有100多種[1],中國共計76個種和亞種,占全球裂腹魚亞科魚類的80%以上[2],廣泛分布于西藏水系,是西藏魚類區系的優勢群體,具有明顯的區域特性[3],曹文宣[4]首次對中國裂腹魚亞科魚類進行了較系統的整理,也為西藏水系裂腹魚的分類奠定了基礎。
DNA條形碼技術自用于物種鑒別體系以來,在魚類的鑒定應用方面越來越多,尤其是在魚卵、仔稚魚等[5]通過外部形態難以辨識的物種上。Ariagna等[6]用DNA條形碼技術分析了古巴126尾淡水魚,認為COI基因序列能夠區分本地的淡水魚,Alba等[7]對亞馬遜河大范圍銷售的“Acará”進行了分析,發現是由7個物種組成,Brito等[8]用DNA條形碼對137個石首魚科進行了鑒定,鑒定率達到90%。在西藏部分及其毗鄰地區以裂腹魚亞科屬間系統發育學、生物地理學和進化中的隔離事件為主線的研究多以線粒體細胞色素b(Cytb)基因為基礎[9],比較系統、全面、客觀地探討了裂腹魚亞科魚類的起源、演化、擴散等與青藏高原的不斷隆升事件的關系,而涉及分類學和物種鑒定的討論并不多,尤其是限定在西藏水系裂腹魚亞科屬間魚種的鑒定鮮有報道,產久林等[10]對采自拉薩市羊卓雍措的60尾5種高原裂腹魚進行了基于COI和16 S rRNA基因部分序列的物種鑒定,表明COI基因能有效鑒定高原裂腹魚魚種。
西藏水系格局復雜,河流縱橫交錯,高原湖泊星羅棋布,裂腹魚亞科魚類作為西藏水系優勢種群,魚類可能存在不同水域遷移和同一水域種間雜交,出現形態相似不同種和同種魚不同形態的現象。近年來,雅魯藏布江魚類資源受到嚴重威脅[11],對其魚類資源的養護迫在眉睫,摸清西藏水系魚類資源現狀,確定優勢物種、普通物種和瀕危物種,制定合理的養護措施,對西藏水系所有魚類進行準確的物種鑒定顯得尤為重要,而傳統的魚類鑒定方法具有一定的局限性[12],歷史上在裂腹魚亞科魚類各屬的界定中也不盡相同[2,13],事實也說明西藏水系現存裂腹魚種類繁多,一部分種間形態學差異不明顯,尤其在幼魚期和裸鯉屬內種間表型極其相似。因此,從分子角度對西藏水系裂腹魚亞科魚類的鑒定顯得尤為重要,應用DNA條形碼技術對西藏水系的裂腹魚亞科魚類進行種類鑒定鮮有報道。本研究是基于線粒體細胞色素C氧化酶1(COI)基因序列的DNA條形碼技術,以目前能捕獲的西藏水系裂腹亞科魚類為研究對象,展開DNA條形碼研究,驗證COI基因序列對采自中國西藏水系的裂腹魚亞科魚類樣本進行物種鑒定的適用性,對于西藏水系裂腹魚亞科魚類種類鑒定,摸清西藏地區魚類資源現狀,制定合理的魚類養護進程具有一定的指導意義。
