樟絨枝霉α-淀粉酶在畢赤酵母中的高效表達及在麥芽糖漿制備中的作用
2019-02-15 02:56:22趙寧王玉川易萍閆巧娟江正強
食品與發酵工業 2019年2期
趙寧,王玉川,易萍,閆巧娟,江正強*
1(中國農業大學 食品科學與營養工程學院,北京,100083)2(中國農業大學 工學院,北京,100083)
α-淀粉酶廣泛應用于食品、飼料、造紙等工業中,在整個酶制劑市場中約占25%份額[1]。微生物是α-淀粉酶的主要來源,其中產α-淀粉酶的細菌主要是芽孢桿菌屬微生物[2],真菌主要是曲霉和青霉屬[3],此外還有一些嗜熱真菌產α-淀粉酶,如Scytalidiumthermophilum[4]和Thermomyceslanuginosus[5]。細菌α-淀粉酶的最適溫度和最適pH分布廣泛,真菌α-淀粉酶的作用條件較溫和,最適溫度一般在50~60 ℃,最適pH一般在5.0~6.0[3]。HAN等[6]研究了嗜熱真菌樟絨枝霉(Malbrancheacinnamomea)產α-淀粉酶的產酶條件、純化及性質,但產酶水平低,無法工業生產應用。
構建工程菌株是實現外源基因高效表達的有效途徑,巴斯德畢赤酵母作為較完善的真核表達系統,目前,已有多種來源α-淀粉酶在畢赤酵母中高效表達。WANG等[7]將地衣芽孢桿菌(Bacilluslicheniformis)α-淀粉酶在畢赤酵母高效表達,經5 L發酵罐分批補料發酵168 h,α-淀粉酶產量為8.3 mg/mL,在 50 L發酵罐中α-淀粉酶產量為12.2 mg/mL;PARASHAR等[8]克隆來源于Bacillusacidicola和Geobacillusthermoleovorans嵌合的α-淀粉酶基因,成功表達于畢赤酵母,在7 L發酵罐中發酵,最終α-淀粉酶表達量為0.120 mg/mL,比酶活為1 180 U/mg(DNS法,每分鐘反應產生1 μmol麥芽糖定義為1個酶活單位)。可以看出α-淀粉酶在畢赤酵母中高效表達具有較大應用潛力,但是目前關于嗜熱真菌α-淀粉酶在畢赤酵母中表達的報道較少,僅有Rhizomucorpusillus來源α-淀粉酶。ZHANG等[9]將Rhizomucorpusillus來源α-淀粉酶和糖化酶在畢赤酵母共表達,與單一表達糖化酶相比,共表達淀粉酶的糖化活力提高了79%。
