三級淋巴樣組織在腫瘤微環境中的形成及作用
2019-01-24 09:03:08金向宇
醫學綜述 2019年2期
關鍵詞:環境
李 暉,金向宇,洪 衛
(浙江省腫瘤醫院腫瘤內科,杭州 310022)
免疫細胞招募和聚集是實體腫瘤微環境的重要特征,并隨著腫瘤類型、分期及個體情況的差異呈現出現廣泛異質性[1]。免疫細胞在腫瘤中的作用也隨著免疫檢查點抑制劑的臨床應用而日益受到愈來愈多的關注。免疫細胞一方面具備針對腫瘤的特異性免疫應答進而控制腫瘤的作用,另一方面腫瘤微環境中大量炎性細胞的浸潤也是造成腫瘤微環境中免疫抑制的重要原因[2-5]。針對免疫細胞在腫瘤患者的分期、預后及臨床治療效果中的作用已經進行了較為深入的研究,同時通過恢復免疫細胞的活化進而控制腫瘤的設想也從理論上進入了臨床實踐[6]。三級淋巴樣組織(tertiary lymphoid tissue,TLS)是慢性感染、腫瘤中出現的具備次級淋巴樣組織類似結構并具有啟動免疫應答能力的新生免疫結構[7-8]。該結構的形成對于塑造腫瘤微環境中的免疫表型,從而決定腫瘤生物學行為具有重要意義。現就TLS的形成及機制進行綜述,分析該結構在腫瘤微環境免疫應答中的作用,進而概述總結其在臨床實踐中的應用價值。
1 腫瘤微環境
1.1腫瘤微環境的異質性 腫瘤微環境是由多種細胞包括腫瘤細胞、免疫細胞、內皮細胞、成纖維細胞及基質等相互作用形成的網絡性的微生態系統[5]。其中,免疫細胞是免疫微環境極為重要的部分,腫瘤微環境在很大程度上反映了機體免疫與腫瘤細胞相互博弈的動態過程,因此,深入分析腫瘤微環境中免疫細胞浸潤的位置、密度及類型對于明確腫瘤免疫的狀態是極為關鍵而有效的參數。隨著免疫檢測點抑制劑的臨床應用,免疫微環境中浸潤的免疫細胞得到越來越多的關注,其在腫瘤不同生物學進程中多樣性的功能備受關注。總之,腫瘤浸潤免疫細胞同時扮演完全相反的角色:一方面淋巴細胞通過適應性免疫應答發揮免疫監視功能;另一方面免疫細胞產生的炎性環境提供了腫瘤發生、發展及轉移的重要環境基礎[3-4]。因此,在腫瘤微環境中既有發揮抗腫瘤作用的活化的T淋巴細胞,又存在M2型巨噬細胞、髓系來源抑制細胞及調節性T細胞等免疫抑制細胞[9-11]。這種看似自相矛盾的情況卻在同時存在于腫瘤微環境中,而兩部分的平衡則因腫瘤部位及分期的不同存在較大差異。基于此,旨在恢復免疫細胞殺傷腫瘤能力或逆轉抑制性的免疫環境的免疫治療策略以細胞毒T淋巴細胞相關抗原4及程序性細胞死亡受體1/程序性細胞死亡配體1單抗在臨床的應用得以實踐。但如何更全面且準確地把握腫瘤發生不同階段介導免疫細胞抗腫瘤的具體因素,并將其應用到臨床實踐中仍處于起始階段。腫瘤細胞的異質性對于腫瘤微環境中浸潤的免疫細胞數量、種類、位置及預后有重要意義。利用此特點通過檢測免疫細胞浸潤情況評估腫瘤的生物學特點及臨床病理特征是目前研究的熱點。一般認為,腫瘤微環境中浸潤T淋巴細胞往往與術后患者的預后密切相關[12],另有學者認為采用免疫細胞浸潤進行分期對于臨床預后的預測較TNM分期未見明顯差異[13]。