1 材料與方法
1.1 樣品采集
本研究中,所采集到的裂腹魚亞科魚類形態鑒定主要依據樂佩琦等[13]的分類體系及其對各物種的形態描述。在西藏水域共采集到裂腹魚亞科包括5屬20種的260個樣本,其中包括裂腹魚屬(Schizothorax)的107個樣本,尖裸鯉屬(Oxygymnocypris)30個樣本,裸鯉屬(Gymnocypris)32個樣本,葉須魚屬(Ptychobarbus)22個樣本,裸裂尻屬(Schizopygopsis)69個樣本,代表了西藏水域現能采集到的大部分裂腹魚亞科的魚種。本研究中所測物種均采集于2014—2017年間,野外采樣時,取鰭條組織保存在70%的酒精中,帶回實驗室后保存于-70 ℃備用。
1.2 樣品DNA提取
采用傳統氯仿法從鰭條中提取DNA。
1.3 線粒體基因組COI基因片段擴增和測序
COI基因片段PCR擴增的引物為F(5′-TCAACCAACCACAAAGACATTGGCAC-3′),L(5′-TAGACTTCTGGGTGGCCAAAGAATCA-3′)[10]。引物由美因基因公司合成。PCR反應體系為5×FastPfu Buffer 4 μL,2.5 mmol/L dNTPs 2 μL,Forward Primer(5 μmol/L)0.8 μL,Reverse Primer(5 μmol/L)0.8 μL,FastPfu Polymerase 0.4 μL,Template DNA 10 ng,ddH2O補至20 μL。PCR擴增程序為95 ℃,5 min,95 ℃,30 s,55 ℃,30 s,72 ℃,45 s,27個循環,最后延伸72 ℃,10 min。PCR擴增儀為ABI GeneAmpR9700型。PCR產物通過瓊脂糖凝膠(2%)電泳5 V/cm,20 min,凝膠圖片經凝膠成像系統檢測陽性后送美因基因公司進行測序。
1.4 數據處理
分段進行正反鏈雙向測序的數據經CHROMAS軟件比對后,適當進行人工校對,然后用DNAMAN軟件進行分段數據的拼接,所有COI序列利用CLUSTAL X2.1軟件的默認參數設置為基礎進行同源序列的多重比對,并實施人工校對。比對后的序列在NCBI數據庫中進行Blast比對分析,檢索相似性,確定物種分類地位。