在麥芽糖漿的工業生產中,淀粉需經液化和糖化兩個步驟,液化主要采用耐高溫α-淀粉酶,糖化采用β-淀粉酶、真菌α-淀粉酶等糖化酶。真菌α-淀粉酶作為生產麥芽糖漿的關鍵酶之一,已廣泛應運于生產,如以大米淀粉為原料,經真菌α-淀粉酶水解制備得麥芽糖含量為45.18%的麥芽糖漿[10]。為制備高麥芽糖漿,常將α-淀粉酶與普魯蘭酶、β-淀粉酶等復配,亢潘潘[11]等將真菌α-淀粉酶與β-淀粉酶復配,制備得到麥芽糖含量為62.1%的麥芽糖漿。此外還采用色譜分離、酵母發酵等方法提高麥芽糖含量,黃偉紅[12]等將色譜分離后的麥芽糖漿經活性干酵母發酵,制備得到麥芽糖含量為99%的超高麥芽糖漿。本文將嗜熱真菌樟絨枝霉α-淀粉酶基因McAmyA在畢赤酵母GS115中高效表達和高密度發酵,研究重組α-淀粉酶McAmyA的酶學性質,并直接以工業化生產的液化產物為底物,研究該酶在制備麥芽糖漿中的應用。
1 材料和方法
1.1 材料
1.1.1 試驗材料
菌株:樟絨枝霉S168由本實驗室保存,大腸桿菌E.coliDH5α,購于北京博邁德生物技術有限公司,畢赤酵母GS115和質粒pPIC9K,購于美國Invitrogen公司,淀粉液化液由山東隆科特酶制劑有限公司提供(玉米淀粉為底物,細菌α-淀粉酶高溫液化制得)。
液化液原液:干物質含量32%,DE值32,葡萄糖、麥芽糖、麥芽三糖、麥芽四糖的含量分別為(以干物質計):1.5%、11.8%、14.1%、8.7%,殘留液化酶活為150 U/mL。
1.1.2培養基
LB、YPD、MD、BMGY、BMMY等培養基的配制方法參照Invitrogen公司的《畢赤酵母表達操作手冊》。
PTM1(g /L):CuSO4·5H2O 6,NaI 0.08,MnSO4·H2O 3,Na2MoO4·2H2O 0.2,H3BO30.02,CoCl20.5,FeSO4·7 H2O 65,生物素 0.2,濃硫酸 5 mL。
1.2 方法
1.2.1 樟絨枝霉α-淀粉酶基因McAmyA的克隆和表達載體構建
根據HAN等[6]的報道,以cDNA為模板,設計引物McAmyADNAF(5′—ATGGTTTCAACAGCACTATTCCT—3′)和McAmyADNAR(5′—TCAGTTCTCGCAAAGCCCGGA—3′),克隆樟絨枝霉α-淀粉酶基因McAmyA,然后將PCR擴增回收的DNA片段和巴斯德畢赤酵母表達載體pPIC9K雙酶切,酶切位點為SnaBI和AvrⅡ,用試劑盒回收DNA片段和表達載體pPIC9K,T4連接酶連接目的基因片段和表達載體后,轉化至大腸桿菌DH5α,涂布于含氨芐青霉素的LB抗性平板培養基,挑取轉化子進行菌落PCR和雙酶切驗證,陽性轉化子命名為pPIC9K-McAmyA。