1.2腫瘤免疫循環 隨著臨床及轉化性研究數據的積累,Chen和Mellman[14]提出了腫瘤免疫循環理論,該理論全面地論述了以T細胞為代表的適應性免疫細胞實現腫瘤細胞殺傷的7個關鍵步驟:①腫瘤抗原的釋放;②腫瘤抗原的呈遞;③T細胞預刺激及活化;④T細胞遷移至腫瘤部位;⑤T細胞浸潤至腫瘤內部;⑥腫瘤細胞被T細胞識別;⑦T細胞殺傷腫瘤細胞。其中,T細胞的活化建立在適當的抗原釋放,正確的抗原呈遞以及合適的預刺激基礎上,而T細胞活化一般發生在次級淋巴組織中,但在腫瘤中往往缺乏特異性的次級淋巴組織。而有多位學者觀察到在腫瘤免疫及炎癥免疫的微環境中存在一種類似淋巴結的TLS結構[15-16]。TLS具備一個完整淋巴結的結構,包括包含成熟樹突狀細胞(dendritic cells,DC)的T細胞區域;包含濾泡DC、增殖的B細胞及高內皮靜脈(high endothelial venule,HEV)的生發中心區域[17-18],并能夠成功而有效地觸發適應性免疫應答[19-20]。鑒于TLS在誘發適應性免疫應答中的重要作用,很多研究發現其與腫瘤的臨床預后密切相關[9,16,21-24]。
2 TLS的形成與腫瘤微環境
2.1次級淋巴結的形成 在胚胎發育時期,次級淋巴結的形成依賴于造血前體誘導細胞與基質形成細胞的相互作用[25]。而這種相互作用主要是由表達于基質形成細胞表面的淋巴毒素β受體及表達于前體誘導細胞表面的相應配體介導的,兩者結合后可誘導基質形成細胞表面表達特異性的黏附分子,包括血管細胞黏附分子1、細胞間黏附分子(intercellular adhesion molecule,ICAM)-1、黏膜黏附分子1以及淋巴形成趨化因子,包括CC趨化因子配體[chemokine (C-C motif) ligand,CCL]19、CCL21及CXC趨化因子配體[chemokine (C-X-C motif) ligand,CXCL]13,進而招募T細胞及B細胞組織性的聚集。TLS的形成與次級淋巴結的形成過程類似[26]。受到來自腫瘤微環境導致淋巴樣結構形成的各種信號刺激,基質細胞活化后亦可通過表達多種黏附分子啟動TLS的形成,而腫瘤微環境中的各種炎性因子則可促進這一進程的發生。臨床前研究發現組織特異性過表達淋巴毒素α或CCL21和CXCL13可誘導TLS的形成[8,27-29],而這些基因的缺失造成多種感染及自身免疫性疾病動物模型的病理部位中TLS的形成障礙[28,30]。
2.2腫瘤微環境中TLS的形成 無論是免疫細胞聚集還是細胞代謝出入,都需要血管和淋巴管提供暢通的道路,因此,在TLS形成過程中脈管系統的構建是極為關鍵的步驟。首先,基質細胞表面的淋巴毒素β受體激活后可誘導下游的淋巴血管形成相關基因表達,如血管內皮生長因子C、血管內皮生長因子D、成纖維細胞生長因子2等,從而進一步誘導淋巴管內皮透明質酸受體1陽性淋巴管的形成[31-32]。其次,血管及淋巴管內皮細胞與周圍免疫細胞(如DC)之間的相互作用對組織內脈管系統的維續也較為重要。以目前研究較為清楚的HEV為例,在多種實體瘤組織中,HEV的密度與患者的臨床預后明顯相關,還對T細胞的招募有正性調控作用[26]。在偶氮甲烷/硫酸葡聚糖誘導的腸癌模型中,腫瘤組織的TLS明顯多于正常組織,而其中T細胞的浸潤與CD31的表達及外周淋巴結地址素(peripheral node addressin,PNAd)陽性HEV明顯相關。尾靜脈注射綠色熒光蛋白標記的脾淋巴細胞更多地在腫瘤的TLS中聚集,而在正常組織中并未發現,也進一步證實TLS在招募免疫細胞中的重要作用[24]。