依據物種COI序列相似度達到99%以上認定為同一種,相似度92%~99%為同一屬,83%~92%為同一科[14]。通過Mega5.0軟件分析序列的堿基組成及其變異位點等統計數據,計算序列Kimura雙參數種內及種間個體間遺傳距離,在物種鑒定中,遺傳距離以0.02作為種內及種間的鑒定標準[15,16],采用鄰接法(Neighbor-Joining method,NJ)構建分析系統發育樹,并通過Bootstrap檢驗聚類樹各分支的置信度,系統發育樹中的斷線脂鯉(Phenacogrammusinterruptus)和墨西哥麗脂魚(Astyanaxmexicanus)為外群[17]。
2 結果
2.1 傳統形態學鑒定
對所采集的西藏水系裂腹魚亞科260個樣本經形態學鑒定(表1),包括裂腹魚屬10個種的拉薩裂腹魚(S.racomawaltoni)31尾,異齒裂腹魚(S.schizothoraxo'connori)38尾,巨須裂腹魚(S.macropogon)31尾,長絲裂腹魚(S.schizothoraxdolichonema)、瀾滄江裂腹魚(S.racomalantsangensis)、怒江裂腹魚(S.schizothoraxnukiangensis)、西藏裂腹魚(S.labiatus)、墨脫裂腹魚(S.molesworthi)、橫口裂腹魚(S.plagiostomus)、弧唇裂腹魚(S.curvilabiatus)各1尾;尖裸鯉屬1個種的尖裸鯉(O.stewaritii)30尾;裸鯉屬4個種的蘭格胡裸鯉(G.chui)29尾,納木錯裸鯉(G.namensis)、硬刺裸鯉(G.firmispinatus)、色林錯裸鯉(G.selincuoensis)各1尾;葉須魚屬3個種的雙須葉須魚(P.dipogon)20尾,錐吻葉須魚(P.conirostris)、裸腹葉須魚(P.kaznakovi)各1尾;裸裂尻魚屬2個種的拉薩裸裂尻(S.younghusbandi)34尾,高原裸裂尻(S.stoliczkai)35尾。
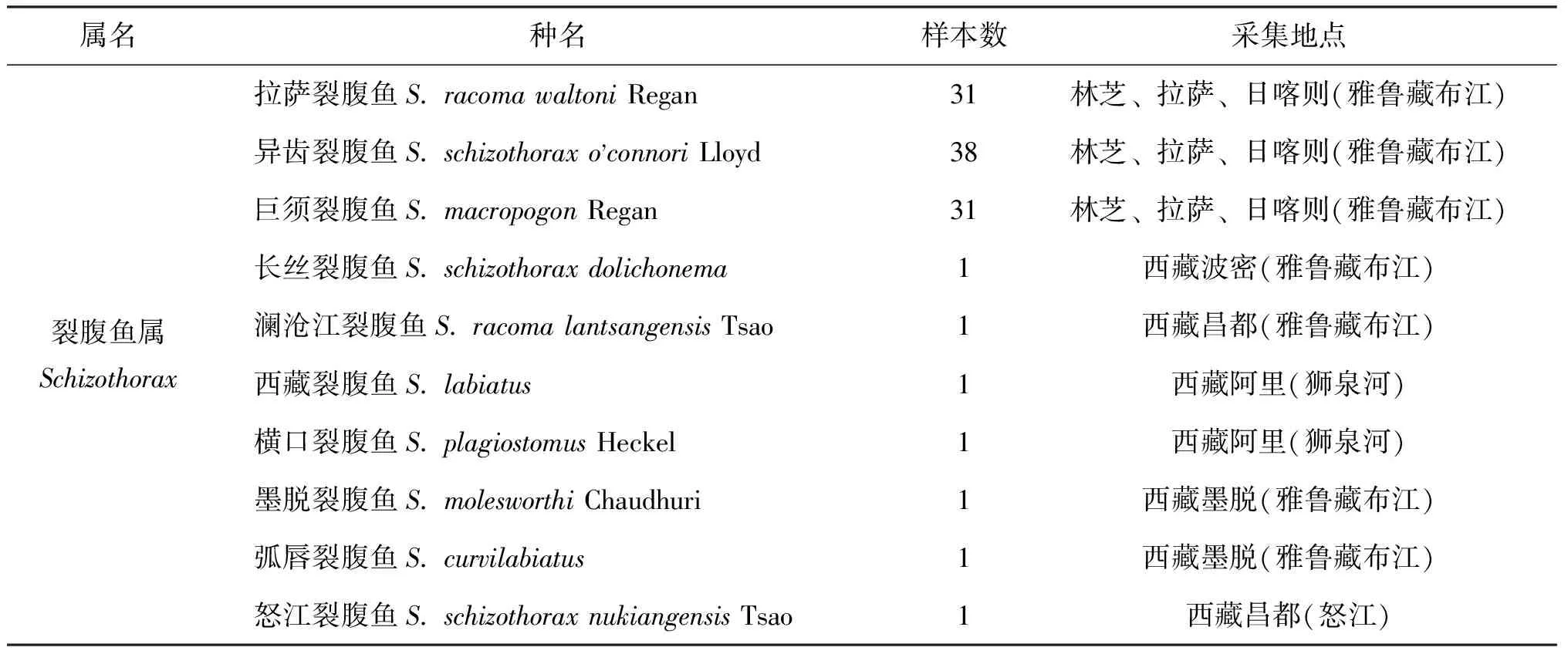
表1 樣本采集信息Tab.1 The collection information of samples
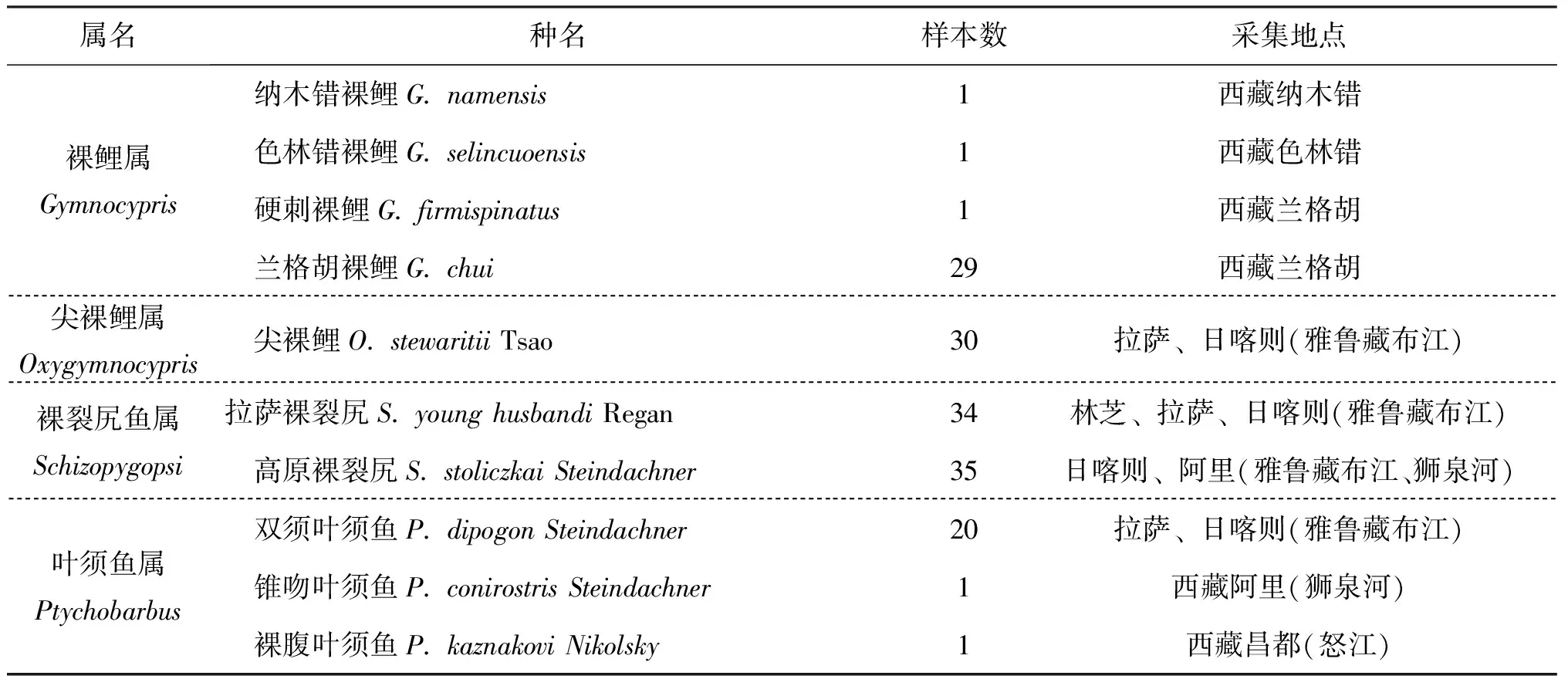
續表
2.2 COI基因序列特征