1.2.2 α-淀粉酶基因McAmyA在畢赤酵母中表達
提取陽性轉化子質粒,進行線性化,電轉化至畢赤酵母感受態GS115,涂布于MD平板培養基,30 ℃培養4 d,待轉化子長出后,將其涂布在含不同濃度遺傳霉素G418的平板培養基。長出酵母后,挑取單菌落到BMGY培養基,30 ℃恒溫搖床培養至OD600=2~6,離心收集菌體,用BMMY培養基重懸并接種至BMMY培養基,使OD600達1.0,30 ℃恒溫搖床誘導3 d,每24 h補充甲醇,使甲醇終體積分數為0.5%。
1.2.3 畢赤酵母高密度發酵
選取搖瓶發酵酶活最高的菌株,在5 L發酵罐進行高密度發酵,工作體積為1.5 L,發酵方法和發酵培養基的配制參照PichiaFermentation Guidelines (Version B,053002,Invitrogen)。發酵過程主要分為分批培養、甘油流加培養和甲醇誘導培養3個階段。
分批培養:裝1.5 L基本培養基,滅菌發酵罐,28%濃氨水調節pH至4.0,加入PTM14.35 mL/L,接種量10%,起始轉速600 r/min,溫度30 ℃,發酵18~24 h。
甘油流加培養:待分批培養至培養基中的甘油耗盡,流加50%甘油,28%濃氨水調節pH至5.0,流速為18.4 mL/(h·L),流加時間4~5 h,待OD600達180~220,停止流加甘油。
甲醇誘導培養:停止流加甘油后,饑餓30 min左右,28%濃氨水調節pH至6.0,轉速調為800 r/min,流加甲醇,誘導產酶,監控溶氧變化,保持DO>20%,發酵過程中取樣分析菌體濕重、酶活力和蛋白含量。
1.2.4 α-淀粉酶活力及蛋白含量的測定
α-淀粉酶活力測定:采用DNS法[13]測定酶活力,將900 μL用緩沖液配置的10 g/L可溶性淀粉,于一定溫度水浴鍋中預熱5 min后加入100 μL適當稀釋的酶液,在一定溫度下,水浴反應10 min,加入1 mL DNS終止反應,沸水浴10 min后測吸光值OD540,以葡萄糖作為標準品。酶活定義:每分鐘反應生成1 μmol葡萄糖所需要的酶量定義為1個酶活力單位。