腫瘤微環境中的TLS最早發現于肺癌及黑色素瘤。Ladanyi等[33]在黑色素瘤原位腫瘤術后標本中發現TLS中溶酶體相關膜糖蛋白(lysosome-associated membrane glycoprotein,LAMP)陽性的成熟DC與腫瘤微環境中活化T細胞的浸潤以及患者的無病生存期呈正相關。而在非小細胞肺癌中,DC-LAMP+成熟DC選擇性的聚集于T細胞浸潤區域,并且與生發中心區域(包含Ki-67陽性B細胞、濾泡T輔助細胞、CD68+巨噬細胞以及CD21+CD23+濾泡CD)鄰近[34]。腫瘤微環境中的TLS被特異性的PNAd陽性血管和淋巴管包圍,脈管內皮細胞表面還表達多種黏附分子(如ICAM-2、ICAM-3、血管細胞黏附分子1)及黏膜地址素細胞黏附分子1,從而形成典型的HEV系統[34]。而在腫瘤微環境中HEV往往與TLS共存,也從另一個側面說明HEV對于TLS形成的關鍵作用。進一步研究發現,TLS浸潤的淋巴細胞表面大部分表達PNAd的CD62配體,而TLS中則組成性表達一系列招募免疫細胞的黏附分子及趨化因子,如黏附分子(ICAM-2、ICAM-3、血管細胞黏附分子1、黏膜黏附分子1)、整合素(αL、α4、αD)、趨化因子(CCL19、CCL21、CXCL13)、CC趨化因子受體4配體(CCL17、CCL22)[15]。
目前無法嚴格的區分是典型的TLS還是簡單的淋巴細胞聚集,已有大量研究報道了類似于TLS的結構在多種人類腫瘤組織中發現,如結直腸癌[24]、乳腺癌[21,23]、黑色素瘤[23,35]、黏膜相關淋巴組織淋巴瘤[36]以及非小細胞肺癌[15,23]。進一步的研究發現[16],無論是在早期(接受新輔助化療)還是轉移性階段的腫瘤中均發現TLS存在,說明TLS的產生是腫瘤中較為普遍的現象。而在TLS結構方面也存在較大異質性,有研究發現在黑色素瘤的轉移病灶中,淋巴樣組織區域中T/B細胞、HEV、濾泡DC及DC的聚集并不完全相同,多種細胞可以不同組合進行聚集,從而形成不同成熟程度的淋巴樣組織[35]。不僅如此,在不同的原發腫瘤中存在的淋巴樣組織亦差別較大。有學者分析了多種腫瘤的肺轉移灶中淋巴樣組織的差別,發現來源于肺癌、前列腺以及結腸癌的肺轉移灶較乳腺癌及腎癌的肺轉移灶具有更高密度的TLS[37]。
3 TLS與抗腫瘤免疫應答
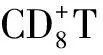
4 TLS與臨床預后
5 結 語
TLS作為腫瘤微環境中的重要組成部分,在臨床預后中的價值已得到學術界的公認,而使用不同的TLS標志物并沒有改變對臨床預后的價值。因此,TLS作為腫瘤局部形成的淋巴結類似結構對于腫瘤微環境中淋巴細胞的招募、活化和增殖均具有無可替代的作用,這也提示通過恰當的臨床治療策略如腫瘤疫苗、免疫檢測點抑制劑等增加腫瘤微環境中TLS的密度,對于塑造正性的抗腫瘤免疫應答,從而改善患者的無病生存期、無進展生存期甚至總生存期均是未來的研究方向。但TLS產生的具體機制,如何調控腫瘤免疫應答,如何與腫瘤局部溝通、交流等一系列問題尚無結論,仍需大量的基礎研究和臨床轉化性研究深入挖掘。相信隨著對整個腫瘤免疫更清晰的認知,TLS將會在已經到來的腫瘤免疫治療時代發揮重要作用。
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學院學報(2017年2期)2017-04-20 08:44:31
中國環境監察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38