西藏水域裂腹魚亞科魚類COI基因片段經比對和人工校對后,序列長度為620 bp。包括外群所分析的262條序列620個位點中,有225個可變位點(無插入或缺失),其中161個為簡約性位點,64個為單突變位點,信息位點最多的是第二位密碼子(142個簡約性信息位點),第三位密碼子次之(18個簡約性信息位點),而第一位密碼子最少(1個簡約性信息位點)。序列的堿基組成為A=25.74%、T=29.00%、C=27.44%、G=17.81%,平均轉換/顛換值為8.754。裂腹魚亞科內群260條序列620 bp位點中,有162個可變位點(無插入或缺失),其中143個為簡約性位點,19個為單突變位點,序列的堿基組成為A=25.76%、T=28.98%、C=27.44%、G=17.82%,平均轉換/顛換值為8.899。裂腹魚亞科COI基因片段G+C含量見表2,堿基組成表現出反G偏倚的線粒體基因組的典型特征。
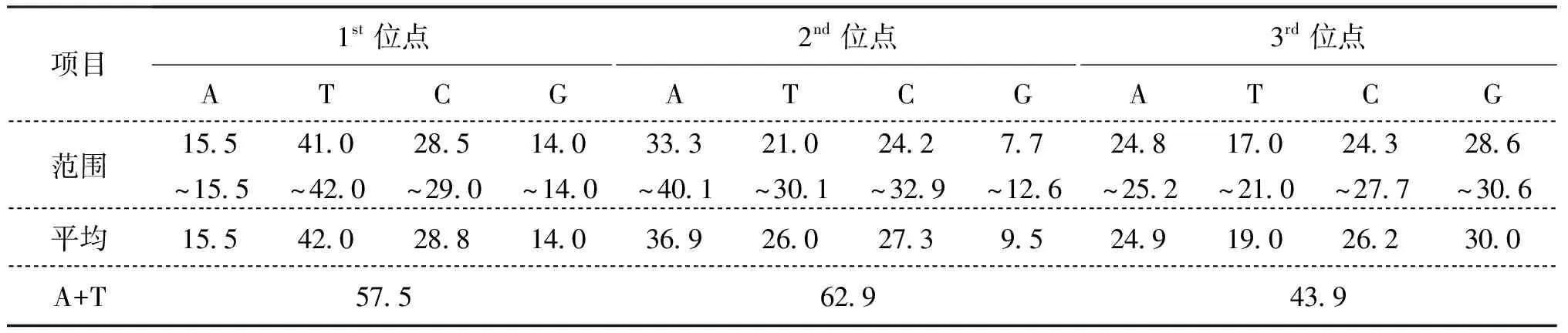
表2 裂腹魚亞科魚類不同密碼子位堿基組成Tab.2 The comprises of base different codon position of Schizothroachinae %
2.3 遺傳距離計算
基于形態學鑒定結果,本研究中構建了西藏水域裂腹魚DNA條形碼數據庫,通過本地數據庫和GenBank雙重比對進行鑒定。計算260個樣本種內及種間遺傳距離,根據種內及種間遺傳距離進行分組,分組原則是種內樣本間遺傳距離小于0.02,種間樣本間遺傳距離大于0.02,根據這一原則,260個個體共分為10組(表3)。對比形態學鑒定結果,Gp 1包含29尾拉薩裂腹魚、38尾異齒裂腹魚、29尾巨須裂腹魚、1尾弧唇裂腹魚和1尾長絲裂腹魚;Gp2包含1尾納木錯裸鯉、1尾硬刺裸鯉、1尾色林錯裸鯉、9尾蘭格湖裸鯉、34尾拉薩裸裂尻、4尾尖裸鯉和2尾拉薩裂腹魚;Gp 3包含35尾高原裸裂尻和20尾蘭格湖裸鯉;Gp 4包含26尾尖裸鯉;Gp5包含20尾雙須葉須魚和3尾巨須裂腹魚;Gp6包含1尾怒江裂腹魚和1尾瀾滄江裂腹魚;Gp7包含1尾西藏裂腹魚和1尾橫口裂腹魚;Gp8包含1尾錐吻葉須魚;Gp9包含1尾裸腹葉須魚;Gp10包含1尾墨脫裂腹魚。10個組中每個種隨機選取個體序列在NCBI中調用Blast程序進行比對以確定其種。
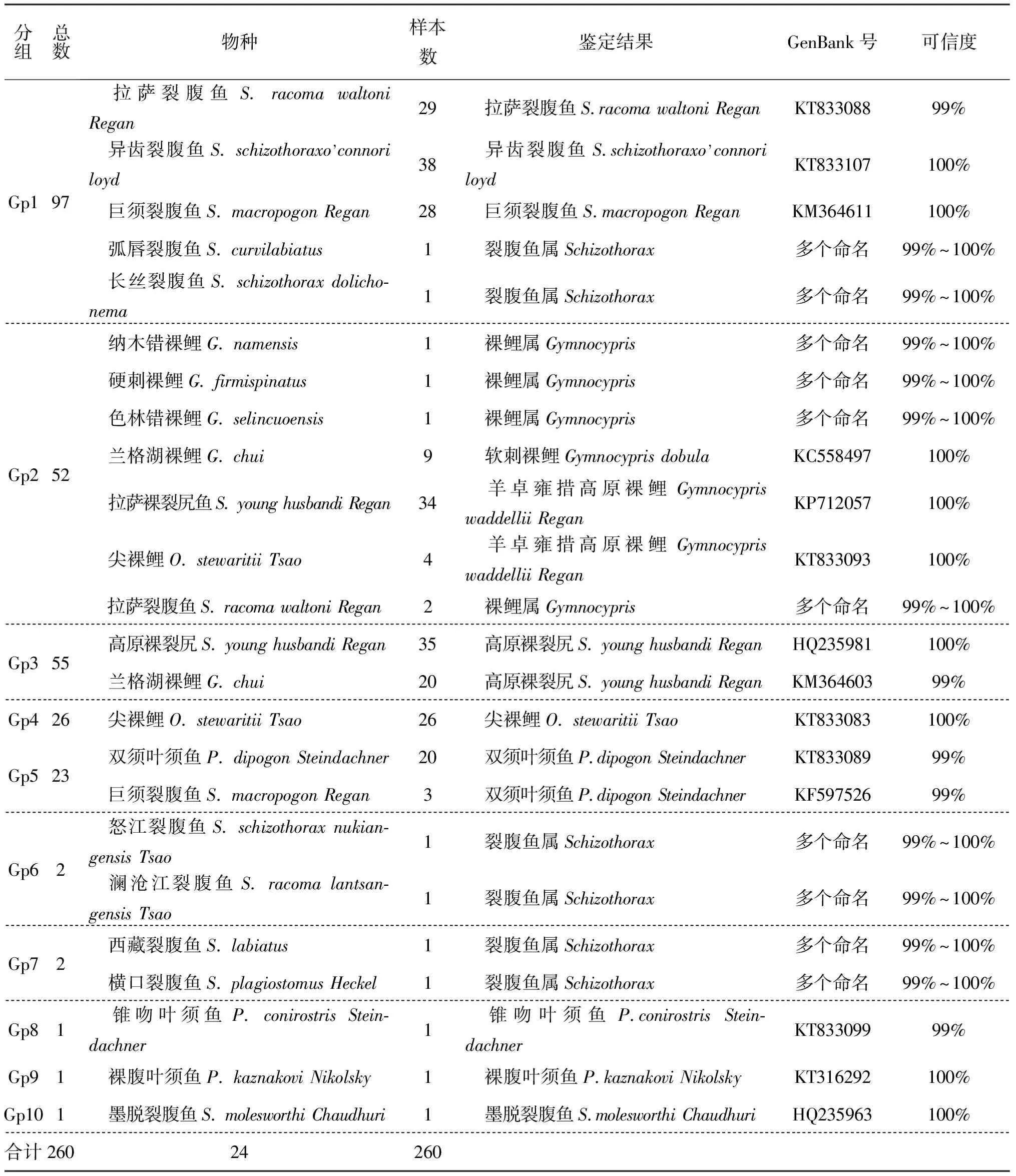