另外,同時采用輕工行業標準(QB 2526—2001)測定酶活,將10 mL可溶性淀粉溶液在40 ℃水浴鍋中預熱10 min,加入適當稀釋的酶液1 mL,40 ℃準確加熱30 min后,加入費林試劑4 mL使酶失活,硫代硫酸鈉滴定測定生成的還原糖。酶活定義:反應30 min,反應液中產生相當于10 mg葡萄糖的還原糖所需的酶量,定義為1個淀粉糖化酶活力單位。
蛋白含量測定:參照LOWRY等的方法[14],以牛血清蛋白(BSA)作為標準蛋白。
1.2.5 重組α-淀粉酶McAmyA的純化
重組α-淀粉酶McAmyA經甲醇誘導168 h后,10 000 r/min 離心5 min收集上清液,用50 mmol/L Tris-HCl (pH 8.0)在4 ℃條件下透析過夜,然后10 000 r/min離心10 min,收集上清液即為粗酶液。將粗酶液過QSFF強陰離子交換柱(50 mmol/L pH 8.0 Tris-HCl緩沖液平衡),流速為1.0 mL/min,用0~500 mmol/L 的NaCl溶液洗脫,收集有α-淀粉酶活性的組分,采用SDS-PAGE分析蛋白純度。
1.2.6 最適pH和最適溫度測定
最適pH的測定:選擇不同pH的50 mmol/L 6種緩沖液,包括檸檬酸-檸檬酸鈉(pH3.0~6.0)、醋酸-醋酸鈉(pH4.0~6.0)、MES(pH5.5~7.0)、MOPS(pH6.5~8.0)、CHES(pH8.0~10.0)和CAPS(pH10.0~11.0),配制1%可溶性淀粉底物,在65 ℃條件下,按照DNS法,測定重組α-淀粉酶McAmyA在不同緩沖體系中的酶活,測定酶活最高點設為100%。
最適溫度的測定:用最適pH緩沖液配制1%可溶性淀粉底物,在40~90 ℃條件下,按照DNS法測定重組α-淀粉酶McAmyA酶活,測定酶活最高點設為100%。
1.2.7 重組α-淀粉酶McAmyA水解液化液制備麥芽糖漿
1.2.7.1 重組α-淀粉酶McAmyA加酶量對麥芽糖含量的影響
以液化液為底物,在60 ℃和自然pH(pH 5.9)下,按照液化液質量分別添加α-淀粉酶40、60、80、100、120、140、160、180 U/g,水解8 h,HPLC檢測麥芽糖含量,考察重組α-淀粉酶McAmyA加酶量對麥芽糖含量的影響,并計算在最優加酶量下的水解率。
1.2.7.2 水解溫度對麥芽糖含量的影響
以液化液為底物,按照120 U/g的加酶量,自然pH(pH 5.9),水解溫度分別為50、55、60、65、70、75 ℃下水解8 h,HPLC檢測麥芽糖含量,考察水解溫度對麥芽糖含量的影響,并計算在最佳水解溫度下的水解率。
1.2.7.3 水解時間對麥芽糖含量的影響
以液化液為底物,在加酶量為120 U/g、溫度60 ℃和自然pH(pH 5.9)的條件下分別水解8、10、12、14、16、20、24 h,HPLC檢測麥芽糖含量,考察水解時間對麥芽糖含量的影響,并計算在最佳水解時間下的水解率。
1.2.7.4 糖含量測定和水解率計算
糖含量采用高效液相(HPLC)測定。
色譜柱:氨基鍵合柱(4.6×250 mm,5 μm);檢測器:ELSD 2424 蒸發光檢測器;柱溫:45 ℃;流動相:乙腈∶水=60∶ 40,流速1 mL/min,進樣體積10 μL,糖化液過0.22 μm濾膜,稀釋一定倍數備用。
水解液中各糖含量和水解率的計算如下:

(1)

(2)
干物質質量測定:阿貝折光儀直接測定(GB/T 20885—2007)。
2 結果與分析
2.1 α-淀粉酶基因McAmyA的畢赤酵母表達和高密度發酵
α-淀粉酶基因McAmyA在畢赤酵母GS115中成功表達,高拷貝篩選酶活最高的菌株進行5 L發酵罐高密度發酵,測定發酵液的酶活、蛋白含量和菌體濕重,并對發酵上清液進行SDS-PAGE分析,結果如圖1所示。
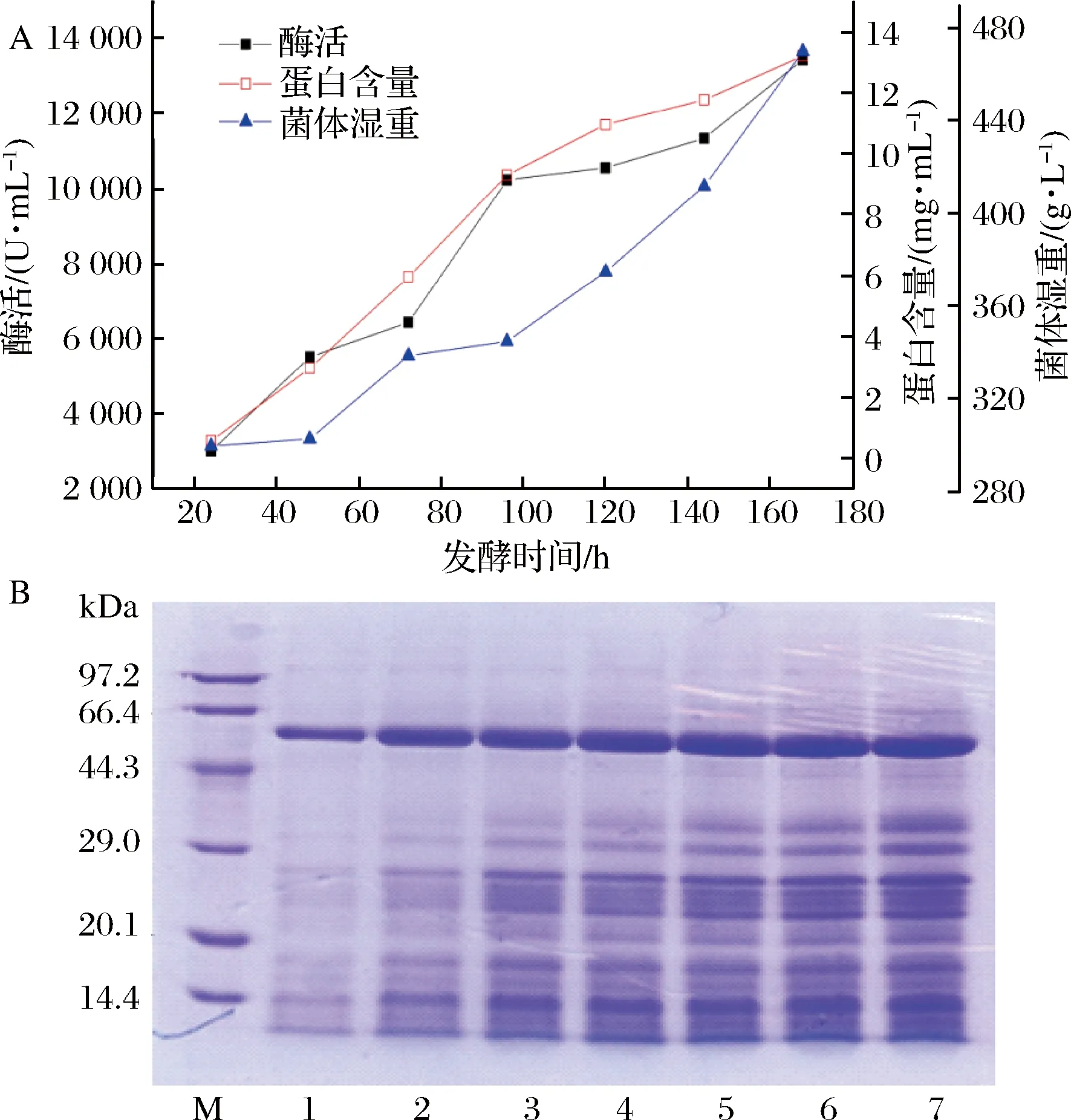
M-低分子質量標準蛋白;1-7分別為甲醇誘導24、48、72、96、118、144、168 h的發酵上清液圖1 畢赤酵母表達重組α-淀粉酶McAmyA的高密度發酵歷程(A)及SDS-PAGE(B)Fig.1 Time-course of recombinantα-amylase (McAmyA) expression in Pichia pastoris by high cell density fermention (A) and SDS-PAGE analysis of curde enzyme (B)
隨甲醇誘導時間增加,菌體濕重、蛋白含量和α-淀粉酶酶活力逐漸上升(圖1-A),由SDS-PAGE可看出,目標蛋白條帶隨發酵時間增加逐漸變粗(圖1-B)。甲醇誘導168 h時,菌體濕重為470 g/L,胞外酶活力達13 440 U/mL(QB 2526—2001法測定酶活為:253.6 U/mL),蛋白含量為13.2 mg/mL,酶活力相比最初搖瓶發酵(554 U/mL),提高了24.3倍。