表3 260個樣本根據遺傳距離分組及鑒定結果Tab.3 The grouping and the identification results of 260 individuals based on genetic distance
通過Blast,Gp1中的弧唇裂腹魚和長絲裂腹魚,Gp2中的納木錯裸鯉、硬刺裸鯉、色林錯裸鯉、拉薩裂腹魚,Gp6中的怒江裂腹魚、瀾滄江裂腹魚,Gp7中的西藏裂腹魚、橫口裂腹魚由于數據缺失以及數據庫中存在可信度較高的多個命名,只能鑒定到屬水平。
考慮到各組中包含了由形態學鑒定結果的多個魚種,并且有些魚種在形態特征上差異比較明顯,因此,對各組中個體數量大于1的魚種按形態學鑒定結果再分組,計算組內及組間遺傳距離用于進一步分析(表4)。
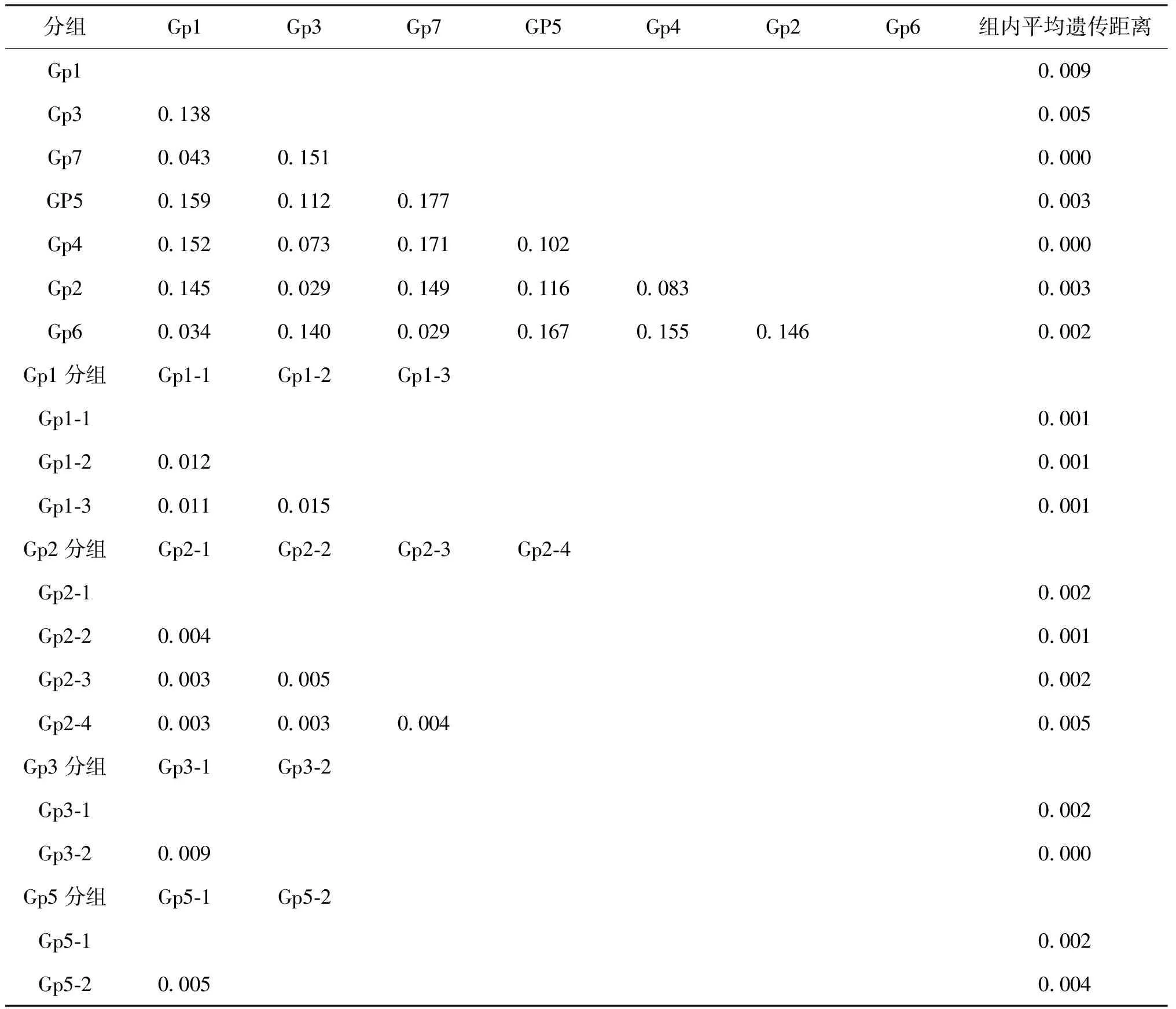
表4 組內及組間平均遺傳距離Tab.4 The average genetic distance of with-in group and inter block
注:對應表3,Gp1-1為異齒裂腹魚;Gp1-2為拉薩裂腹魚;Gp1-3為巨須裂腹魚;Gp2-1為拉薩裸裂尻;Gp2-2為蘭格湖裸鯉;Gp2-3為尖裸鯉;Gp2-4為拉薩裂腹魚;Gp3-1為高原裸裂尻;Gp3-2為蘭格湖裸鯉;Gp5-1 為雙須葉須魚;Gp5-2為巨須裂腹魚。
對各組進一步分析發現,Gp1中Gp1-1(異齒裂腹魚)、Gp1-2(拉薩裂腹魚)和Gp1-3(巨須裂腹魚)之間形態特征具有明顯的差異,再分組發現,組間遺傳距離均大于0.01,組內遺傳距離均小于0.01,其中組間遺傳距離最小的是Gp1-1和Gp1-3,為0.011,組內平均遺傳距離Gp1-1和Gp1-3均為0.001,組間遺傳距離是組內遺傳距離的11倍,Blast結果與再分組鑒定結果一致。Gp2中Gp2-1(拉薩裸裂尻)、 Gp2-2(蘭格湖裸鯉)和Gp2-3(尖裸鯉)之間形態特征差異不明顯,且遺傳距離分析沒有明顯特征,Gp2-4為拉薩裂腹魚,但拉薩裂腹魚與該組中其他魚種形態差異較大,由于樣本時間收集跨度大,查證原始數據確定是樣本分裝時記錄出錯,且Blast結果證實該2個樣本為裸鯉屬,但存在多個命名。Gp2-1、Gp2-3經Blast比對,發現大部分命名為羊卓雍措高原裸鯉,Gp2-2為軟刺裸鯉,均屬裸鯉屬,可信度均為100%。