野生型樟絨枝霉菌株產α-淀粉酶活力低(308.5 U/mL),α-淀粉酶基因McAmyA在畢赤酵母GS115中成功表達,進行高密度發酵后,相對于原始菌株發酵,酶活提高了43.6 倍。已報道畢赤酵母表達中α-淀粉酶的表達水平較低。米曲霉(Rhizopusoryzae)α-淀粉酶在畢赤酵母中表達,分批補料發酵140 h后,蛋白表達量為0.382 mg/mL[15]。地衣芽孢桿菌(Bacilluslicheniformis)α-淀粉酶在畢赤酵母中表達,經5 L發酵罐發酵168 h,蛋白表達量為8.3 mg/mL(酶活8 100 U/mL,DNS法,每分鐘水解1 mg淀粉定義為1個酶活單位)[7]。本研究中樟絨枝霉α-淀粉酶在畢赤酵母GS115中高效表達,表達量在α-淀粉酶研究中處于較高水平。
2.2 重組α-淀粉酶McAmyA的純化
重組α-淀粉酶McAmyA的純化表如表1所示。

表1 重組α-淀粉酶McAmyA純化表Table 1 Purification summary of McAmyA
注:a酶活測定條件:可溶性淀粉為底物,MES緩沖液(pH 6.5),65 ℃;b蛋白質測定采用Lowry法,以牛血清蛋白(BSA)為標準品。
發酵粗酶液經QSFF強陰離子交換柱后一步純化得到電泳級純酶,純化倍數為1.2,比酶活為1 230.2 U/mg,酶活力回收率為63.5%,純酶在SDS-PAGE上顯示為單一蛋白條帶(見圖2),分子質量約為55 kDa。基因序列預測該蛋白分子質量為51.9 kDa,含有1個可能的N-糖基化位點,純酶經Endo H處理后,分子質量沒有發生明顯變化,說明該蛋白可能未發生糖基化。
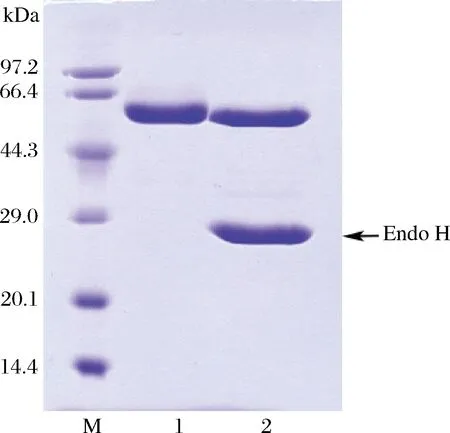
M-低分子質量標準蛋白;1-純酶液;2-經Endo H酶切除糖基后的蛋白圖2 重組α-淀粉酶McAmyA的SDS-PAGE分析Fig.2 SDS-PAGE of recombinant α-amylase McAmyA
2.3 重組α-淀粉酶McAmyA的最適pH和最適溫度