Gp3中Gp3-1(高原裸裂尻)和Gp3-2(蘭格湖裸鯉)遺傳距離分析沒有明顯特征,經Blast兩種均在高原裸裂尻魚。Gp 5中Gp 5-1(雙須葉須魚)和Gp 5-2數據記錄為巨須裂腹魚,但巨須裂腹魚與該組中其他魚種形態差異較大,由于樣本時間收集跨度大,查證原始數據確定是樣本分裝時記錄出錯,Blast結果證實3個巨須裂腹魚為雙須葉須魚,可信度為99%。
2.4 系統進化樹構建
基于表3中的分組,每組中選取各物種代表性序列,總共24條,采用鄰接法(Neighbor-Joining method,NJ)構建系統發育樹,并通過Bootstrap檢驗聚類樹各分支的置信度,系統發育樹中的斷線脂鯉和墨西哥麗脂魚為外群,見圖1。
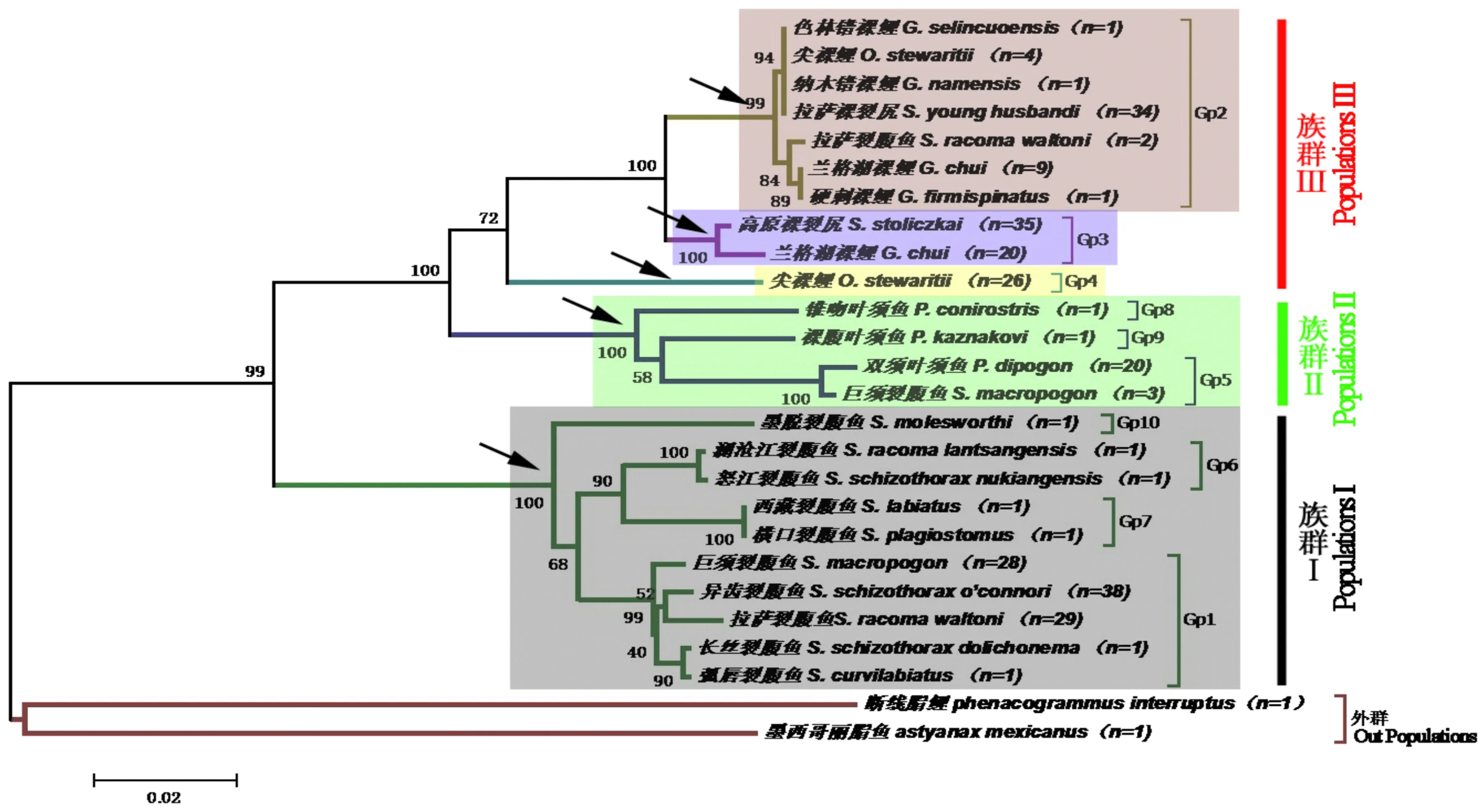
圖1 裂腹魚亞科系統進化樹Fig.1 The phylogenetic tree of Schizothroacinae in Tibet water system
對應表3,圖中不同的顏色區域代表不同的屬,箭頭所示為各個屬分支Bootstrap檢驗支持率。
圖1中,粉紅色區域對應表3中的Gp2組,為裸鯉屬,Bootstrap檢驗支持率為99%;淡藍色區域對應表3中的Gp3組,為裸裂尻魚屬;支持率為100%,淡綠色區域對應表3中的Gp5、Gp8、Gp9組,為葉須魚屬,支持率為100%;淺灰色區域對應表3中的Gp1、Gp6、Gp7、Gp10組,為裂腹魚屬,支持率為100%;淡黃色區域對應表3中的Gp4組,為尖裸鯉屬。斷線脂鯉和墨西哥麗脂魚為外群。裂腹魚亞科魚類形成一個支持率較高的單系群,Bootstrap檢驗支持率為99%。裂腹魚亞科魚類形成三個主要族群,族群I僅包括裂腹魚屬,Bootstrap檢驗支持率為100%,族群II僅包括葉須魚屬,Bootstrap檢驗支持率為100%,族群III包括裸鯉屬、裸裂尻魚屬和尖裸鯉屬,Bootstrap檢驗支持率為72%。系統進化樹聚類方式與表3的分組方式及鑒定結果高度一致。
3 討論
3.1 裂腹魚亞科的形態學鑒定
本研究中的魚類標本是在2014—2017年間所采集,采集到的魚類以形態學鑒定共260尾,分屬為5屬20個種,均為裂腹魚亞科,260尾個體種,249尾以分子生物學手段鑒定到了種水平,鑒出率為95.8%,179尾分子生物學鑒定結果與形態學鑒定結果一致,一致率為68.8%。在野外采樣時,由于條件的限制,不能對所采集魚類進行細致的形態學觀察,尤其是裸鯉屬的魚類大多表觀形態極為相似,不能準確鑒定到種,傳統形態學在對物種的分類過程中耗時耗力,并且有些物種由于環境的差異使同一種類的物種呈現顯著的形態學差異等[18],這都會給物種的鑒定帶來一定的困難。在裂腹魚亞科魚類的分類過程中,也在不斷修正,1962年,曹文宣等澄清了1個屬名和8個種的同物異名[19],1964年又合并了20個同物異名[4],此后,張春霖等[20]、武云飛等[21]、陳毅峰等[5]均對西藏魚類的分類進行了深入的研究,并修正了以往混亂的分類體系。由于物種間形態特征的模糊及分析樣本量的限制,在裂腹魚類個別種屬的界定方面認為有些屬并未形成單系群[11],說明在裂腹魚亞科個別種屬的界定方面仍存在較大爭議,這與本研究中裸鯉屬中種的界定比較混亂是一致的,而且,在形態特征相似物種的鑒定過程中,主觀意志的存在也是不可避免的,本研究中拉薩裸裂尻魚并未與高原裸裂尻歸為一類,而歸為裸鯉屬,可能是形態學鑒定過程中的差異造成,若非形態學鑒定的錯誤,那么很有可能拉薩裸裂尻在分類學中本該歸于裸鯉屬,具體如何界定拉薩裸裂尻,在本研究中還需要補充裸鯉屬其他種類的樣本量。
3.2 DNA條形碼在裂腹魚亞科物種鑒定中的有效性
本研究中對所采集的260尾裂腹魚亞科魚類以組內遺傳距離值小于0.02,組間遺傳距離值大于0.02為原則,進行了分子生物學的鑒定,并與現場采集時形態特征鑒定的結果進行了互相印證。以遺傳距離值為0.02為西藏水系裂腹魚亞科物種鑒定的閾值在裂腹魚屬、裸鯉屬的鑒定中具有一定的局限性。不同物種COI基因進化速率可能是不一致的,因此,使用DNA條形碼進行物種的鑒定中,統一的遺傳距離的閾值很難確定[22,23],本研究中,異齒裂腹魚、拉薩裂腹魚和巨須裂腹魚之間以0.02作為遺傳距離閾值,這3種魚鑒定為一個種,事實上,它們在形態特征上具有比較明顯的差異,而將其遺傳距離閾值設定為0.01,則這3種魚可以有效地得到區分。由于環境的影響,生存在不同水域的同一種魚在形態特征上可能表現出較大的差異,而被認為是不同物種,但這些魚類的基因序列在進化中并未表現出明顯差異,Mohd Shamsudin等[24]研究表明龍魚的體色變化與COI基因分化程度沒有關系,陳信忠等[25]通過檢索BOLD SYSTEMS和GenBank數據庫中慈鯛、神仙魚等觀賞魚的COI基因,檢驗DNA條形碼在觀賞魚物種鑒定中的應用,表明這些觀賞魚由于頻繁雜交,親本的多樣性等,COI基因相似性非常高,遺傳距離小于0.01,使得DNA條形碼難以鑒別這些魚類。