重組α-淀粉酶McAmyA的最適pH和最適溫度見圖3。α-淀粉酶McAmyA的最適pH為6.5 (圖3-A),最適反應溫度為65 ℃(圖3-B),大于65 ℃,酶活降低,在90 ℃基本失活。
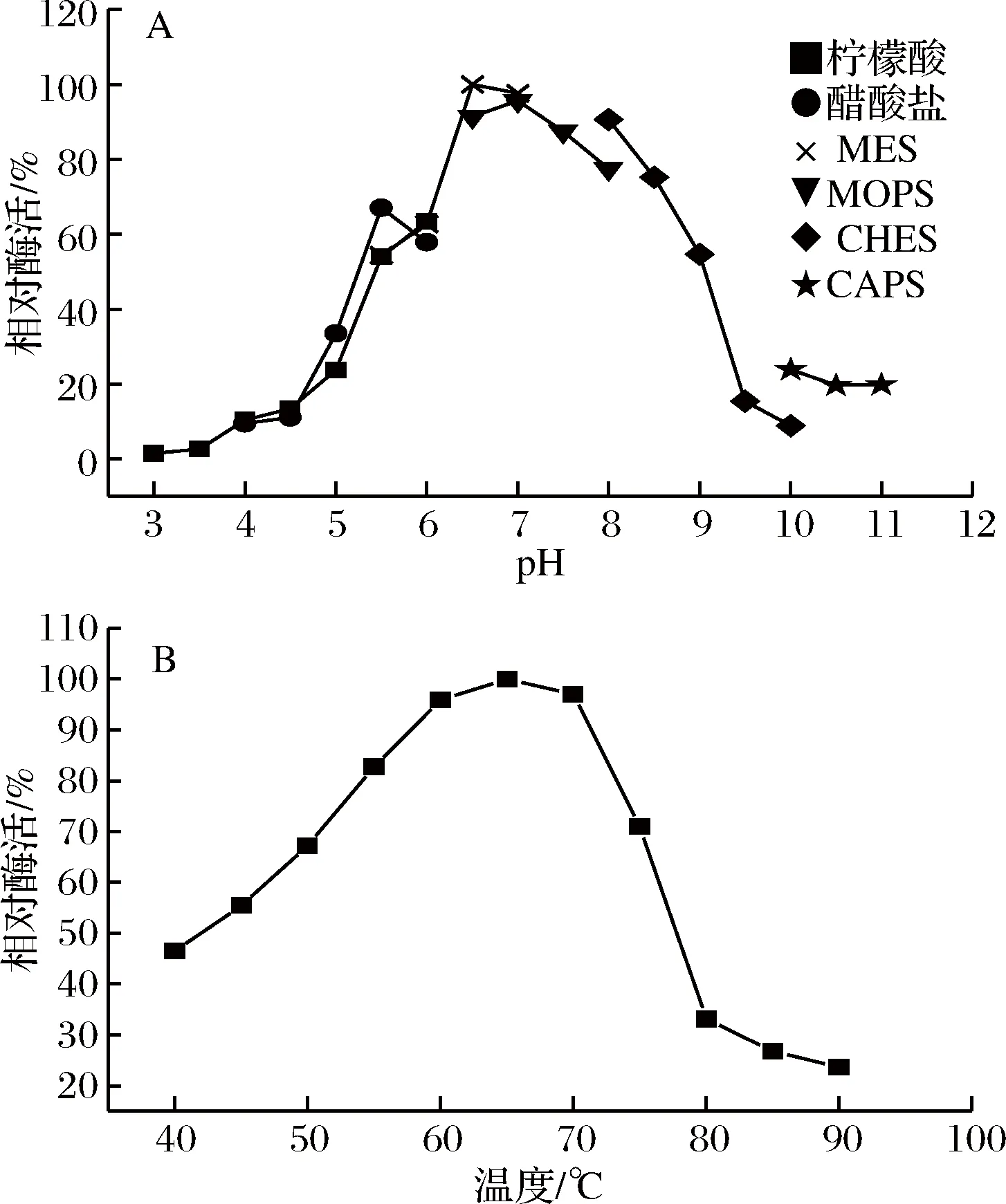
圖3 重組α-淀粉酶McAmyA的最適pH(A)和最適溫度(B)Fig.3 Optimal pH (A) and optimal temperature (B) of McAmyA
重組α-淀粉酶McAmyA與野生型樟絨枝霉α-淀粉酶的最適pH和最適溫度一致,最適pH接近于Fusicoccumsp.來源的α-淀粉酶[16](pH 7.0),高于Paecilomycessp.來源的α-淀粉酶[17](pH 4.0)。最適溫度低于Fusicoccumsp.[16]來源的α-淀粉酶(70 ℃),高于Geomycespannorum來源的α-淀粉酶[18](40 ℃)。
2.4 重組α-淀粉酶McAmyA制備麥芽糖漿
2.4.1 重組α-淀粉酶McAmyA加酶量對麥芽糖含量的影響
工業生產中,淀粉液化后直接進入糖化階段,因此選用液化液為底物,在溫度60 ℃和自然pH的條件下水解8 h,重組α-淀粉酶McAmyA加酶量對麥芽糖含量的影響如圖4所示。
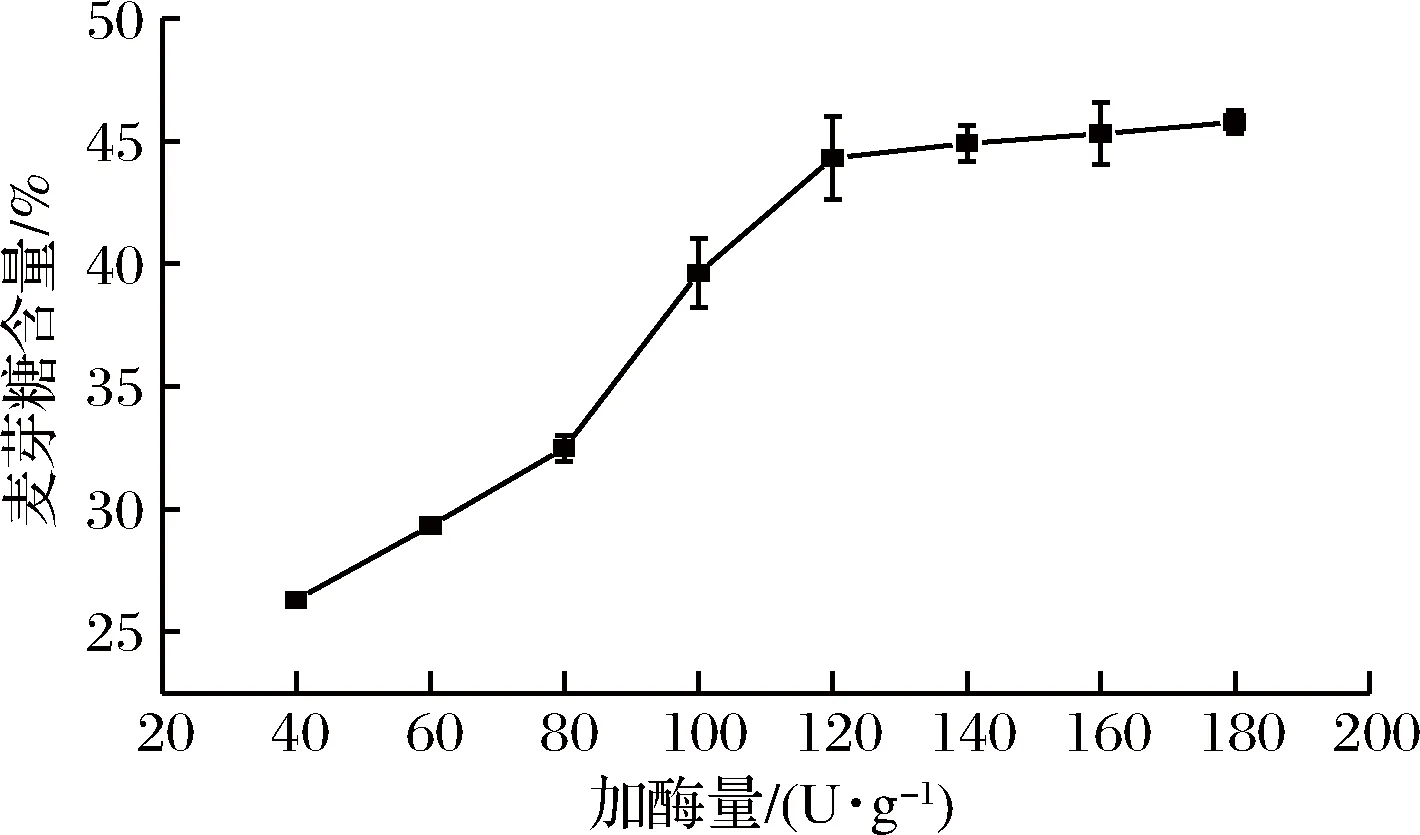
圖4 重組α-淀粉酶McAmyA加酶量對麥芽糖含量的影響Fig.4 Effect of recombinant α-amylase McAmyA dosage on maltose content
加酶量小于120 U/g時,隨加酶量增加,麥芽糖含量明顯增加,加酶量大于120 U/g后,麥芽糖含量增加緩慢,同時考慮生產成本和糖化效率,選擇120 U/g為最適加酶量,此時的麥芽糖含量為44.3%,葡萄糖含量為14.4%(質量分數),水解率為54.6%。
2.4.2 水解溫度對麥芽糖含量的影響
以液化液為底物,在加酶量120 U/g,水解8 h的條件下,水解溫度對麥芽糖含量的影響見圖5。
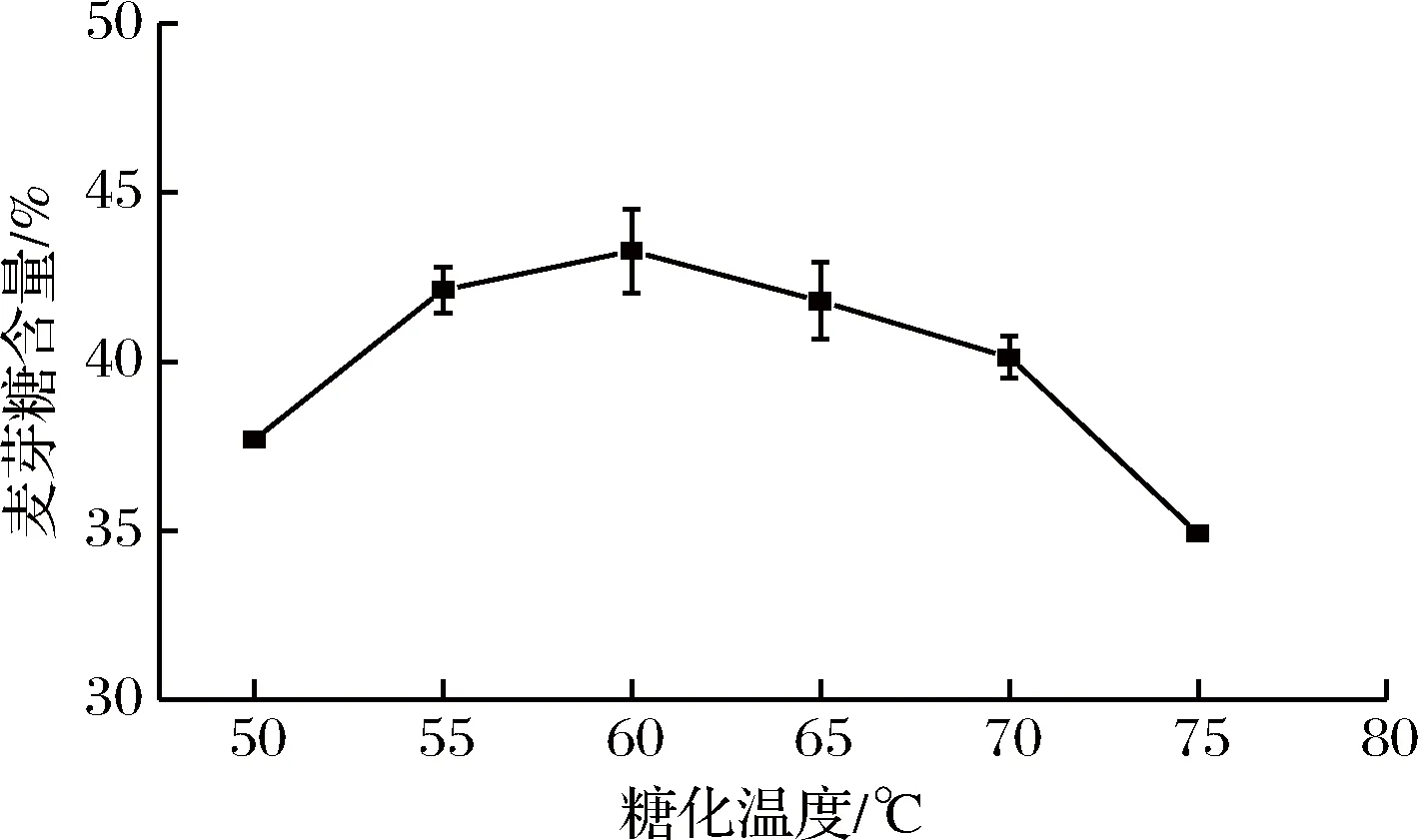
圖5 水解溫度對麥芽糖含量的影響Fig.5 Effect of hydrolysis temperature on maltose content
溫度小于60 ℃時,隨溫度升高,麥芽糖含量增加,糖化溫度60 ℃時,麥芽糖含量達到最大,最大為43.3%,葡萄糖含量為15.7%,此時水解率為54.9%,溫度大于60 ℃后,酶活穩定性差,水解率降低,麥芽糖含量減少。
2.4.3 水解時間對麥芽糖含量的影響
以液化液為底物,在加酶量120 U/g和水解溫度60 ℃的條件下,水解時間對麥芽糖含量的影響結果如圖6所示。隨水解時間增加,麥芽糖含量逐漸增加(圖6-A),水解時間大于14 h,麥芽糖含量增加緩慢,水解24 h時,主要產物是葡萄糖、麥芽糖、麥芽三糖和少量麥芽四糖(圖6-B),麥芽糖含量為50.0%(圖6-B),葡萄糖含量為40.4%,此時的水解率為83.44%,達到GB/T 20883—2007對麥芽糖漿的要求,即麥芽糖含量≥50%。
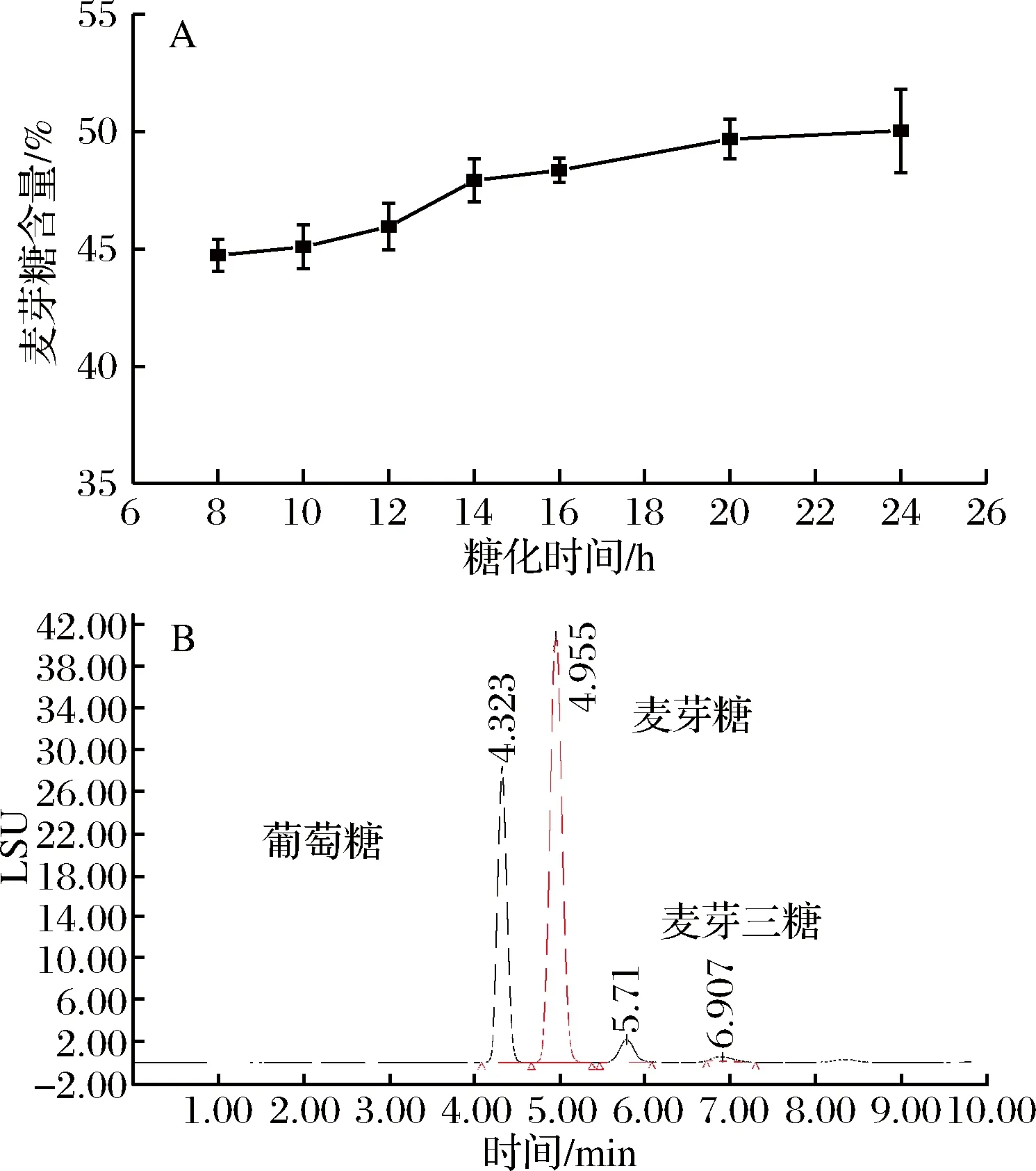
圖6 水解時間對麥芽糖含量的影響(A)和水解24 h糖化液的HPLC色譜圖(B)Fig.6 Effect of time on maltose content (A) and HPLC chromatogram of maltose syrup at the optimal condition (B)
與已報道制備麥芽糖漿的研究相比,如張正文等[19]利用真菌α-淀粉酶在60 ℃和pH 5.5的條件下,水解40 h制備得麥芽糖含量為60%,葡萄糖含量為9.0%的啤酒用麥芽糖漿。易翠平等[10]以大米淀粉為原料,利用真菌淀粉酶在58 ℃下,水解10 h,制備得麥芽糖含量為45.18%,葡萄糖含量為4.75%的麥芽糖漿。郭俊珍等[20]在55 ℃下,水解44 h制備得76.8%的麥芽糖漿。重組α-淀粉酶McAmyA制備的麥芽糖漿中葡萄糖雖然含量較高,但是糖化溫度較高,時間較短,在糖化過程中具有一定優勢。
3 結論
將嗜熱真菌樟絨枝霉α-淀粉酶基因McAmyA在畢赤酵母中高效表達,高密度發酵168 h時,蛋白表達量為13.2 mg/mL,胞外酶活力達13 440 U/mL。重組α-淀粉酶McAmyA粗酶液經強陰離子交換層析純化得到電泳級純酶,酶學性質研究表明,重組α-淀粉酶McAmyA的最適pH和最適溫度分別為6.5 和65 ℃。以淀粉液化液為底物,在60 ℃、加酶量120 U/g和自然pH的條件下,水解24 h,重組α-淀粉酶McAmyA制備得麥芽糖含量為50.0%的麥芽糖漿。表明該酶在制備麥芽糖漿中具有很大應用潛力。