本研究中,裸鯉屬種間遺傳距離均小于0.01,最大為0.005,怒江裂腹魚和瀾滄江裂腹魚種間平均遺傳距離為0.002,西藏裂腹魚和橫口裂腹魚種間平均遺傳距離為0.000,弧唇裂腹魚和長絲裂腹魚與異齒裂腹魚之間的遺傳距離均小于0.001,因此,本研究中DNA條形碼在這些魚種間不能被有效區分。由于西藏水系縱橫交錯復雜,這些魚類可能存在物種間的雜交種,或是同一物種在不同生存地理環境當中表現出的形態差異,比如蘭格湖裸鯉和硬刺裸鯉均生活在蘭格湖中,怒江和瀾滄江均屬于長江水系,西藏裂腹魚和橫口裂腹魚均采自雅魯藏布江上游水系,長絲裂腹魚和弧唇裂腹魚均采自雅魯藏布江下游水系等生存環境和水系的影響可能使這些魚類COI基因序列分化程度較低,也有同物異名存在的可能性。由于本研究中樣本收集時間跨度較大,在樣本整理過程中,存在樣本分裝、標記等人為失誤,然而,通過COI基因片段可以將這部分樣本有效區分。本研究中通過Blast程序比對后,裸鯉屬中除羊卓雍措高原裸鯉外,其他均具多個命名,樣本數只有1個的裂腹魚屬也具多個命名,事實上裸鯉屬由于在形態上極為相似,在以往文獻中的分類也是比較混亂的,提示裸鯉屬內種間可能存在同種異名的情況,而樣本數只有1個的裂腹魚屬中的幾個種,在西藏水系中屬非常見種,樣本收集量本來就少,數據庫中記錄也很有限,且數據庫中的數據也不一定是正確的。因此,本研究下一步將繼續應用分子生物學鑒定手段,積累樣本,著重分析上述問題。本研究中所構建的系統進化樹直觀地反映了以遺傳距離閾值為0.02對裂腹魚亞科鑒定分類結果,族群I作為原始族群置于發育樹底部,而族群III以特化群置于發育樹頂部。雖然線粒體DNA在物種鑒定中與形態學鑒定存在不一致性,但是能夠很好地反映水系間的地理和歷史聯系,總體上相近的水系的物種具有更近的親緣關系[26]。本研究中,采自長江水系的怒江裂腹魚和瀾滄江裂腹魚以100%的支持率聚在一支,采自雅魯藏布江上游水系的西藏裂腹魚和橫口裂腹魚以100%的支持率聚在一支,采自雅魯藏布江下游水系的長絲裂腹魚和弧唇裂腹魚以90%的支持率聚在一支。本研究中分析結果及推理仍受限于部分魚種標本單一的事實,無法進行深入分析,這也是下一步需要開展的工作。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
